
氮肥运筹对不同夏玉米品种碳氮代谢协调性的影响
2017-07-24 15:09苌建峰董朋飞王秀玲刘卫玲李潮海
中国农业科学 2017年12期
苌建峰,董朋飞,王秀玲,刘卫玲,李潮海
(河南农业大学农学院,郑州 450002)
氮肥运筹对不同夏玉米品种碳氮代谢协调性的影响
苌建峰,董朋飞,王秀玲,刘卫玲,李潮海
(河南农业大学农学院,郑州 450002)
【目的】在不同的氮肥水平下研究不同夏玉米品种碳、氮代谢差异,分析其协调性,提出高产、稳产的生理学指标,探索根据代谢型进行差异化管理的新途径。【方法】2009—2011年在河南温县和郑州,利用大田和盆栽两种种植方式,60 000株/hm2密度,设计0、120、240、360 kg·hm-24个氮肥水平,比较郑单958(ZD958)、农大108(ND108)、浚单20(XD20)、豫单2002(YD2002)等4个夏玉米品种的产量、叶片持绿性、氮吸收和转运、不同器官C/N、叶片PEP羧化酶和RUBP羧化酶活性等指标的差异,分析4个品种在不同氮肥水平下的碳、氮代谢协调性。【结果】(1)盆栽试验中,不同氮肥水平间产量比较,XD20差异最大,ZD958差异最小;随着氮肥水平提高,产量呈增加趋势。但大田试验中,除XD20之外,其余品种在N240和N360条件下差异不显著。(2)对灌浆期氮的吸收和转运比较,低氮条件下,ZD958营养器官氮优先向叶片转移,能保持较好的持绿性;ND108对氮的吸收和运转量都较大,持绿性好,但有效利用较低;XD20在低氮条件下氮的吸收量较低,缺乏优先供应叶片的机制,易早衰;YD2002在4个氮肥水平下均表现为灌浆期营养器官氮向籽粒转移量过大,根系吸收量小,易早衰。(3)在灌浆期,正常施肥条件下,ZD958和XD20叶片碳氮比介于YD2002和ND108之间;在成熟期,ZD958和XD20的籽粒碳氮比较高。(4)在灌浆中期,ZD958的PEPCase/RUBPCase最高,且与其余3个品种差异显著;YD2002始终处于较低水平;XD20和ND108随氮肥水平增加提高显著。【结论】玉米品种良好的碳氮代谢协调性表现在碳水化合物和氮素的转运过程中,碳氮代谢协调性较好的品种能很好地协调光合产物和氮素在籽粒灌浆和维持营养器官功能需求之间的矛盾,具有较高的PEPCase/RUBPCase,从而保证高产、稳产。
玉米;碳代谢;氮代谢;C/N;PEP羧化酶;RUBP羧化酶
0 引言
【研究意义】碳、氮代谢是植物体内最基本的两大代谢过程。不同玉米品种在氮的吸收、同化、利用等方面均存在差异[1-6],对氮肥的反应也不相同[3,4,7-9]。根据不同玉米品种的碳、氮代谢所表现出来的差异,采取不同的氮肥水平进行田间管理,是优化施肥,提高养分利用效率,提高产量的一种重要途径。【前人研究进展】氮效率高的玉米品种在氮的吸收和利用上达到完美的组合,能够较多地利用营养器官中的氮[3]。ECHARTE L等[4]研究发现,新品种对于低氮的忍耐能力高于老品种。向春阳等[10]研究发现,不同基因型玉米的吸氮量差异在低氮条件下更显著。PEP羧化酶和RUBP羧化酶是光合作用的关键酶,有研究表明,将PEP羧化酶导入到水稻、小麦等C3作物以后,显著提高了光合速率[11-13]。对不同玉米品种在不同氮肥水平下光合作用关键酶活性比较发现,低氮条件下PEP羧化酶活性差异显著,RUBP羧化酶活性差异较小[14]。碳代谢和氮代谢相辅相成,互相影响,有研究表明,PEPC的高表达基因会对氮代谢相关酶活性产生影响[15],而提高氮肥用量可以显著提高RUBP羧化酶的活性[16]。【本研究切入点】前人关于玉米碳、氮代谢的研究较多,但多集中在不同基因型品种氮效率的差异上。对碳、氮代谢的协调性进行系统研究,并采用不同的氮肥水平进行调控,使碳、氮代谢更加协调,从而实现节肥高效的研究较少。【拟解决的关键问题】本试验通过对国内推广面积较大的部分玉米杂交种进行长期观察,对叶色、持绿性、稳产性、丰产性等指标进行比较,筛选了4个具有不同代谢类型特征的典型玉米杂交种作为研究材料,比较其在不同氮肥水平下碳、氮代谢表现出的差异,提出玉米高产、稳产的生理学指标,探索根据代谢类型进行差异化管理的新途径,为玉米新品种选育和栽培管理提供理论和技术参考。
1 材料与方法
1.1 试验概况
于2009—2010年在河南省温县进行大田试验,2010—2011年在位于河南郑州市的河南农业大学科教园区进行盆栽试验。两个区域均位于华北平原南部,属北温带大陆性季风气候。郑州市2010和2011年平均气温分别为15.6和15.1℃,年降雨量分别为600.3和706.5 mm。试验地为偏砂质潮土,取0—20 cm土层土,混匀后过筛装盆。土壤有机质含量10.2 g·kg-1,全氮含量为0.93 g·kg-1,速效磷含量为11.2 mg·kg-1,速效钾含量157.6 mg·kg-1。
温县2009和2010年平均气温分别为15.0和15.1℃,年降雨量分别为497.8和603.7 mm,试验地为壤质潮土,0—20 cm土层有机质含量10.8 g·kg-1,全氮含量为1.1 g·kg-1,速效磷含量为16.7 mg·kg-1,速效钾含量137 mg·kg-1。
1.2 试验材料与设计
选用4个玉米杂交种,分别为郑单958(ZD958)、浚单20(XD20)、农大108(ND108)、豫单2002(YD2002)。4个氮肥处理,分别0、120、240、360 kg·hm-2,氮肥(尿素)于5叶期和9叶期按1﹕1比例分两次施入,磷(过磷酸钙)、钾肥(氯化钾)于5叶期施入,用量分别为150 kg·hm-2和120 kg·hm-2。大田和盆栽均按照60 000株/hm2种植,采用裂区设计,以氮肥为主区,品种为副区,3次重复,采用随机区组摆列。盆栽试验的盆深为30 cm,直径35 cm,每盆装土16 kg,每个小区种植20盆,采用1 000 mL烧杯定量浇灌,保证每盆浇水量一致。试验均在每年的6月10日播种,9月底收获。大田试验管理同一般高产田。
1.3 测定项目和方法
1.3.1 植株全氮测定 采用凯氏定氮法。
1.3.2 叶片SPAD测定 用SPAD-502叶绿素计于不同时期测定植株叶片,每个叶片测10次,取平均值。吐丝前,测上部展开叶,吐丝期及以后测穗位叶。
1.3.3 全碳测定 采用重铬酸钾容量法。
1.3.4 PEP羧化酶和RUBP羧化酶活性测定 参考魏爱丽等的方法,略有修改[17]。取各器官鲜样0.5 g,加3 mL预冷的提取缓冲液,含0.1 mol·L-1Tris-HCl(pH7.8),1 mmol·L-1EDTA,7 mmol·L-1巯基乙醇,10%的甘油和1% PVP,研磨后在15 000×g下离心15 min,上清液即为酶提取液。
测定PEP羧化酶活性:1 mL反应混合液含50 mmol·L-1Tris-HCl(pH 7.8),10 mmol·L-1MgCl2,0.25 mmol·L-1EDTA,5.0 mmol·L-1NaHCO3,5.0 mmol·L-1DTT,0.1 mmol·L-1NADH,4 U·mL-1MDH,2.0 mmol·L-1PEP,加酶液启动反应,在340 nm波长下观察3 min光密度的下降值。
测定RuBP羧化酶活性:1 mL反应液含50 mmol·L-1Tricine-NaOH(pH 7.9),10 mmol·L-1EDTA,2 mmol·L-1DTT,0.2 mmol·L-1NADH,5 mmol·L-1DTT ATP,15 mmol·L-1MgCl2,10 mmol·L-1NaHCO3,5 mmol·L-1磷酸肌酸,2 U·mL-1肌酸,4 U·mL-1三磷酸甘油醛脱氢酶,4 U·mL-1三磷酸甘油醛激酶和酶提取液,在25℃下保温5 min,加入0.5 mmol·L-1RUBP启动反应。
1.3.5 籽粒氮来源计算 营养器官氮转移量=吐丝期营养器官氮积累量-成熟期营养器官氮积累量
根系吸收氮量=成熟期籽粒氮积累量-营养器官氮转移量。
由于本试验未测定根系指标,故本文试验结果中涉及营养器官的有关指标,均不包含根系。
1.3.6 测产 成熟期收获,大田试验每个小区选取中间两行,称取鲜重,取标准穗10穗(将所取的全部鲜穗称重,除以株数,得出单株玉米的平均穗重,标准穗选取按照总重量等于单株平均穗重乘以标准穗数量的方法进行),脱粒后计算出籽率和含水率,计算14%含水率的产量。盆栽试验每个处理留10株玉米计产,方法同上。
1.4 统计分析
为保证试验数据的准确度,本试验结果以管理相对精准的盆栽试验数据为主,同时用两年大田试验的产量结果与之对比。利用DPS 14.5对试验数据进行差异显著性检验(LSD),利用SigmaPlot 12.5软件绘制图表。
2 结果
2.1 氮肥水平对不同品种产量的影响
从图1可以看出,在盆栽试验中,ZD958两年均以N240处理产量最高,且与其他氮肥处理差异显著;其余3个品种两年均以N360处理最高,但ND108在2011年与N240处理差异不显著。盆栽试验中XD20产量在氮肥处理间差异最大,N360处理两年分别比N0处理高出28.1%和12.9%;氮肥处理间差异最小的为ZD958,N240处理两年分别比N0处理高出2.0%和4.3%。
在大田试验中, XD20两年的产量均表现为N360最高,且与其余氮肥处理差异显著;其余3个品种均表现为N240和N360处理较高,且差异不显著。氮肥水平间的差异以XD20最大,N360处理两年分别比N0处理高出22.4 %和32.7 %。
对盆栽和大田试验的4个品种在不同氮肥水平下产量分别综合比较,均表现为ZD958>XD20>ND108>YD2002。
2.2 氮肥水平对不同品种叶片SPAD的影响
从图2可以看出,吐丝以后,ZD958叶片SPAD两年平均值均表现为N360>N240>N120>N0,且N360处理两年均显著高于N0处理。成熟期,N360处理两年平均分别比N240、N120、N0处理高出1.0%、3.9%、11.8%。
ND108两年平均在吐丝以后均表现为N240>N360>N120>N0;成熟期,N240处理两年平均分别比N360、N120、N0处理高出0.6%、2.5%、8.6%,N360和N0处理两年均差异显著。
XD20处理在全生育期均表现为N360>N240>N120>N0;随着生育期的推移,N0处理与其余3个氮肥处理的差异逐渐加大,在成熟期,N360处理两年平均分别比N240、N120、N0处理高出5.8%、18.2%、66.5%,且4个处理在两年间均差异显著。
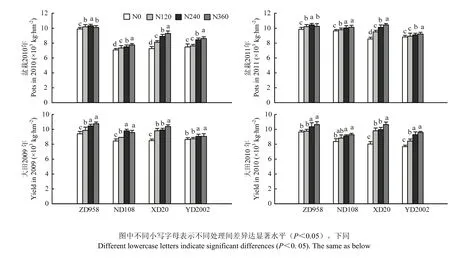
图1 不同玉米品种在不同氮肥水平下产量比较Fig. 1 Comparison of yield of different maize varieties under different nitrogen levels
YD2002吐丝以后,两年均表现为N360>N240>N120>N0;吐丝后35 d开始,叶片SPAD值迅速下降;成熟期,N360处理两年平均分别比N240、N120、N0处理高出6.2%、145.6%、215.9%,2010年4个处理间均差异显著,2011年表现为N360和N240处理显著高于N120和N0处理。
2.3 氮肥水平对不同玉米品种氮积累量及籽粒氮来
源的影响
从表1可以看出,吐丝期营养器官氮积累量,4个氮肥水平下均表现为ND108和YD2002高于ZD958和XD20;成熟期营养器官氮积累量,4个氮肥水平下均以ND108最高,且与其余3个品种均差异显著。
对成熟期籽粒氮来源比较发现,营养器官氮转移量随着氮肥水平升高表现出增加趋势,但在品种和氮肥处理间存在差异。ZD958、ND108、XD20、YD2002籽粒氮来源中,营养器官转移量在N360处理下比N0处理下分别高出28%、24%、26%、3%。对籽粒氮来源中的根吸收量比较发现,ND108在4个氮肥水平下均最高;YD2002均最低,且与其余3个品种差异显著;营养器官转移量,YD2002在4个氮肥水平下均显著高于其余3个品种。
2.4 氮肥水平对不同品种各器官C/N的影响
从图3可以看出,4个品种茎秆C/N随着生育期延长,呈现出逐渐升高的趋势;且在9叶期以后,4个品种均表现出高氮处理低于低氮处理的现象,且差异显著。品种间比较,吐丝以后, N0和N120(低氮)处理下,ZD958和YD2002显著高于ND108和XD20;在N360(高氮)处理下,4个品种吐丝以后C/N差异不显著。
从图4可以看出,4个品种叶片C/N随着生育期延长呈现出逐渐升高的趋势;叶片C/N随氮肥水平的变化在4个品种间表现不一致。品种间比较,在吐丝后的21 d和44 d,在4个氮肥处理中,都表现为YD2002最高,ND108最低,ZD958和XD20介于二者之间。
从图5可以看出,4个品种籽粒C/N随着灌浆期的延长均表现出“先升高后降低”的趋势;在灌浆后期,4个品种均表现出N0处理高于其余处理的现象。品种间比较,灌浆后期4个氮肥水平下均以ZD958最高,XD20次之,YD2002最低。
2.5 氮肥水平对不同品种碳代谢相关酶活性的影响
从图6可以看出,叶片PEP羧化酶活性在整个生育期呈现“先上升后下降”的趋势。品种间比较,6叶期,YD2002在4个氮肥处理中都处于较高水平。吐丝期,N360处理下比较,ZD958最高,XD20最低;其余3个氮肥水平下均以ND108最高。吐丝后21 d,ZD958在4个氮肥处理中均最高;随着氮肥施用量增加,ND108迅速上升。吐丝后44 d,YD2002始终处于较低水平。吐丝后,XD20随着氮肥水平提高,PEP羧化酶活性提高显著,但和其余3个品种比较,仍处于较低水平。

图2 不同氮肥水平下4个玉米杂交种叶片SPAD值比较Fig. 2 Comparison of leaf SPAD values of 4 maize hybrids under different nitrogen levels
从图7可以看出,叶片RUBP羧化酶活性在整个生育期较为平稳。品种间比较,吐丝以后,ND108在各氮肥水平均处于较高水平,XD20始终处于较低水平。吐丝后21 d和44 d,YD2002在低氮水平下与其余3个品种比较处于较高水平,但在高氮水平下处于较低水平。6叶期,ZD958在4个氮肥处理中均处于较低水平。
从图8可以看出,在吐丝后21 d,ZD958的PEPCase/RUBPCase在4个氮肥处理中始终最高,且与其余3个品种差异显著;而YD2002始终最低。吐丝后21 d,随着氮肥施用量的增加,XD20和ND108的PEPCase/RUBPCase提高显著,而ZD958和YD2002随施氮量变化不显著。

表1 不同氮肥水平下4个玉米杂交种氮积累量及籽粒氮来源比较(2010年和2011年平均,郑州)Table 1 Comparison of nitrogen accumulation and grain nitrogen sources in 4 maize hybrids under different nitrogen levels (Average of 2010 and 2011, Zhengzhou)
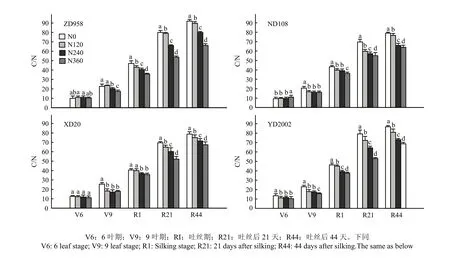
图3 4个玉米杂交种在不同氮肥水平下不同生育时期茎秆C/N比较(2010和2011年平均,郑州)Fig. 3 C/N in stalks of 4 hybrids of maize under different nitrogen levels at different growth stages (Average of 2010 and 2011, Zhengzhou)
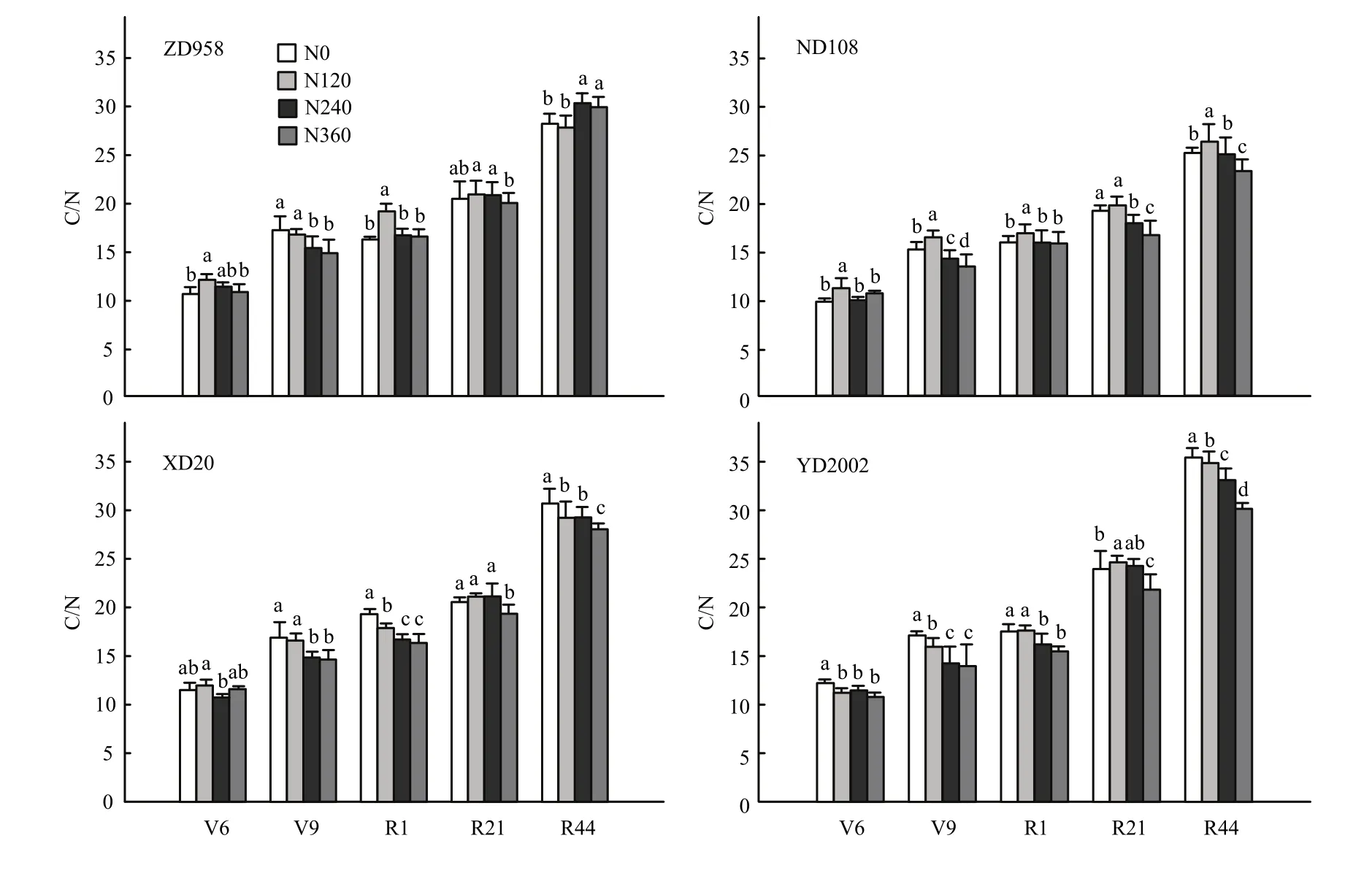
图4 4个玉米杂交种在不同氮肥水平下不同生育时期叶片C/N比较(2010和2011年平均,郑州)Fig. 4 C/N in leaves of 4 hybrids of maize under different nitrogen levels at different growth stages (Average of 2010 and 2011, Zhengzhou)

图5 4个玉米杂交种在不同氮肥水平下吐丝后籽粒C/N比较(2010和2011年平均,郑州)Fig. 5 C/N in grains of 4 hybrids of maize under different nitrogen levels after silking stages (Average of 2010 and 2011, Zhengzhou)
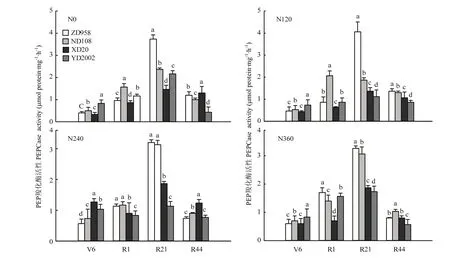
图6 4个玉米杂交种在不同氮肥水平下不同时期叶片PEP羧化酶活性比较(2010和2011年平均,郑州)Fig. 6 PEPCase in leaves of 4 hybrids of maize under different nitrogen levels at different growth stages (Average of 2010 and 2011, Zhengzhou)

图7 4个玉米杂交种在不同氮肥水平下不同时期叶片RUBP羧化酶活性比较(2010和2011年平均,郑州)Fig. 7 RUBPCase in leaves of 4 hybrids of maize under different nitrogen levels at different growth stages (Average of 2010 and 2011, Zhengzhou)

图8 4个玉米杂交种在不同氮肥水平下不同时期叶片PEPCase/RUBPCase比较(2010和2011年平均,郑州)Fig. 8 Ratios of PEPCase and RUBPCase in leaves of 4 hybrids of maize under different nitrogen levels at different growth stages (Average of 2010 and 2011, Zhengzhou)
3 讨论
3.1 品种间产量比较
前人研究证明,作物产量不会随着氮肥水平的提高一直增加,到达阈值后会停止增加或降低[18,19],但是,不同玉米品种对氮肥的需求量和氮效率不尽相同。ECHARTE等[4]对不同年代玉米杂交种氮利用效率研究发现,玉米对氮的吸收能力存在基因型差异,在低氮条件下差异更为显著,表现为新品种对于低氮的耐性优于老品种。根据氮效率将玉米划分为若干个类型的相关研究也较多[20-24]。在盆栽试验中,4个品种产量均表现出随着氮肥水平提高而增加的趋势,但在两年的大田试验中,除XD20外,其余3个品种在N360和N240水平下,产量差异均不显著。由此可见,对于大部分品种,在氮肥使用量达到240 kg·hm-2之后,增加氮肥施用量对产量的增益并不明显。品种间比较,ZD958随着施氮水平的变化,产量变化相对较小;XD20变化较大。由此可见,ZD958对低氮的耐性更强,XD20较弱。
3.2 氮吸收与转运比较
玉米对土壤氮的利用由氮的吸收效率与利用效率共同决定,MONASTORIO O等[25]认为,玉米在低氮下氮效率的变异主要来源于吸收效率,而在高氮水平下利用效率起主要作用。米国华等[26]有不同的研究结果,低氮高氮下,玉米的吸收效率和利用效率对氮效率均作用显著。有研究认为,至少3种氮代谢过程影响玉米产量,分别为抽丝期后的吸氮能力,籽粒灌浆速率与持续时间,体内醇溶蛋白的合成速度[27]。在本试验中,4个品种叶片SPAD叶色值变化各有特点。其中,ZD958和ND108属于持绿性较好的品种,叶片功能期较长,且在不同的氮肥水平下变化不显著;XD20在低氮条件下易早衰,但提高氮肥用量以后,持绿性明显改善;YD2002也属于早衰性品种,但提高氮肥水平对其叶片的持绿性并无明显改善。
叶片的持绿性与氮素的吸收和转运密切相关。与其余品种相比,ZD958在吐丝期和成熟期茎中的氮积累量较低,特别是在低氮水平下表现更加明显;而叶片中的氮积累量则处于正常水平,且随氮肥水平变化较小。由此可见,ZD958在低氮条件下吸收的氮会优先分配给叶片,从而保证正常的光合作用,这种对氮素的良好调节能力保证其有良好的持绿性外,还使其更加耐受低氮。对于该类品种,生产中可以适当降低氮肥用量。
从试验结果看,XD20和ZD958地上部营养器官对氮的吸收量差异不显著,但是在低氮水平下,XD20茎秆的氮积累量显著高于ZD958,高氮水平下二者差异不显著。由此可见,与ZD958相比,XD20在低氮水平下缺乏优先保证叶片氮素供应的机制,表现为对氮肥敏感,低氮条件下易早衰;提高氮肥供应水平以后,对氮素的吸收和利用则趋于平衡。
ND108在吐丝以前,地上部营养器官中积累了大量的氮,吐丝以后对氮的吸收量较大。由此可见,ND108叶片良好的持绿性主要依靠的是对氮的大量吸收。另外,ND108成熟期营养器官残留的氮显著高于其余3个品种,降低了氮素的利用效率。在生产中,该类品种应保证充足的氮素供应。
YD2002吐丝期地上部营养器官具有较高的氮积累量,吐丝以后,营养器官中的氮向籽粒的转移量显著高于其他品种,根系的氮吸收量却显著低于其他品种。由此可见,该品种早衰是由于吐丝后营养器官氮大量向籽粒转移,根系吸收氮能力较弱造成的。从籽粒氮来源看,在低氮水平下,ZD958、ND108、XD20都会降低氮由营养器官向籽粒的运转量,从而保证叶片功能;YD2002在4个氮肥水平下,营养器官向籽粒的运转量无显著差异,缺乏上述优良机制,因此,提高施氮量对于改善该品种持绿性并无明显效果。
3.3 器官中C/N比较
碳代谢和氮代谢存在着密切的关系,氮代谢的进行需要依赖于碳代谢提供碳源和能量,而碳代谢又需要氮代谢提供酶蛋白和光合色素[28]。碳、氮代谢之间即相互依赖,又存在一定的矛盾。前人研究表明,光合作用产生的能量及其中间产物大部分用于碳、氮代谢,在某些组织中氮代谢甚至可消耗掉光合作用能量的55%[29],可见,过高的氮代谢不利于光合产物积累,从而影响产量形成。DUVICK[30]对不同年代的玉米杂交种籽粒蛋白质和淀粉含量的比较结果也印证了上述观点:籽粒蛋白质含量平均每十年降低0.3%,淀粉含量平均每十年增加0.3%,究其原因,也是因为生产淀粉比生产蛋白质消耗的能量少,所以在品种选育过程中间接选择了淀粉多蛋白少的基因型。可见,在灌浆期间,适当降低植株体内的氮代谢,增加碳代谢,从而保持籽粒中较高的碳氮比,有利于提高玉米的增产潜力。
与籽粒较高的碳氮比不同,灌浆期间叶片的碳氮比应该保持在一个合理的区间。原因在于叶片碳氮比过低,会造成碳水化合物含量较低,或者叶片中氮含量过高,前者不利于光合物质的贮藏和转运,后者消耗过多的能量,都会影响产量形成。而碳氮比过高,会造成叶片含氮量过低,或者碳水化合物含量过高,前者不利于维持叶片功能,后者也会对光合作用产生反馈抑制[31],原因有3个:(1)淀粉颗粒分解,淀粉含量增加,导致叶肉细胞中CO2扩散阻力增加[32];(2)蔗糖合成和己糖循环导致磷酸中间产物增加,降低无机磷酸盐水平,低含量的无机磷酸盐降低叶绿体中RUBPCO的活性,降低ATP/ADP比率,导致RUBPCO失活[33];(3)糖含量控制着光合速率基因的表达[34-35]。根据本试验研究的结果,在正常的氮肥供应水平下,ZD958和XD20灌浆期叶片C/N介于YD2002和ND108之间,籽粒C/N高于ND108和YD2002,属于优良性状。
3.4 碳代谢相关酶活性比较
前人对不同年代的玉米杂交种在不同氮肥水平下光合关键酶活性进行比较发现,老品种在低氮处理中PEP羧化酶的减少比较显著,但是所有品种中,RUBP羧化酶活性都没有显著差异[14,36],分析原因,可能是氮素缺乏时,可利用的氮优先保证了RUBP羧化酶的合成,导致了PEP羧化酶合成的减少[36]。在本试验中,也发现了同样的规律。由此可见,在碳代谢的过程中,作为在直接合成碳水化合物的C3过程中起重要作用的RUBP羧化酶,其活性相对稳定;而PEP羧化酶的活性受到外界环境、品种等因素的影响较大。因此,在品种选育过程中,叶片PEP羧化酶能够保持较高且稳定的活性,不仅有利于提高增产潜力,还有利于提高品种的抗逆性,显然是一种十分优良的性状。在本试验中,用PEPCase/RUBPCase来表征上述性状,对本试验4个品种比较发现,灌浆中期ZD958叶片的PEPCase/RUBPCase在4个氮肥水平下均显著高于其余3个品种;YD2002最低,XD20和ND108随着氮肥水平提高,增加显著。这个结果也对ZD958良好的丰产性和稳产性进行了印证。
4 结论
本试验通过对不同碳氮代谢特征的玉米品种进行比较,发现碳氮代谢协调性较好的品种(如ZD958)在碳水化合物和氮素的转运过程中,能很好地协调光合产物和氮素在籽粒灌浆和维持营养器官功能需求之间的矛盾,特别是在土壤氮素水平较低的条件下,这种优势更加明显;另外,较高的PEPCase/RUBPCase也是该类品种的特征。上述特征保证了该类品种适应性更广,高产潜力更大,可以作为玉米品种选育和栽培管理的依据。
[1] GERLOFF G C. Intact-plant screening for tolerance of nutrient-deficiency stress. Plant and Soil, 1987, 99(1): 3-16.
[2] MOLL R H, KAMPRATH E J, JACKSON W A. Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization. Agronomy Journal, 1982, 74(3): 562-564.
[3] WORKU M, BÄNZIGER M, ERLEY G S A , FRIESEN D, DIALLO A O, HORST W J. Nitrogen uptake and utilization in contrasting nitrogen efficient tropical maize hybrids. Crop Science, 2007, 47(2): 519-528.
[4] ECHARTE L, ROTHSTEIN S, TOLLENAAR M. The response of leaf photosynthesis and dry matter accumulation to nitrogen supply in an older and a newer maize hybrid. Crop Science, 2008, 48(2): 656-665.
[5] 钱春荣, 于洋, 宫秀杰, 姜宇博, 赵杨, 郝玉波, 李梁, 张卫建. 黑龙江省不同年代玉米杂交种产量对种植密度和施氮水平的响应.作物学报, 2012, 38(10): 2069-2077.
QIAN C R, YU Y, GONG X J, JIANG Y B, ZHAO Y, HAO Y B, LI L, ZHANG W J. Response of grain yield to plant density and nitrogen application rate for maize hybrids from different eras in Heilongjiang province. Acta Agronomica Sinica, 2012, 38(10): 2069-2077. (in Chinese)
[6] 张仁和, 杜伟莉, 郭东伟, 张爱瑛, 胡富亮, 李凤艳, 薛吉全. 陕西省不同年代玉米品种产量和氮效率性状的变化. 作物学报, 2014, 40(5): 915-923.
ZHANG R H, DU W L, GUO D W, ZHANG A Y, HU F L, LI F Y, XUE J Q. Changes of grain yield and nitrogen use efficiency of maize hybrids released in different eras in Shaanxi province. Acta Agronomica Sinica, 2014, 40(5): 915-923. (in Chinese)
[7] TSAI C Y, HUBER D M, GLOVER D V, WARREN H L. Relationship of N deposition to grain yield and N response of three maize hybrids. Crop Science, 1984, 24(2): 277-281.
[8] 陈新平, 周金池, 王兴仁, 张福锁. 冬小麦、夏玉米不同品种(系)之间的氮营养效率的差异. 中国农业大学学报, 2000, 5(1): 80-83.
CHEN X P, ZHOU J C, WANG X R, ZHANG F S. Difference in nitrogen efficiency among different winter wheat and summer corn varieties. Journal of China Agricultural University, 2000, 5(1): 80-83. (in Chinese)
[9] 张瑞珍, 张恩和, 孙长占. 不同基因型玉米品种氮素营养效率差异的研究. 吉林农业大学学报, 2003, 25(2): 183-186.
ZHANG R Z, ZHANG E H, SUN C Z. Study different genotype maize variety on diversity of nitrogen nutrition efficience. Journal of Jilin Agricultural University, 2003, 25(2): 183-186. (in Chinese)
[10] 向春阳, 凌碧莹, 关义新, 马兴林, 张宝石. 氮肥及基因型差异对玉米吸氮量的反应. 中国农学通报, 2003, 19(3): 26-28.
XIANG C Y, LING B Y, GUAN Y X, MA X L, ZHANG B S. Effect of nitrogen fertilizer and the genotypes on nitrogen uptake of maize. Chinese Agricultural Science Bulletin, 2003, 19(3): 26-28. (in Chinese)
[11] REN C G, LI X, LIU X L, DAI C C. Hydrogen peroxide regulated photosynthesis in C4-pepc transgenic rice. Plant Physiology & Biochemistry, 2014, 74: 218-229.
[12] 季本华, 朱素琴, 焦德茂. 转玉米C4光合酶基因水稻株系中的光合C4微循环. 作物学报, 2004, 30(6): 536-543.
JI B H, NIU S Q, JIAO D M. Photosynthetic C4-microcycle in transgenic rice plant lines expressing the maize C4-photosynthetic enzymes. Acta Agronomica Sinica, 2004, 30(6): 536-543. (in Chinese)
[13] 张庆琛, 许为钢, 胡琳, 李艳, 张磊, 齐学礼. 玉米C4型全长pepc基因导入普通小麦的研究. 麦类作物学报, 2010, 30(2): 194-197.
ZHANG Q C, XU W G, HU L, LI Y, ZHANG L, QI X L. Development of transgenic wheat plants with maize C4-specific pepc gene by particle bombardment. Journal of Triticeae Crops, 2004, 30(2): 194-197. (in Chinese)
[14] DING L, WANG K J, JIANG G M, BISWAS D K, XU H, LI L F, LI Y F. Effects of nitrogen deficiency on photosynthetic traits of maize hybrids released in different years. Annals of Botany, 2005, 96(5): 925- 930.
[15] LI X, Wang C. Physiological and metabolic enzymes activity changes in transgenic rice plants with increased phosphoenolpyruvate carboxylase activity during the flowering stage. Acta Physiologiae Plantarum, 2013, 35(5): 1503-1512.
[16] 王仁雷, 李霞, 陈国祥, 华春, 魏锦城. 氮肥水平对杂交稻汕优63剑叶光合速率和RuBP羧化酶活性的影响. 作物学报, 2001, 27(6): 930-934.
WANG R L, LI X, CHEN G X, HUA C, WEI J C. Effect of N-fertilizer levels on photosynthetic rate and RuBP carboxylase activity in flag leaves of hybrid rice Shanyou 63. Acta Agronomica Sinica, 2001, 27(6): 930-934. (in Chinese)
[17] 魏爱丽, 张英华, 黄琴, 王志敏. 小麦不同绿色器官光合速率与碳同化酶活性及其基因型差异研究. 作物学报, 2007, 33(9): 1426-1431.
WEI A L, ZHANG Y H, HUANG Q, WANG Z M. Dynamic characteristics of photosynthetic rate and carbon assimilation enzyme activities of different green organs in different genotypes of wheat. Acta Agronomica Sinica, 2007, 33(9): 1426-1431. (in Chinese)
[18] ZHAO C S, HU C X, HUANG W, SUN X C, TAN Q L, DI H J. A lysimeter study of nitrate leaching and optimum nitrogen applicationrates for intensively irrigated vegetable production systems in central China. Journal of Soils and Sediments, 2010, 10(1): 9-17.
[19] 杨宪龙, 路永莉, 同延安, 马海洋, 陈毓君, 丁燕. 陕西关中小麦-玉米轮作区协调作物产量和环境效应的农田适宜氮肥用量. 生态学报, 2014, 34(21): 6115-6123.
YANG X L, LU Y L, TONG Y A, MA H Y, CHEN Y J, DING Y. Optimum-N application rate to maximize yield and protect the environment in a wheat-maize rotation system on the Guanzhong Plain, Shaanxi province. Acta Ecologica Sinica, 2014, 34(21): 6115-6123. (in Chinese)
[20] 曹敏建, 衣莹. 耐低氮胁迫玉米的筛选与评价. 玉米科学, 2000, 8(4): 64-69 .
CAO M J, YI Y. Screening and evaluation of low nitrogen stress tolerance in maize. Journal of Maize Sciences, 2000, 8(4): 64-69. (in Chinese)
[21] 徐祥玉, 张敏敏, 翟丙年, 李生秀, 张兴昌, 王朝辉. 夏玉米氮效率基因型差异研究. 植物营养与肥料学报, 2006, 12(4): 495-499.
XU X Y, ZHANG M M, ZHAI B N, LI S X, ZHANG X C, WANG Z H. Genotypic variation in nitrogen use efficiency in summer maize. Plant Nutrition and Fertilizer Science, 2006, 12(4): 495-499. (in Chinese)
[22] 陈范骏, 米国华, 张福锁. 氮高效玉米新品种中农99的选育. 作物杂志, 2009(6): 103-104 .
CHEN F J, MI G H, ZHANG F S. Breeding of new maize varieties with high nitrogen efficiency of Zhongnong 99. Crops, 2009(6): 103-104. (in Chinese)
[23] 崔文芳, 高聚林, 王志刚, 崔超, 胡树平, 于晓芳, 孙继颖, 苏治军.玉米自交系氮效率基因型差异分析. 玉米科学, 2013, 21(3): 6-12.
CUI W F, GAO J L, WANG Z G, CUI C, HU S P, YU X F, SUN J Y, SU Z J. Analysis on genotypic difference in nitrogen efficiency of maize inbred lines. Journal of Maize Sciences, 2013, 21(3): 6-12. (in Chinese)
[24] 王晓慧, 曹玉军, 魏雯雯, 张磊, 王永军, 边少锋, 王立春. 我国北方37个高产春玉米品种干物质生产及氮素利用特性. 植物营养与肥料学报, 2012, 18(1): 60-68 .
WANG X H, CAO Y J, WEI W W, ZHANG L, WANG Y J, BIAN S F, WANG L C. Characteristics of dry matter production and nitrogen use efficiency of 37 spring maize hybrids with high-yielding potential in North of China. Plant Nutrition and Fertilizer Science, 2012, 18(1): 60-68. (in Chinese)
[25] ORTIZ-MONASTERIO J I, SAYRE K D, RAJARAM S, McMahon M. Genetic progress in wheat yield and nitrogen use efficiency under four nitrogen rates. Crop Science, 1997, 37(3): 898-904.
[26] 米国华, 刘建安, 张福锁. 玉米杂交种的氮农学效率及其构成因素剖析. 中国农业大学学报, 1998 (S4): 97-104.
MI G H, LIU J A, ZHANG F S. Analysis on agronomic nitrogen efficiency and its components of maize hybrids. Journal of China Agricultural University, 1998(S4): 97-104. (in Chinese)
[27] TSAI C Y, HUBER D M, GLOVER D V, WARREN H L. Relationship of N deposition to grain yield and response of three maize hybrids. Crop Science, 1984, 24(2): 277-281.
[28] 史宏志, 韩锦峰. 烤烟中碳氮代谢几个问题的探讨. 烟草科技, 1998(2): 34-36.
SHI H Z, HAN J F. Discussion on several problems of carbon and nitrogen metabolism in flue-cured tobacco. Tobacco Science and Technology, 1998(2): 34-36. (in Chinese)
[29] AND H, TURPIN D H. Integration of carbon and nitrogen metabolism in plant and algal cells. Journal of the Chemical Society, 2003, 45(1): 577-607.
[30] DUVICK D N. The contribution of breeding to yield advances in maize (Zea mays L.). Advances in Agronomy, 2005, 86(5): 83-145.
[31] RENNIE E A, TURGEON R, ZAMBRYSKI P C. A comprehensive picture of phloem loading strategies. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(33): 14162-14167.
[32] GRUB A, MACHLER F. Photosynthesis and light activation of ribulose 1,5-bisphosphate carboxylase in the presence of starch. Journal of Experimental Botany, 1990, 41(12): 1293-1301.
[33] SHARKEY T D. Feedback limitation of photosynthesis and the physiological role of ribulose bisphosphate carboxylase carbamylation. Shokubutsugaku Zasshi, 1991(2): 87-105.
[34] SCHÄFER C, SIMPER H, HOFMANN B. Glucose feeding results in coordinated changes of chlorophyll content, ribulose-1,5-bisphosphate carboxylase-oxygenase activity and photosynthetic potential in photoautotrophic suspension cultured cellsof Chenopodium rubrum. Plant, Cell & Environment, 1992, 15(3): 343-350.
[35] KRAPP A, HOFMANN B, SCHÄFER C, STITT M. Regulation of the expression of rbc S and other photosynthetic genes by carbohydrates: a mechanism for the ‘sink regulation’ of photosynthesis? Plant Journal, 1993, 3(6): 817-828.
[36] SUGIHARTO B, MIYATA K, NAKAMOTO H, SASAKAWA H, SUGIYAMA T. Regulation of expression of carbon-assimilating enzymes by nitrogen in maize leaf. Plant Physiology, 1990, 92(4): 963- 969.
(责任编辑 杨鑫浩)
Effect of Nitrogen Application on Carbon and Nitrogen Metabolism of Different Summer Maize Varieties
CHANG JianFeng, DONG PengFei, WANG XiuLing, LIU WeiLing, LI ChaoHai
(College of Agronomy, Henan Agricultural University, Zhengzhou 450002)
【Objective】The study is designed to put forward the physiological indexes of high and stable yield in maize and explore the new management ways by comparing the differences of carbon and nitrogen metabolism of different maize varieties under different nitrogen levels and analyzing its coordination. 【Method】 The study was carried out to compare the differences of yield, the staying-green duration of leaf, nitrogen uptake and translocation, the ratio of C/N, PEP carboxylase and RUBP carboxylase activity by field and pot experiments in Wenxian and Zhengzhou from 2009 to 2011, in which 4 varieties of summer maize, including ZD958, ND108, XD20 and YD2002, were planted at 60 000 plants/hm2under four nitrogen levels of 0, 120, 240, 360 kg·hm-2.【Result】(1) The grain yield of 4 maize varieties increased significantly with nitrogen level increasing. In pot experiments, the yield gap of XD20 between different nitrogen levels was the biggest, and that of ZD958 was the smallest. However, under the field condition, the difference of maize except XD20 between N240 and N360 was not significant. (2) The comparison of nitrogen uptake and translocation during grain-filling stage indicated that the nitrogen in ZD958 vegetative organs could be preferentially transferredto the leaves under low nitrogen condition which contributed to the longer staying-green duration of maize leaf. For ND108, both of nitrogen uptake and translocation were larger, thus its leaf can also stay green longer, but the effective utilization of nitrogen was lower. For XD20, due to lack of mechanism of priority supply to leaves and lower N uptake of under low nitrogen condition, the leaf showed early senescence. For YD2002, because of larger nitrogen transfer amount from vegetative organs to grain under four nitrogen levels and smaller nitrogen absorption in root, the leaf also showed early senescence. (3)During the grain filling stage, the C/N ratio of leaf in ZD958 and XD20 were between YD2002 and ND108 in the condition of normal fertilization. For the mature stage, the grain C/N ratio of ZD958 and XD20 were higher than the other two varieties. (4)At mid filling stage, ratio of PEPCase and RUBPCase of ZD958 was significantly higher than the other three cultivars, that of YD2002 was lower, and that of XD20 and ND108 increased significantly with the increase of nitrogen fertilizer. 【Conclusion】The excellent coordination of carbon and nitrogen metabolism of maize was reflected in the translocation process of carbohydrate and nitrogen. The maize that be able to coordinate the contradiction between grain and vegetative organ demand for carbohydrate and nitrogen, and with a higher PEPCase/RUBPCase, could reach a high and stable yield.
maize; carbon metabolism; nitrogen metabolism; C/N; PEPCase; RUBPCase
2016-07-19;接受日期:2017-02-10
国家现代农业产业技术体系岗位科学家专项(CARS-02-19)
联系方式:苌建峰,E-mail:chjfchina@163.com。通信作者李潮海,E-mail:lichaohai2005@163.com
猜你喜欢
中国化肥信息(2022年9期)2022-11-23
商品与质量(2021年16期)2021-04-23
中国化肥信息(2019年4期)2019-05-31
中国化肥信息(2019年3期)2019-04-25
意林(2018年18期)2018-09-30
中国化肥信息(2017年3期)2017-12-23
作文周刊·小学一年级版(2017年35期)2017-10-18
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
江苏农业科学(2017年1期)2017-02-27
湖北农业科学(2014年16期)2014-10-20
