
祁连山大野口流域典型灌丛植物与土壤中氮磷的化学计量特征①
2017-07-21 05:05赵维俊敬文茂赵永宏4
土壤 2017年3期
赵维俊,敬文茂*,赵永宏4,马 剑
祁连山大野口流域典型灌丛植物与土壤中氮磷的化学计量特征①
赵维俊1,2,3,敬文茂1,2,3*,赵永宏4,马 剑1,2,3
(1 甘肃省祁连山水源涵养林研究院,甘肃张掖 734000;2 甘肃省森林生态与冻土水文水资源重点实验室,甘肃张掖 734000; 3 甘肃张掖生态科学研究院甘肃省祁连山生态科技创新服务平台,甘肃张掖 734000;4 甘肃农业大学林学院,兰州 730070)
通过野外取样与室内分析相结合的方法,对祁连山西水林区的鬼箭锦鸡儿()、吉拉柳()、金露梅()、鲜黄小檗()和甘青锦鸡儿()等5种典型灌丛植物和土壤氮(N)和磷(P)含量进行了测定和分析,旨在阐明植物与土壤中N、P的化学计量特征。结果表明:①5种灌丛植物叶片的N、P含量均显著高于其他器官的N、P含量,N含量的变化范围为15.72 ~ 29.08 g/kg,P含量的变化范围为1.58 ~ 3.47 g/kg;细根和粗根的N含量的变化范围分别为5.11 ~ 12.20 g/kg和4.46 ~ 11.84 g/kg,P含量的变化范围分别为0.87 ~ 1.58 g/kg和0.63 ~ 2.02 g/kg;叶片N∶P值范围为7.00 ~ 9.95,细根和粗根中的N∶P值分别为5.60 ~ 8.97和5.09 ~ 8.84,叶片和根系对N、P元素有着相似的利用格局。②5种灌丛土壤N含量均随土层深度增加逐渐减小,不同土层N含量的变化范围为1.40 ~ 6.80 g/kg,P含量均随土层深度增加较为稳定,不同土层P含量的变化范围为0.45 ~ 0.96 g/kg;N∶P值随土层深度增加其值不断减小,主要取决于土壤N含量。0 ~ 10 cm土层中,吉拉柳、鬼箭锦鸡儿、金露梅、鲜黄小檗和甘青锦鸡儿N∶P值分别为11.02、7.30、7.41、7.40和2.69。③5种灌丛叶片N含量与土壤N含量呈显著或极显著正相关,与土壤P含量无显著相关(甘青锦鸡儿除外);叶片N∶P值与土壤N含量呈显著负相关,而与土壤P含量呈显著正相关;灌丛叶片和土壤N、P含量及化学计量比与其他变量之间均无显著相关。同时研究还发现,祁连山5种典型灌丛生长可能主要受N素限制。
典型灌丛;叶片;土壤;氮磷含量;N∶P;化学计量;祁连山
氮(N)、磷(P)元素是植物体的重要组成部分,是维系植物生命活动所吸收的大量营养元素,也是影响森林生产力的主要限制因素[1],N、P 元素比值是表征植物群落或植物生产力受那种元素限制的指标[2],同时N、P 元素的协同作用影响植物个体的功能运行甚至整个生态系统的进程[3]。因此,植物和土壤N、P化学计量研究逐渐成为生态学研究的热点。由于全球气候变化和人类活动的加剧,山地森林生长环境也受到了严重的影响,诸如N沉降、过度放牧等使得土壤N含量的增加或减少,而土壤P含量较为稳定,土壤N、P的失衡在一定程度上影响了森林结构和功能的稳定性[4-5]。如何阐述N、P元素平衡或元素平衡对生态交互作用的影响?生态化学计量学为解决这一问题提供了切实可行的工具[6]。祁连山区水热条件的垂直差异造就了植被分布的垂直地带性[7],加上祁连山区森林生态系统地处西北高寒山地地带,土壤-植物系统对养分元素的循环具有一定的独特性,阐明不同灌丛群落优势种植物和土壤的N、P化学计量特征,有助于了解不同灌丛群落的功能差异及其对环境变化的适应性。
祁连山区适宜的热量和水分条件孕育了4类典型森林生态系统,包括落叶阔叶林、针阔混交林、常绿针叶林和落叶阔叶灌丛林,其中落叶阔叶灌丛林面积(12 866.49 km2)占祁连山区林地总面积(16 133.01 km2)的79.75%[8],作为祁连山森林生态系统的重要组成部分,在区域生态环境保护和替代能源方面具有非常重要的地位和作用[9-10]。目前,对祁连山灌丛林的研究,多集中在灌丛水文生态功能[9,11]、灌丛生物量[8,12]、灌丛保护和经营管理等方面[13-15],而对灌丛养分特征研究较少。因此,本文以分布在祁连山中段西水林区大野口流域不同垂直植被带上典型灌丛群落的优势种鬼箭锦鸡儿()、吉拉柳()、金露梅()、鲜黄小檗()和甘青锦鸡儿()等5种典型灌丛为研究对象,在测定不同灌丛植物和土壤中全N、全P含量的基础上,分别对植物各器官的N、P化学计量学特征、土壤N、P化学计量学特征以及二者之间关系3个方面进行研究,以揭示不同灌丛群落的土壤-植物养分变化特征,并判断灌丛养分限制因子,为祁连山灌丛植被的退化恢复和科学管理提供依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区地处祁连山腹地西水林区的大野口流域,中心地理坐标100°15′ E,38°31′ N,流域总面积73.32 km2,坡度为32°,海拔为2 590 ~ 4 645 m[16]。该区年平均气温–0.6 ~ 2.0℃,年均日照时数1 893 h,日辐射总量均值为110.28 kW/m2,年均降水量为433.6 mm,年均蒸发量为1 081.7 mm,年均相对湿度为60%,属高寒半干旱山地森林草原气候[17]。流域自然条件复杂,水热条件差异大,受水分和热量制约形成了多种具有明显垂直梯度的植被类型和土壤类型,海拔从低到高,流域植被类型主要有森林草原带、森林灌丛带、亚高山灌丛草甸带、高山草甸带和高山寒漠带,土壤类型主要有山地栗钙土、山地灰褐森林土、山地森林草甸土、亚高山灌丛草甸土和高山草甸土[18],其中山地栗钙土是生长干性灌丛的主要土壤,亚高山灌丛草甸土是湿性灌丛的主要分布带[19]。建群种青海云杉林呈斑块状或条状分布在实验区海拔2 600 ~ 3 300 m 阴坡和半阴坡地带,与阳坡草地犬齿交错分布;灌丛优势种有干性灌丛甘青锦鸡儿、鲜黄小檗和金露梅,主要分布在低海拔地段,湿性灌丛优势种有鬼箭锦鸡儿和吉拉柳等,主要分布在高海拔地段,草本主要有珠牙蓼()、黑穗苔()、苔草()、马苋蒿()和针茅()等。
1.2 研究方法
2014年8月中旬,在不同灌丛植物生长的旺盛期,根据植被调查中普遍应用的典型样地法[20],选择流域分布的鬼箭锦鸡儿、吉拉柳、金露梅、鲜黄小檗和甘青锦鸡儿等5种典型灌丛群落,在每种灌丛群落选择海拔等立地条件相似的坡面建立3个临时调查样地,样地面积大小为20 m × 20 m,其中因甘青锦鸡儿群落密度较小,样地面积大小为30 m × 30 m,共计15个样地。然后,用栅格法将样地划分为5 m × 5 m的样格,调查每个样格内植株的地径、盖度、平均高和生长情况等,样地植物群落概况见表1。
在每个样地随机选取长势相近且较好的植株5 ~ 10株(丛),用铁锹连其根系挖出,尽快带回实验室。同时,在每个样地按梅花型用直径2 cm土钻采集5点的土壤混合成1个土样[21]。采集前先清除土壤表层的枯落物层,然后按0 ~ 10、10 ~ 20、20 ~ 30、30 ~ 40、40 ~ 60 cm分层取样。同一土层取3个重复,将同一土层质量大致相当的土样置于塑料布上,充分混合组成待测样品。每份样品约1 kg,每种灌丛3个样地共3份土壤样品。

表1 不同灌丛植物群落基本情况
采集的植物样品先用清水快速冲洗1 ~ 2次,再用去离子水快速冲洗2 ~ 3次,晾干后用剪刀将叶片、枝条、干、细根和粗根分开,分类混合取部分样置于105℃烘箱杀青2 h,80℃烘干至恒重。然后用粉碎机将同一样品粉碎过100目筛,装袋密封,用于植物N、P含量测定。每种灌丛3个样地,不同植物器官均为3个重复样。采集的土壤样品剔除石砾、植被残根等杂物后,在室温条件下自然风干,研磨过100目筛,装袋备用,用于土壤N、P含量测定。
植物各器官用H2SO4-H2O2溶液消煮,全N采用定氮法测定,全P用钼锑抗比色法测定;土壤采用不包含硝态氮和亚硝态氮的半微量凯式法消化,全N再用全自动凯氏定氮仪进行测定,全P采用钼锑抗比色法测定。各项化学指标的测定均做3次平行,具体测定方法参照《土壤农化分析》[22]。
1.3 数据处理
应用Excel 2007和SPSS16.0对不同灌丛植物和土壤N、P含量化验数据进行前期处理、统计分析和作图。植物和土壤N、P含量和N∶P值(化学计量比采用N、P质量比表示[23])均采用Dixon 检验法消除3次平行测定的偏离值,取其算术平均数作为测定值,不同灌丛的同一器官和同一土层及同一灌丛的不同器官和不同土层的N、P含量、N∶P值差异显著性采用单因素方差分析法(One-Way ANOVA)中的最小显著极差法(LSD)进行多重比较,显著性水平设为=0.05,即差异显著性为<0.05。植物和土壤N、P含量和N∶P值之间相关性分析采用双变量相关分析法(Bivariate Correlations)计算各指标两两之间的Pearson相关系数[24],差异显著性分<0.05和<0.01。在对变量进行单因素方差分析和Pearson相关分析时先对分析变量进行正态分布检验,对服从正态分布检验的变量直接进行分析,否则需要数据转换方可进行下一步分析。本文拟分析的植物和土壤N、P含量和N∶P值经检验均服从正态分布,直接进行单因素方差分析和Pearson相关分析。
2 结果与分析
2.1 植物N、P含量及化学计量比
不同灌丛植物器官叶片、枝、干、细根和粗根的N、P含量各不相同(图1)。叶片的N、P含量显著高于其他器官的N、P含量(<0.05),N含量的变化范围为15.72 ~ 29.08 g/kg,其中甘青锦鸡儿叶片N含量显著高于其他灌丛叶片N含量(<0.05),这与其豆科植物的根瘤菌固氮有很大的关系。同是豆科植物的鬼箭锦鸡儿N含量较低,这可能与其地处高海拔弱的N素利用有关。鬼箭锦鸡儿、吉拉柳和金露梅叶片N含量居中,且差异性不显著(>0.05),鲜黄小檗叶片N含量最低。不同灌丛叶片P含量的变化范围为1.58 ~ 3.47 g/kg,叶片P含量大小及差异显著性与叶片N含量大小及差异显著性相同。
不同灌丛不同器官中干的N、P含量最低,与其他器官枝、细根和粗根的N、P含量差异性不显著(>0.05),干的N含量的变化范围为4.82 ~ 8.16 g/kg,其中甘青锦鸡儿干的N含量最高,金露梅N含量最低,不同灌丛干的N含量差异性不显著(>0.05)。P含量的变化范围为0.49 ~ 1.54 g/kg,其中甘青锦鸡儿P含量显著高于其他灌丛叶片P含量(<0.05),其他4种灌丛P含量次之,且差异性不显著(>0.05)。
不同灌丛器官枝、细根和粗根的N、P含量相差不大,其中细根和粗根N含量差异显著性和P含量差异显著性变化规律相同,鬼箭锦鸡儿和甘青锦鸡儿根系N含量显著高于其他灌丛(<0.05),而P含量仅鬼箭锦鸡儿P含量显著高于其他灌丛(<0.05)。细根和粗根的N含量的变化范围分别为5.11 ~ 12.20 g/kg和4.46 ~ 11.84 g/kg,P含量的变化范围分别为0.87 ~ 1.58 g/kg和0.63 ~ 2.02 g/kg。
不同灌丛植物的N∶P值也各不相同(图1),其中叶片、细根和粗根的N∶P值在不同灌丛较为接近,鬼箭锦鸡儿、吉拉柳、金露梅、鲜黄小檗和甘青锦鸡儿叶片的N∶P值范围为7.00 ~ 9.95,相应根系(细根和粗根)中的N∶P值分别为5.60 ~ 8.97和5.09 ~ 8.84,而且不同灌丛叶片和细根、粗根的N∶P值之间差异性不显著(<0.05),叶片、根系作为植物体最为活跃的器官且对N、P元素有着相似的利用格局。枝和干的N∶P值在不同灌丛变化较大,枝的N∶P值范围为6.18 ~ 11.79,干的N∶P值范围为4.96 ~ 16.68。
2.2 土壤N、P含量及化学计量比
不同灌丛林地土壤不同土层N、P含量各不相同(图2),随土层深度的增加,不同灌丛土壤N含量逐渐减小,递减程度大小在不同土层各不相同,原因是土壤N素主要来源于枯落物的归还,N素首先在土壤表层富集,然后随大气降水等介质向下层进行迁移扩散。其中,0 ~ 10 cm土层中N含量的变化范围为2.03 ~ 6.80 g/kg,40 ~ 60 cm土层中N含量的变化范围为1.40 ~ 5.95 g/kg,吉拉柳和金露梅灌丛土壤N含量较为接近,其含量高于其他灌丛土壤N含量;甘青锦鸡儿和鲜黄小檗灌丛土壤氮含量大小居中;鬼箭锦鸡儿土壤氮含量较小。P含量随土层深度增加没有明显的变化规律且含量较为稳定,大多土层间的差异性不显著(>0.05),主要与岩石的风化有关。不同土层P含量的变化范围为0.45 ~ 0.90 g/kg,金露梅土壤P含量最高,鬼箭锦鸡儿土壤P含量最低。
因土壤N含量随土层深度的增加其含量不断减小,而P含量较为稳定(图2),所以土壤N∶P值随土层深度的增加而降低,但不同土层N∶P值差异显著性各不相同,其中鬼箭锦鸡儿、金露梅土壤N∶P值在0 ~ 30 cm土层范围内稳定,在30 cm以下土层明显降低并趋于稳定;吉拉柳土壤N∶P值在10 ~ 20 cm土层明显减小,20 cm以下稳定不变,鲜黄小檗和甘青锦鸡儿土壤分别在20 cm和40 cm以下土层明显减小,然后明显不变,呈现浅层土壤N∶P值高于深层土壤N∶P值的格局。在0 ~ 10 cm土层中,吉拉柳土壤的N∶P值最大,为11.02,鬼箭锦鸡儿、金露梅和鲜黄小檗土壤的N∶P值较为接近,分别为7.30、7.41和7.40,甘青锦鸡儿土壤仅为2.69。
2.3 植物与土壤N、P含量和化学计量比之间的关系
在生态化学计量学研究中,通常用植物叶片和表层土壤N、P比值研究生态系统化学元素平衡的科学问题[25-27]。因此,本研究对不同灌丛器官叶片N、P含量及其计量比和0 ~ 10 cm土层土壤N、P含量及其化学计量比进行了相关性分析(表2)。从表2可以看出,5种灌丛植物叶片N含量与土壤N含量均呈显著或极显著正相关,灌丛叶片P含量与土壤P含量相关性均不显著(<0.05),除甘青锦鸡儿叶片P含量与土壤P含量之间呈显著相关性(<0.05)外。灌丛叶片N∶P值与土壤N含量存在显著的负相关(<0.05),而叶片N∶P值与土壤P含量存在显著的正相关(<0.05),不同灌丛其他变量之间均无显著相关性(>0.05)。
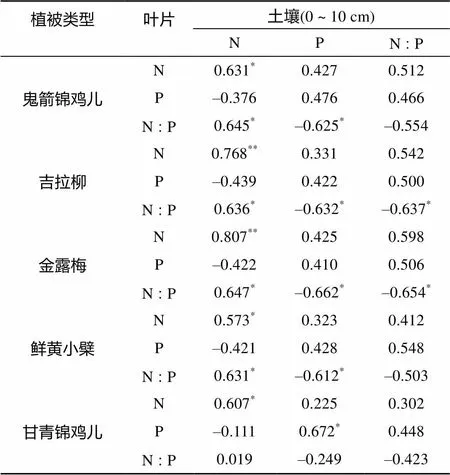
表2 不同灌丛叶片和土壤N、P含量及化学计量比之间的相关性
注:* 表示在<0.05水平显著相关;** 表示在<0.01水平显著相关。
3 讨论与结论
3.1 不同灌丛植物N、P化学计量学特征
本研究表明,祁连山5种典型灌丛在生长旺盛期,除吉拉柳和甘青锦鸡儿叶片N含量稍高于全国平均水平的(20.2 ± 8.4) mg/g[28],也稍高于全球平均水平的(20.6 ± 12.2) mg/g[24],其他3种灌丛叶片N含量均低于全国水平和全球水平[28-29]。灌丛叶片P含量均大于全国平均水平(1.46 ± 0.99) mg/g[31],除鲜黄小檗灌丛(1.58 mg/g)P含量较低外,其他灌丛均高于全球平均水平的(1.99 ± 1.49) mg/g[30]。说明祁连山灌丛植物叶片N含量相对较低,而P含量相对较高。灌丛叶片中,豆科植物甘青锦鸡儿N、P含量最高,小檗科植物鲜黄小檗N、P含量最低,同是豆科植物的鬼箭锦鸡儿和柳科植物吉拉柳、蔷薇科植物金露梅N、P含量居中,表明祁连山不同灌丛植物对N、P含量的需求差别较大,体现了植物的选择性吸收和不同科植物间化学元素分布的不均一性[31]。不同灌丛N、P养分在不同器官的分配格局与其所在的海拔、立地因子、养分有效性等环境条件和其生理特征、生活史特性、自身生长型等物种特点有关,即环境和物种系统发育共同作用的结果[32]。本研究中,吉拉柳和金露梅这两种灌丛地处中海拔地段的阴坡和半阴坡,适宜的土壤水分和温度有利于土壤有机质的积累,可供吸收的养分相对较多;甘青锦鸡儿和鲜黄小檗灌丛地处低海拔地段,坡度较大,土壤水分含量较低,植被密度较低,少量的枯落物又因大气降水和放牧踩踏等原因不容易积累在林地地面,养分积累少;鬼箭锦鸡儿灌丛地处高海拔地段,土层较薄,下面主要分布砂岩,土壤贫瘠,加上鬼箭锦鸡儿群落生产力很小,养分含量低。
植物叶片的N、P含量和N∶P临界值常被用来评判不同地区不同物种受到哪种元素的限制[33]。Güsewell和Koerselman[34]研究发现,当陆地植物的叶片N∶P<10时,植被的生长主要受N限制,而N∶P>20时,植被的生长主要受P限制,介于两者中间表明受到N、P的共同限制。本研究中,不同灌丛植物叶片N∶P值范围为7.00 ~ 9.73,其值均小于中国753个物种叶片平均水平(16.3)[28]和全球平均水平(12.7)[30],而且5种灌丛叶片N∶P值均低于10。因此,较低的N含量和较高的P含量及较低的N∶P值在一定程度上说明祁连山灌丛生长更易受N素限制,也说明了退化灌丛的群落发育与植被生产力主要受N素的限制,也验证了高纬度、高海拔地区植物生长更易受N限制[30]。叶片和根系作为植物体营养代谢和吸收的输出端和输入端,二者之间实现动态平衡方可满足植物体的生长需要。研究发现陆生植物根系与叶片N、P含量相比,根系N、P浓度比叶片低,而且根系N∶P值与叶片的N∶P值基本相似[35-36],本研究的结果也证明了这一点,而且不同灌丛细根和粗根的N∶P值差异性不显著,叶片和根系的N∶P值可以作为灌丛生长状况的一个评价指标。
3.2 不同灌丛土壤N、P化学计量学特征
本研究表明,5种灌丛土壤中N含量在剖面上呈“倒金字塔”的分布,P含量在剖面上呈“圆柱体”分布模式。在0 ~ 10 cm土层中,5种灌丛土壤N含量均大于全国平均水平1.88 mg/g[1],这与研究区地处亚高山地带的山地森林草原气候有关。低温潮湿环境使得有机层微生物活性较低,枯落物分解缓慢,有机质不断得到累积,但也不排除牲畜排泄物含有大量N素,增加了土壤中N素含量。尽管土壤N含量较为丰富,结合叶片N∶P值判断,说明可供灌丛吸收的有效N含量可能较低。除金露梅和甘青锦鸡儿土壤P含量稍高于全国平均水平的0.78 mg/g外,其他灌丛土壤均低于全国平均水平[37],5种灌丛土壤P含量均远低于全球平均水平的2.8 mg/g[28]。总体来看,研究区灌丛土壤P含量较低,这可能与5种灌丛群落“自肥”作用较弱和岩石风化缓慢有很大的关系。
土壤N∶P值可以作为养分限制类型的有效预测指标[38],与全球不同生态系统土壤N∶P值的平均水平13和全国土壤N∶P值的平均水平9.3相比[35,39],研究区不同灌丛土壤N∶P值均较低(除吉拉柳土壤高于全国平均值),较低的N∶P值指示5种灌丛生长受N素限制,尽管豆科植物甘青锦鸡儿和鬼箭锦鸡儿虽然具有固氮作用,但是低海拔的甘青锦鸡儿和高海拔的鬼箭锦鸡儿来自于有机质的养分积累很少,可供植物生长的氮素较少。总之,5种灌丛群落土壤P素含量较低,但相对N素而言,可供灌丛吸收的有效P含量较高,这是因为在高海拔和高纬度地区的N可利用性比P可利用性更容易受到低温的影响[31],是土壤对环境长期适应形成的策略。不同灌丛土壤N∶P值均随土层深度增加其值不断减小,土壤N∶P值对土壤深度的响应各不相同,沿土壤剖面自上而下呈高-低型分布,该研究结果与青藏高原放牧高寒草甸土壤N∶P值变化规律一致[40],但与Tian等[37]得出的土壤0 ~ 10 cm的N∶P值显著高于其他土层的结论有所不同,造成这一现象可能与研究区土壤的分解程度和贮存多年的腐殖质累积有一定的关系[41]。祁连山高山灌丛是我国西北的重要生态屏障,但因生长环境恶劣,灌丛生长缓慢,养分含量相对稳定,受自然干扰特别是人为干扰后其生态自我修复能力较差,因此要加强研究区灌丛保护。
3.3 植物与土壤N、P化学计量学特征的关系
植物体中N、P元素主要来源于根系对土壤养分的吸收,其养分含量的高低与土壤养分含量密切相关[42]。多数研究表明,叶片N、P元素含量与土壤N、P元素含量密切相关[32,43]。刘佳庆等[44]对长白山牛皮杜鹃养分计量特征研究表明,其叶片与1年生枝条中的N、P含量与土壤中的N、P含量均呈极显著和显著正相关。本研究中,5种灌丛植物叶片N浓度和土壤全N含量之间均呈显著或极显著正相关,而植物叶片P浓度和土壤全P含量之间的相关性不显著(除甘青锦鸡儿植物P和土壤P呈显著相关性外),说明灌木生长受到土壤N限制,而不受土壤P限制[45],N、P元素之间协同作用能力较差,这与前面植物叶片和土壤N∶P值的判断一致。同时,叶片N∶P值与土壤N含量显著正相关及叶片N∶P值与土壤P含量显著负相关,表明土壤在一定程度上决定了植物对养分的吸收,从另一角度上验证了土壤N对灌丛生长的限制作用。这与同一研究流域青海云杉林生长所需的P素含量相对N素含量少具有不同的养分吸收特点,从而使得生态化学计量内稳性存在明显差异,这可能也是该流域乔木和灌木两种植被类型分布差异的原因之一[46]。因此,影响祁连山区灌丛生长和发育的瓶颈是N元素的匮乏,在对退化灌丛的植被恢复中,应考虑到灌丛生长受N素的限制超过了P素,从而促进植物与土壤中N、P养分良性循环。
但一次实验得到的相关数据能不能代表研究区不同灌丛植物不同器官和土壤N、P含量?植物和土壤N、P含量及生态化学计量学特征究竟怎样?这些还有待进一步研究。另外,受多种因素的影响,研究区不同灌丛植物和土壤N、P阈值也有待深入研究。
[1] 卢同平, 史正涛, 牛洁, 等. 我国陆生生态化学计量学应用研究进展与展望[J]. 土壤, 2016, 48(1): 29–35
[2] 张向茹, 马露莎, 陈亚南, 等. 黄土高原不同纬度下刺槐林土壤生态化学计量学特征研究[J]. 土壤学报, 2013, 50(4): 818–825
[3] 罗由林, 李启权, 王昌全, 等. 近30年川中丘陵区不同土地利用方式土壤碳氮磷生态化学计量特征变化[J]. 土壤, 2016, 48(4): 726–733
[4] 王晶苑, 张心昱, 温学发, 等. 氮沉降对森林土壤有机质和凋落物分解的影响及其微生物学机制[J]. 生态学报, 2013, 33(5): 1337–1346
[5] 红梅, 余娜, 赵红儒, 等. 放牧对土壤碳、氮含量空间变异的影响[J]. 草业科学, 2013, 30(4): 521–527
[6] 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征[J]. 生态学报, 2008, 28(8): 3937–3946
[7] 王金叶, 常学向, 葛双兰, 等. 祁连山(北坡)水热状况与植被垂直分布[J]. 西北林学院学报, 2001, 16(增): 1–3
[8] 姜桦. 祁连山森林生态系统固碳参量的遥感估算[D]. 兰州: 兰州交通大学, 2013
[9] 张平, 刘贤德, 张学龙, 等. 祁连山林草复合流域灌木林水文生态功能研究[J]. 干旱区地理, 2013, 36(5): 922– 929
[10] 王学福. 灌木林在祁连山区的作用及其发展策略研究[J]. 甘肃林业科技, 2005, 30(2): 32–35
[11] 金铭, 张学龙, 刘贤德, 等. 祁连山林草复合流域灌木林土壤水文效应研究[J]. 水土保持学报, 2009, 23(1): 169–172
[12] 雷蕾. 祁连山高山灌丛生物量沿海拔梯度分配特征研究[D]. 兰州: 甘肃农业大学, 2012
[13] 马永骏, 王金叶, 刘兴明, 等. 祁连山保护区森林生态系统现状与保护对策[J]. 西北林学院学报, 2005, 20(4): 5–8
[14] 倪自银, 汪有奎, 杨全生, 等. 祁连山自然保护区灌木林灾害及防治对策[J]. 水土保持研究, 2005,12(2): 107–110
[15] 韩涛. 用TM资料对祁连山部分地区进行针叶林、灌木林分类研究[J]. 遥感技术与应用, 2002, 17(6): 317–321
[16] 张彦丽, 李丑荣, 王秀琴, 等. 基于WorldView-2制备大野口流域高分辨率DEM及精度分析[J]. 遥感技术与应用, 2013, 28(3): 431–436
[17] 李效雄, 刘贤德, 赵维俊. 祁连山大野口流域青海云杉种群结构和空间分布格局[J]. 干旱区地理, 2012, 35(6): 960–967
[18] 姜林. 祁连山西水林区典型土壤类型发生特性及系统分类研究[D]. 陕西杨凌: 西北农林科技大学, 2012
[19] 王金叶, 王艺林, 金博文, 等. 干旱半干旱区山地森林的水分调节功能[J]. 林业科学, 2011, 37(5): 120–125
[20] 宋永昌. 植被生态学[M]. 上海: 华东师范大学出版社, 2001
[21] 国家林业局. LY/T 1952—2011森林生态系统长期定位观测方法[S]. 南京: 凤凰出版社, 2011
[22] 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000
[23] 阎恩荣, 王希华, 郭明, 等. 浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C:N:P化学计量特征[J]. 植物生态学报, 2010, 34(1): 48–57
[24] 张文彤. SPSS 统计分析高级教程[M]. 北京: 高等教育出版社, 2004: 213–234
[25] Elser J J, Fagan W F, Denno R F, et al. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature, 2000, 408(6812): 578–580
[26] Güsewell S. N: P ratios in terrestrial plants: variation and functional significance[J]. New Phytologist, 2004,164( 2) : 243–266
[27] Townsend A R, Cleveland C C, Asner G P, et al. Controls over foliar N:P ratios in tropical rain forests[J]. Ecology, 2007, 88(1): 107–118
[28] Han W X, Fang J Y, Guo D L, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168:377–385
[29] 杜虎, 彭晚霞, 宋同清, 等. 桂北喀斯特峰丛洼地植物群落特征及其与土壤的耦合关系[J]. 植物生态学报, 2013, 37(3): 197–208
[30] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004,101(30): 11001–11006
[31] 张珂, 陈永乐, 高艳红, 等. 阿拉善荒漠典型植物功能群氮、磷化学计量特征[J]. 中国沙漠, 2014, 34(5): 1261–1267
[32] Aerts R, Chapin F S. The mineral nutrition of wild plants revisited:a re-evaluation of processes and patterns [J]. Advances Ecological Research, 2000, 30(C): 1–67
[33] Zhang L X, Bai Y F, Han X G. Differential responses of N:P stoichiometry ofandto N additions in a steppe ecosystem in Nei Mongol[J]. Acta Botanica Sinica, 2004,46(3): 259–270
[34] Güsewell S, Koerselman W. Variation in nitrogen and phosphorus concentrations of wetland plants[J]. Perspective in Ecology, Evolution and Systematics, 2002, 5(1): 37–61
[35] Yuan Z Y, Chen H Y, Reich P B. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus[J]. Nature Communications, 2011, 2: 344
[36] Wu G L, Li W, Shi Z H, et al. Aboveground dominant functional group predicts belowground properties in an alpine grassland community of western China[J]. Journal of Soils and Sediments, 2011,11(6):1011–1019
[37] Tian H Q, Chen G S, Zhang C, et al. Pattern and variation of C:N:P ratios in China’s soils: A synthesis of observational data[J]. Biogeochemistry, 2009, 98(1/2/3): 139–151
[38] 俞月凤, 彭晚霞, 宋同清, 等. 喀斯特峰丛洼地不同森林类型植物和土壤C、N、P化学计量特征[J]. 应用生态学报, 2014, 25(4): 947–954
[39] Cleveland C C, Liptzin D. C:N:P stoichiometry in soil:Is there a “Redfield ratio”for the microbial biomass? [J]. Biogeochemistry, 2007, 85: 235–252
[40] 张法伟, 李英年, 汪诗平, 等. 青藏高原高寒草甸土壤有机质、全氮和全磷含量对不同土地利用格局的响应[J]. 中国农业气象, 2009, 30(3): 323–326
[41] Callesen I, Raulund-Rasmussen K, Westman C J, et al. Nitrogen pools and C:N ratios in well-drained forest soils related to climate change and soil texture[J]. Boreal Environment Research, 2007, 12: 681–692
[42] 郑淑霞, 上官周平. 黄土高原地区植物叶片养分组成的空间分布格局[J]. 自然科学进展, 2006, 16(8): 965–973
[43] Hedin L O. Global organization of terrestrial plant-nutrient interactions[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 10849–10850
[44] 刘佳庆, 孟莹莹, 包也, 等. 长白山林线植物岳桦和牛皮杜鹃养分化学计量特征及其影响因子[J]. 生态学杂志, 2013, 32(12): 3117–3124
[45] Cheplich G P. Population Biology of Grasses[M].Cambridge: Cambridge University Press,1998: 155–181
[46] 赵维俊, 刘贤德, 金铭, 等. 祁连山青海云杉林叶片-枯落物-土壤的碳氮磷生态化学计量特征[J]. 土壤学报, 2016, 53(2): 201–213
Nitrogen and Phosphorus Stoichiometry of Plants and Soils of Typical Shrubs in Dayekou Basin of Qilian Mountains
ZHAO Weijun1,2,3, JING Wenmao1,2,3*, ZHAO Yonghong4, MA Jian1,2,3
(1 Academy of Water Resources Conservation Forests in Qilian Mountains of Gansu Province, Zhangye, Gansu 734000, China; 2 Gansu Province Key Laboratory of Forest Ecology and Forzen-soil Hydrology and Water Resources, Zhangye, Gansu 734000, China; 3 Gansu Province Science and Technology Innovation Service Platform of Ecology in Qilian Mountains, Zhangye Academy of Ecology Science, Zhangye, Ganshu 734000, China; 4 College of Forest, Gansu Agricultural University, Lanzhou 730070, China)
The N and P contents of plants and soils of five typical shrub communities (,,,and) in the Xishui forest zone of the Qilian Mountains were measured and analyzed by field investigation and laboratory analysis, and the stoichiometry of N and P in the plants and soils of five shrubs were also clarified. The results showed that: 1) N and P contents in the leaves were significantly higher than those in the other organs of five shrubs, N and P contents of the leaves ranged from 15.72 to 29.08 g/kg and 1.58 to 3.47 g/kg, respectively. N contents in fine roots and thick roots ranged from 5.11 to 12.20 g/kg and 4.46 to 11.84 g/kg, respectively, P contents in fine roots and thick roots ranged from 0.87 to 1.58 g/kg and 0.63 to 2.02 g/kg, respectively. N∶P ratio of leaves ranged from 7.00 to 9.95, N:P ratios of fine roots and thick roots ranged from 5.60 to 8.97 and 5.09 to 8.84, respectively. N and P in leaves and roots had similar use pattern. 2) N content in soil increased with the decrease of soil depth in five shrubs, N and P contents in soils at different layers ranged from 1.40 to 6.80 g/kg and 0.45 to 0.96 g/kg, respectively. N∶P ratio, mainly dependent on N content, decreased with the increase of soil depth. N∶P ratios of,,andwere 11.02, 7.30, 7.41, 7.40 and 2.69, respectively at 0-10 cm soil layers. 3) Significant or extremely significant positive correlations appeared between N contents in the leaves and in the soils, while no significant correlations occurred between P contents in the leaves and in the soils (except). N∶P ratio of the leaves was significantly negatively correlated with soil N content while was significantly positively correlated with soil P content. N and P contents and stoichiometric ratios had no significant correlations with other variables of leaves and soils. Furthermore, the study also found that nitrogen may be a limiting factor for the growth and development of five shrubs in the Qilian Mountains.
Typical shrubs; Leaf; Soil; Contents of N and P; N∶P ratio; Stoichiometry; Qilian Mountains
10.13758/j.cnki.tr.2017.03.021
Q948.1
A
甘肃省青年科技基金项目(145RJYG318)、科技基础性工作专项(2014FY120700 )和甘肃省森林生态与冻土水文水资源重点实验室项目资助。
(maodanjing@126.com)
赵维俊(1981—),男,甘肃靖远人,博士,副研究员,主要从事森林与土壤生态研究。E-mail:zhaoweijun1019@126.com
猜你喜欢
水土保持学报(2022年5期)2022-10-10
种子(2022年5期)2022-06-27
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
延边大学学报(自然科学版)(2021年1期)2021-04-27
读者·原创版(2021年2期)2021-02-26
安徽农学通报(2020年6期)2020-04-17
农民致富之友(2018年8期)2018-06-27
现代园艺(2017年13期)2018-01-19
华人时刊(2016年17期)2016-04-05
黄河之声(2016年20期)2016-02-21
