
杨絮纤维及柳絮纤维的三级红外光谱研究
2017-07-19 10:36于宏伟赵婷婷杨晓雪侯美岩张琳曼唐甜甜
纺织科学与工程学报 2017年3期
于宏伟,赵婷婷,杨晓雪,侯美岩,张 静,张琳曼,唐甜甜
(石家庄学院化工学院,河北石家庄 050035)
杨絮纤维及柳絮纤维的三级红外光谱研究
于宏伟,赵婷婷,杨晓雪,侯美岩,张 静,张琳曼,唐甜甜
(石家庄学院化工学院,河北石家庄 050035)
采用傅里叶变换衰减全反射红外光谱技术(ATR-FTIR),分别研究了杨絮纤维和柳絮纤维的三级红外光谱(包括:一维红外光谱,二阶导数红外光谱和同步二维红外光谱)。实验发现: 杨絮纤维和柳絮纤维同时存在着 CH2不对称伸缩振动模式(νasCH2),CH 伸缩振动模式(νCH),CH2对称伸缩振动模式(νsCH2),C=O 伸缩振动模式(νC=O),酰胺峰 Ⅰ(νC=O)模式,酰胺峰 Ⅱ(δN-H+ νC-N)模式,OH 变形振动模式(δOH)和 C-O 伸缩振动模式(νC-O)等。研究发现:在 3000 cm-1~2800 cm-1、1800 cm-1~1550 cm-1、1450 cm-1~1400 cm-1、1350 cm-1~1300 cm-1和 1100 cm-1~1000 cm-1频率区间内,采用传统的红外光谱技术(包括:一维红外光谱和二阶导数红外光谱)不能有效的区分杨絮纤维和柳絮纤维,而同步二维红外光谱可以清晰的区分杨絮纤维和柳絮纤维。本项研究拓展了三级红外光谱技术在天然纺织材料的研究范围。
杨絮纤维 柳絮纤维 三级红外光谱
杨絮纤维(Populus Tomentosa Fibre)和柳絮纤维(Catkin Fibre)是一类常见天然植物纤维。其中杨絮纤维长度短,中空度大,吸油拒水,可以用纱网做骨架制作吸油毡,且加工过程简单,收集效果好[1]。杨絮纤维和柳絮纤维结构相似,均为 α-纤维素[2-10],但相关结构研究却少见报道,红外光谱法由于具有快捷,分辨率高的优点,近年来成为研究天然纤维结构的新方法[11-12]。其中透射红外光谱法(Transmission-FTIR)通常要采用溴化钾压片,会破坏天然植物纤维的结构。而衰减全反射红外光谱法(ATR-FTIR)则是一种较为新型的红外光谱技术,不需要对样品进行任何处理。因此本研究采用 ATR-FTIR 技术,以杨絮纤维和柳絮纤维为研究对象,系统研究了其三级红外光谱(包括:一维红外光谱,二阶导数红外光谱和同步二维红外光谱)。
1 实验
1.1 材料
杨絮和柳絮(其中杨絮采自于石家庄学院北校区化工楼前的白杨树,而柳絮采自于石家庄学院北校区化工楼前的垂柳树,其品种经过生物学谷维娜博士鉴定)
1.2 仪器
红外光谱仪(Spectrum 100 型号,美国 PE 公司生产); ATR-FTIR 变温附件(Golden Gate 型号,英国 Specac 公司生产);ATR-FTIR 变温控件(WEST 6100+型号,英国 Specac 公司生产)。
1.3 方法
1.3.1 实验条件
以空气为背景,每次对于信号进行 8 次扫描累加,测定频率范围3000 cm-1~600 cm-1;测温范围 293 K~393 K(变温步长 10 K)。
1.3.2 数据获得及处理
一维及二阶导数红外光谱的数据获得采用 Spectrum v 6.3.5软件;同步二维红外光谱数据采用 TD Versin 4.2 软件。
2 结果与讨论

A 杨絮纤维和柳絮纤维一维红外光谱

B 杨絮纤维和柳絮纤维二阶导数红外光谱
图1 杨絮纤维和柳絮纤维一维及二阶导数红外光谱(3000 cm-1~600 cm-1)
在 3000 cm-1~600 cm-1频率范围内,同时研究了杨絮纤维和柳絮纤维的一维及二阶导数红外光谱(图 1)发现:杨絮纤维和柳絮纤维的结构非常类似,而其主要特征官能团的红外吸收频率主要集中在 3000 cm-1~2800 cm-1,1800 cm-1~1550 cm-1,1450 cm-1~1400 cm-1,1350 cm-1~1300 cm-1和 1100 cm-1~1000 cm-1等五个频率区间,因此本文主要在这五个频率区间开展了杨絮纤维和柳絮纤维的三级红外光谱的研究。
2.1 3000 cm-1~2800 cm-1频率范围内杨絮纤维和柳絮纤维的三级红外光谱的研究
2.1.1 3000 cm-1~2800 cm-1频率范围内杨絮纤维和柳絮纤维的一维及二阶导数红外光谱的研究
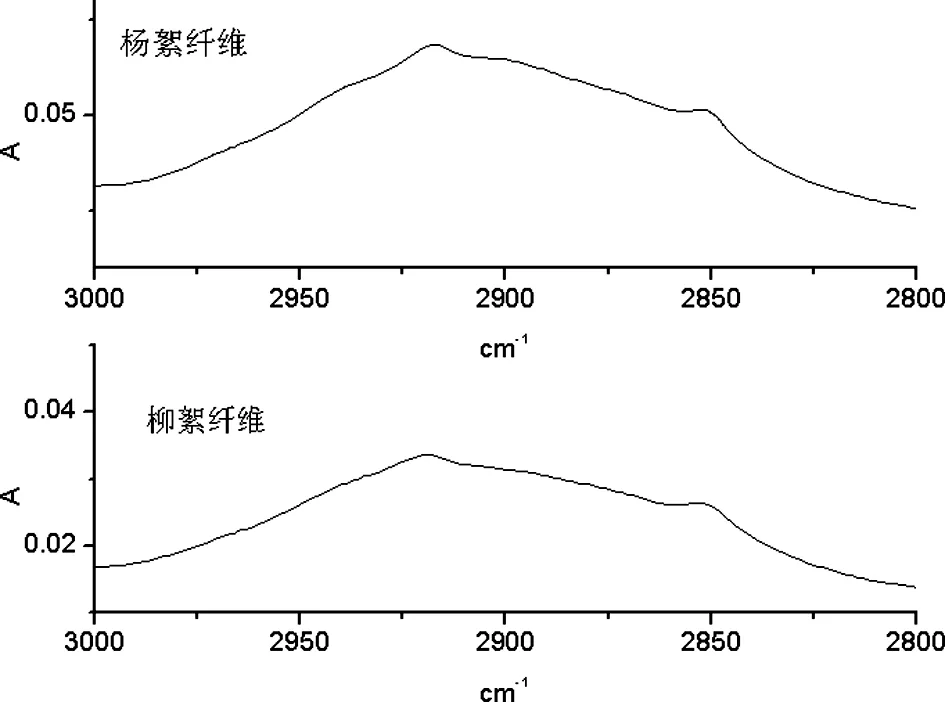
A 杨絮纤维和柳絮纤维一维红外光谱

B 杨絮纤维和柳絮纤维二阶导数红外光谱
图2 杨絮纤维和柳絮纤维一维及二阶导数红外光谱(3000 cm-1~2800 cm-1)
在 3000 cm-1~2800 cm-1频率范围内首先开展了杨絮纤维和柳絮纤维的一维红外光谱的研究(图 2A)。其中 2920 cm-1频率处吸收强度较弱的红外吸收峰归属于CH2不对称伸缩振动模式(νasCH2),而 2850 cm-1频率处吸收强度较弱的红外吸收峰则归属于CH2对称伸缩振动模式(νsCH2)。进一步开展了杨絮纤维及柳絮纤维的二阶导数红外光谱研究(图 2B),其分辨率有了显著的提高,其中 2920 cm-1频率处较强红外吸收峰归属于 νasCH2,而 2850 cm-1频率处较强红外吸收峰归属于 νsCH2,相关红外光谱数据见表 1。
2.1.2 3000 cm-1~2800 cm-1频率范围内杨絮纤维和柳絮纤维的同步二维红外光谱的研究
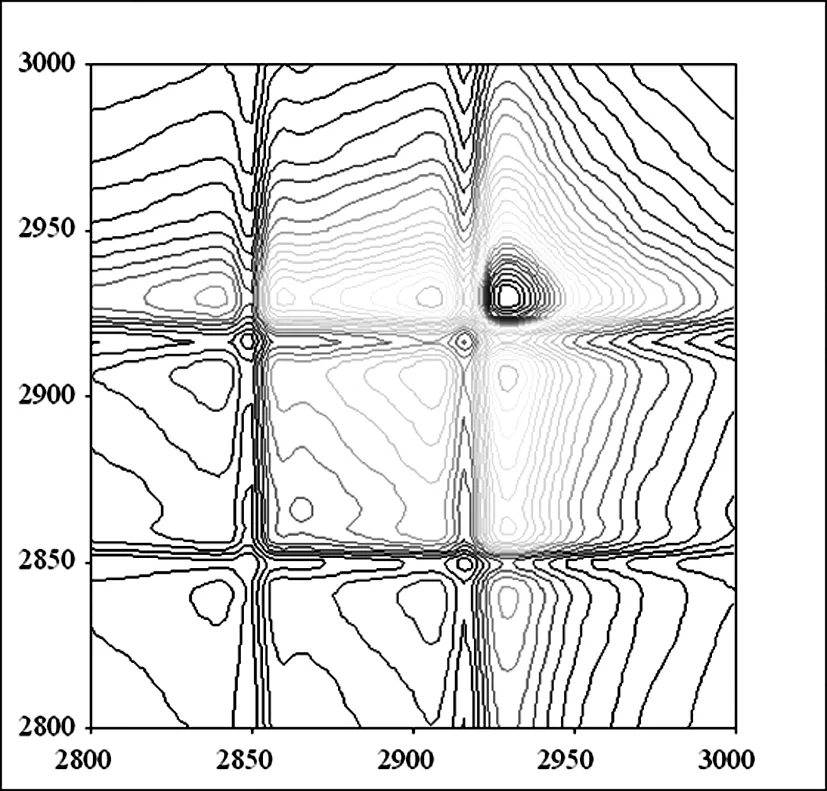
A 杨絮纤维同步二维红外光谱
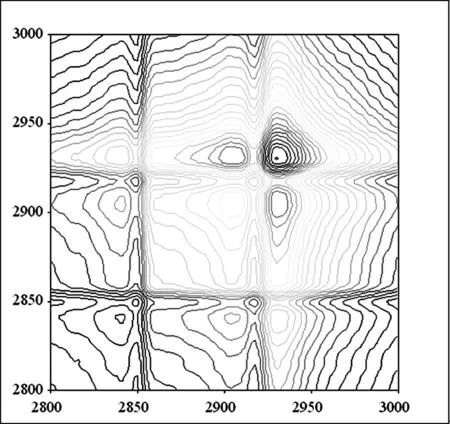
B 柳絮纤维同步二维红外光谱
图3 杨絮纤维和柳絮纤维的同步二维红外光谱(3000 cm-1~2800 cm-1)
在3000 cm-1~2800 cm-1的频率范围内,开展了杨絮纤维的同步二维红外光谱的研究(图 3A)[13-17]。同步二维红外光谱的分辨率则优于相应的一维及二阶导数红外光谱(图 2)。在(2930 cm-1,2930 cm-1),(2918 cm-1,2918 cm-1),(2905 cm-1,2905 cm-1)和(2850 cm-1,2850 cm-1)频率处发现四个自动峰。其中(2930 cm-1,2930 cm-1)和(2918 cm-1,2918 cm-1)频率处的自动峰归属于 νasCH2;(2850 cm-1,2850 cm-1)频率处的自动峰则归属于 νsCH2;而(2905 cm-1,2905 cm-1)频率处的自动峰则归属于 C-H 伸缩振动模式(νCH)。其中(2930 cm-1,2930 cm-1)频率处的自动峰强度最大,这说明该频率对应的官能团对于温度变化比较敏感。而进一步研究柳絮纤维的同步二维红外光谱(图 3B),则得到了同样的光谱信息,相关数据见表 1。
2.2 1800 cm-1~1550 cm-1频率范围内杨絮纤维和柳絮纤维的三级红外光谱的研究
2.2.1 1800 cm-1~1550 cm-1频率范围内杨絮纤维和柳絮纤维的一维及二阶导数红外光谱的研究

A 杨絮纤维和柳絮纤维一维红外光谱
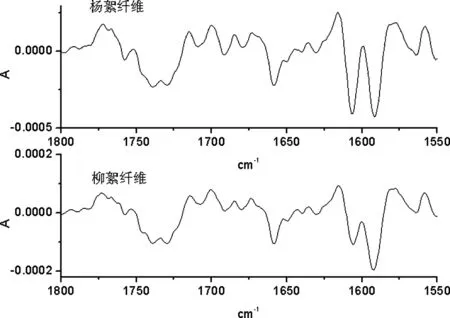
B 杨絮纤维和柳絮纤维二阶导数红外光谱
图4 杨絮纤维和柳絮纤维一维及二阶导数红外光谱(1800 cm-1~1550 cm-1)
在 1800 cm-1~1550 cm-1频率范围内首先开展了杨絮纤维和柳絮纤维的一维红外光谱的研究(图 4A)。其中 1740 cm-1频率处较宽的红外吸收峰归属于油脂的羰基伸缩振动模式(νC=O)。而 1600 cm-1频率处较宽的红外吸收峰归属于蛋白质酰胺峰 Ⅱ(δN-H+ νC-N)模式;进一步开展了杨絮纤维及柳絮纤维的二阶导数红外光谱研究(图 4B),其分辨率有了显著的提高,其中 1740 cm-1频率处较宽的红外吸收峰裂分为 1735 cm-1和 1745 cm-1的两个红外吸收峰。而 1660 cm-1,1610 cm-1和 1580 cm-1频率处发现了三个新的红外吸收峰,其中 1660 cm-1频率处的红外吸收峰归属于蛋白质酰胺峰 Ⅰ(νC=O)模式,而 1610 cm-1和 1580 cm-1频率处的红外吸收峰则归属于蛋白质酰胺峰 Ⅱ(δN-H+ νC-N)模式,这主要是因为杨絮纤维和柳絮纤维含有种子,种子里含有少量的油脂及蛋白质,而相关红外光谱数据见表 1。
2.2.2 1800 cm-1~1550 cm-1频率范围内杨絮纤维和柳絮纤维的同步二维红外光谱的研究

A 杨絮纤维同步二维红外光谱

B 柳絮纤维同步二维红外光谱
图5 杨絮纤维和柳絮纤维的同步二维红外光谱(1800 cm-1~1550 cm-1)
在 1800 cm-1~1550 cm-1的频率范围内,开展了杨絮纤维的同步二维红外光谱的研究(图 5A)。在(1750 cm-1,1750 cm-1),(1620 cm-1,1620 cm-1)和(1580 cm-1,1580 cm-1)频率附近同时发现自动峰,其中(1750 cm-1,1750 cm-1)频率处自动峰强度最大,这说明杨絮纤维 1750 cm-1对应的红外吸收峰对于温度变化比较敏感;此外在(1580 cm-1,1610 cm-1),(1580 cm-1,1750 cm-1)和(1610 cm-1,1750 cm-1)频率附近发现清晰的交叉峰,这说明杨絮纤维 1580 cm-1,1610 cm-1和 1750 cm-1频率对应的官能团之间存在着较强的分子内相互作用。而进一步研究柳絮纤维的同步二维红外光谱(图 5B),在(1740 cm-1,1740 cm-1)和(1580 cm-1,1580 cm-1)频率附近同时发现自动峰,而其中(1750 cm-1,1750 cm-1)频率处自动峰强度最大,同样说明柳絮纤维 1750 cm-1对应的红外吸收峰对于温度变化比较敏感;此外在(1580 cm-1,1750 cm-1)频率附近发现清晰的交叉峰,则进一步证明柳絮纤维 1580 cm-1和 1750 cm-1频率对应的官能团之间存在着较强的分子内相互作用,相关红外光谱数据见表 1。
2.3 1450 cm-1~1400 cm-1频率范围内杨絮纤维和柳絮纤维的三级红外光谱的研究
2.3.1 1450 cm-1~1400 cm-1频率范围内杨絮纤维和柳絮纤维的一维及二阶导数红外光谱的研究
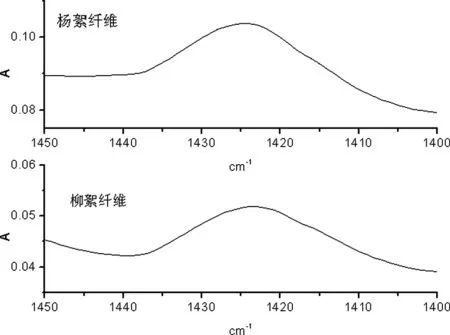
A 杨絮纤维和柳絮纤维一维红外光谱
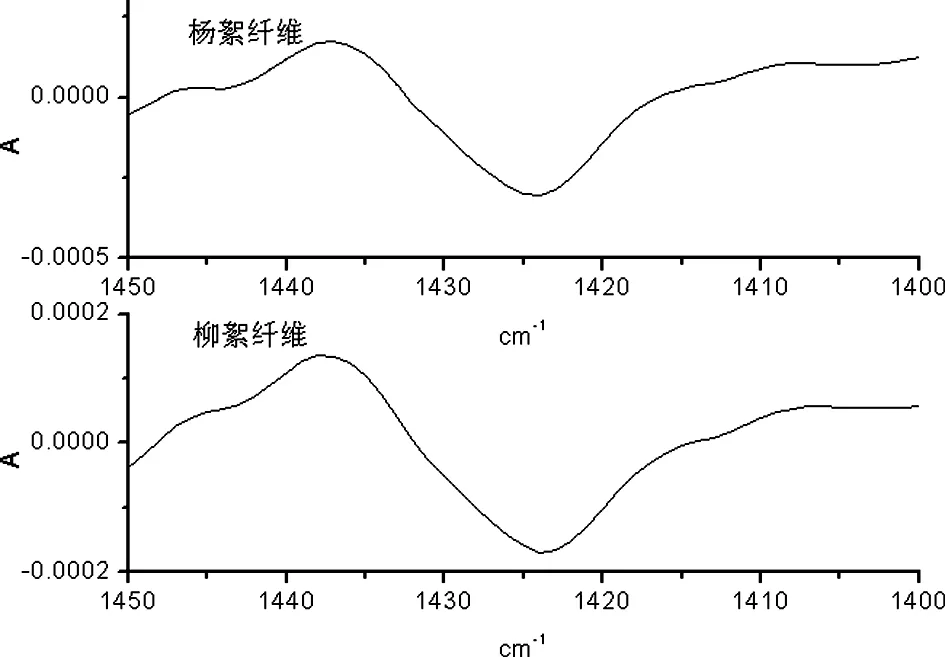
B 杨絮纤维和柳絮纤维二阶导数红外光谱
图6 杨絮纤维和柳絮纤维一维及二阶导数红外光谱(1450 cm-1~1400 cm-1)
在 1450 cm-1~1400 cm-1频率范围内首先开展了杨絮纤维及柳絮纤维的一维红外光谱的研究(图 6A)。其中 1424 cm-1频率处的红外吸收峰归属于纤维素 -OH 变形振动模式(δOH-1),但其受到纤维素 CH2变形振动模式(δCH2)和 CH变形振动模式(δCH)联合作用的影响;而进一步研究了杨絮纤维和柳絮纤维的二阶导数红外光谱(图 6B)则得到了同样的红外光谱信息,相关红外光谱数据见表 1。
2.3.2 1450 cm-1~1400 cm-1频率范围内杨絮纤维和柳絮纤维的同步二维红外光谱的研究
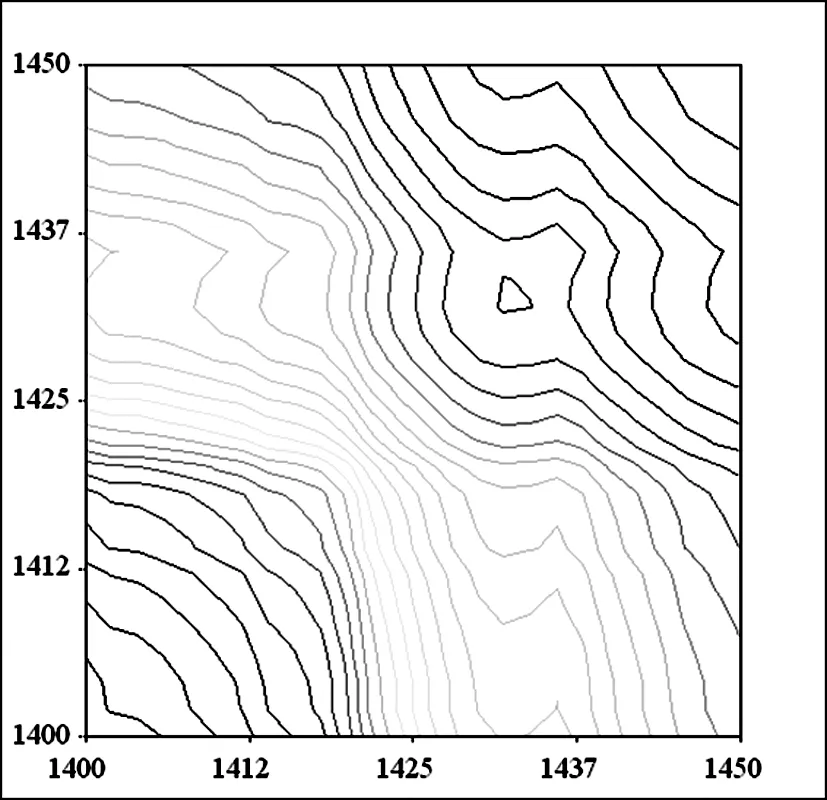
A 杨絮纤维同步二维红外光谱

B 柳絮纤维同步二维红外光谱
图7 杨絮纤维和柳絮纤维同步二维红外光谱(1450 cm-1~1400 cm-1)
在 1450 cm-1~1400 cm-1频率范围继续开展了杨絮纤维和柳絮纤维的同步二维红外光谱的研究(图 7),并没有发现明显的自动峰。
2.4 1350 cm-1~1300 cm-1频率范围内杨絮纤维和柳絮纤维的三级红外光谱的研究
2.4.1 1350 cm-1~1300 cm-1频率范围内杨絮纤维和柳絮纤维的一维及二阶导数红外光谱的研究

A 杨絮纤维和柳絮纤维一维红外光谱

B 杨絮纤维和柳絮纤维二阶导数红外光谱
在 1350 cm-1~1300 cm-1频率范围内首先开展了杨絮纤维和柳絮纤维的一维红外光谱的研究(图 8A),由其于分辨率较低,并不能得到有用的红外光谱信息。而进一步研究了杨絮纤维和柳絮纤维的二阶导数红外光谱(图 8B),其中在 1335 cm-1频率附近发现的红外吸收峰则归属于纤维素 -OH 变形振动模式(δOH-2),但同样受到纤维素δCH2和δCH联合作用的影响。
2.4.2 1350 cm-1~1300 cm-1频率范围内杨絮纤维和柳絮纤维的同步二维红外光谱的研究
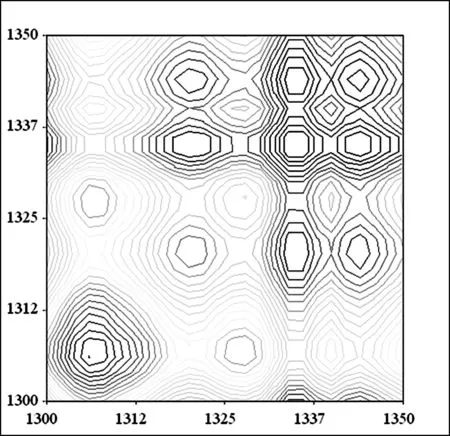
A 杨絮纤维同步二维红外光谱
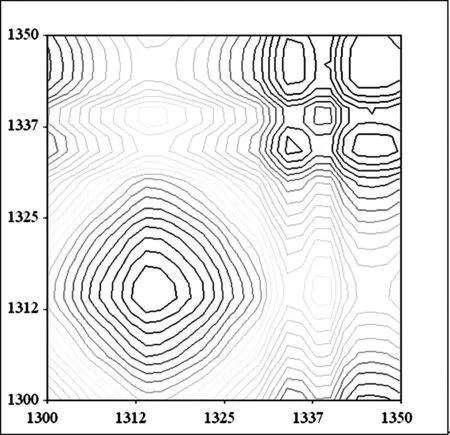
B 柳絮纤维同步二维红外光谱
图9 杨絮纤维和柳絮纤维二维相关同步红外光谱(1350 cm-1~1300 cm-1)
在 1350 cm-1~1300 cm-1频率范围继续开展了杨絮纤维的同步二维红外光谱的研究(图 9A),其中在(1335 cm-1,1335 cm-1)频率附近清晰的发现了一个相对强度较大的自动峰归属于δOH-2。而研究了柳絮纤维的同步二维红外光谱的研究(图 9B),则得到了同样的光谱信息。
2.5 1100 cm-1~1000 cm-1频率范围内杨絮纤维和柳絮纤维的三级红外光谱的研究
2.5.1 1100 cm-1~1000 cm-1频率范围内杨絮纤维和柳絮纤维的一维及二阶导数红外光谱的研究
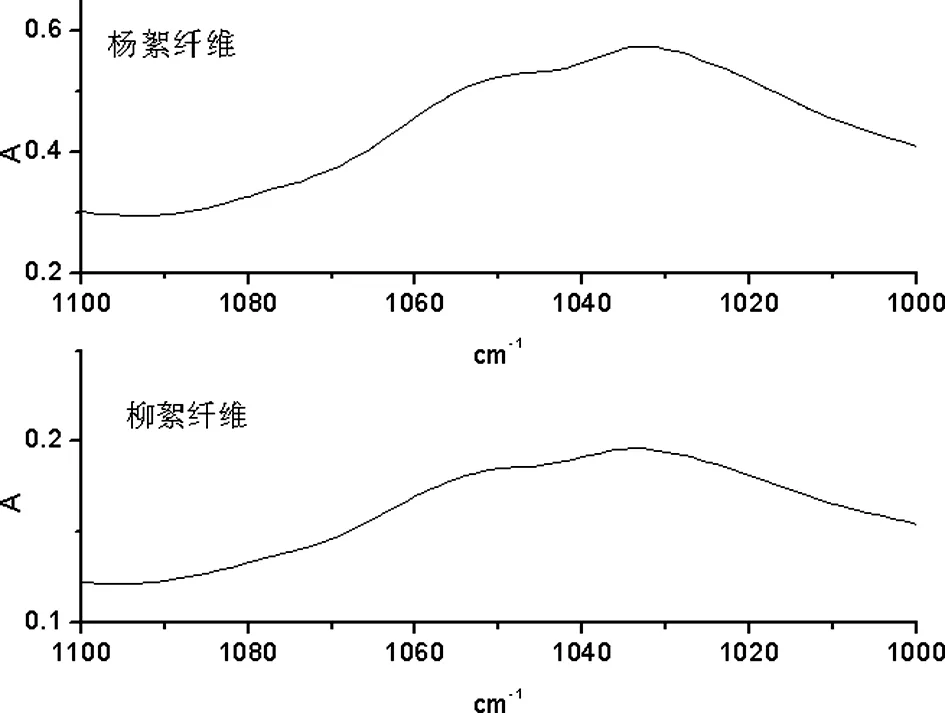
A 杨絮纤维和柳絮纤维一维红外光谱

B 杨絮纤维和柳絮纤维二阶导数红外光谱
图10 杨絮纤维和柳絮纤维一维及二阶导数红外光谱(1100 cm-1~1000 cm-1)
在 1110 cm-1~1000 cm-1频率范围内,首先开展了杨絮纤维和柳絮纤维的一维红外光谱(图 10A)的研究。由于其分辨率过低,并没有发现明显的红外吸收峰;而进一步研究了杨絮纤维和柳絮纤维的二阶导数红外光谱(图 10B),其分辨率有显著的提高,其中在 1032 cm-1和 1055 cm-1频率附近的发现的红外吸收峰归属于纤维素中 C-O 伸缩振动模式(νC-O)。
2.5.2 1100 cm-1~1000 cm-1频率范围内杨絮纤维和柳絮纤维的同步二维红外光谱的研究

A 杨絮纤维同步二维红外光谱
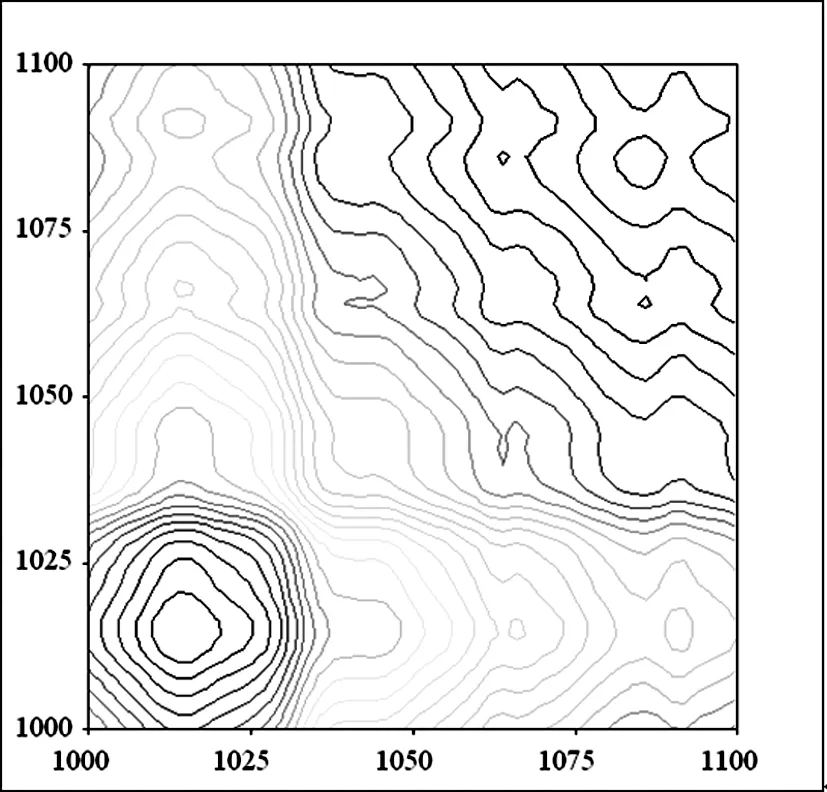
B 柳絮纤维同步二维红外光谱
图11 杨絮纤维和柳絮纤维同步二维红外光谱(1100 cm-1~1000 cm-1)
在 1100 cm-1~1000 cm-1频率范围继续开展了杨絮纤维的同步二维红外光谱的研究(图 11A)。显然,其分辨率要明显优于相应的一维红外光谱和二阶导数红外光谱。其中在(1012 cm-1,1012 cm-1)、(1043 cm-1,1043 cm-1)、(1075 cm-1,1075 cm-1)和(1085 cm-1,1085 cm-1)频率附近清晰的发现了四个相对强度较大的自动峰则归属于 νC-O。而(1012 cm-1,1012 cm-1)频率处的自动峰强度最大,说明该频率对应的官能团对于温度变化比较敏感。而在(1012 cm-1,1043 cm-1),(1012 cm-1,1075cm-1),(1012 cm-1,1085cm-1),(1043 cm-1,1075 cm-1),(1043 cm-1,1085 cm-1)和(1075 cm-1,1085 cm-1)位置处发现六个交叉峰,这说明杨絮纤维 1012 cm-1、1043 cm-1、1075 cm-1和 1085 cm-1频率对应的红外吸收峰之间存在着较强的相互作用。而研究了柳絮纤维的同步二维红外光谱(图 11B)只在(1016 cm-1,1016 cm-1)和(1085 cm-1,1085 cm-1)频率附近发现的两个相对强度较大的自动峰,其中(1016 cm-1,1016 cm-1)频率处自动峰强度较大,说明该频率处的红外吸收峰对于温度变化比较敏感。此外,在(1016 cm-1,1085 cm-1)频率处发现了一个明显的交叉峰,说明,柳絮纤维 1016 cm-1和 1085 cm-1频率对应的红外官能团之间存在着较强的相互作用。
2.6 杨絮纤维和柳絮纤维三级红外光谱的差异性研究
在 3000 cm-1~2800 cm-1、1800 cm-1~1550 cm-1、1450 cm-1~1400 cm-1、1350 cm-1~1300 cm-1和 1100 cm-1~1000 cm-1等五个频率区间内,同时开展了杨絮纤维和柳絮纤维的三级红外光谱,相关数据见表 1。

表1 杨絮纤维和柳絮纤维三级红外光谱数据
注:- 代表没有观察到明显红外吸收峰。
通过研究表 1 数据可知,在 3000 cm-1~2800 cm-1、1800 cm-1~1550 cm-1、1450 cm-1~1400 cm-1、1350 cm-1~1300 cm-1和 1100 cm-1~1000 cm-1等五个频率范围内,杨絮纤维和柳絮纤维的一维红外光谱和二阶导数红外光谱几乎完全一样,这主要是因为杨絮纤维和柳絮纤维的主要化学组成几乎完全相同,均为 α-纤维素和少量油脂及蛋白质。因此采用传统的一维红外光谱和二阶导数红外光谱并不能有效区别杨絮纤维和柳絮纤维。而杨絮纤维和柳絮纤维的同步二维红外光谱却存在着差异,这主要是因为由于杨絮纤维和柳絮纤维微观物理结构存在着差异,因此对于外加的物理扰动因素(温度变化)的敏感程度有所差异,所以采用同步二维相关红外光谱技术可以很好的区分杨絮纤维和柳絮纤维。
3 结论
在 3000 cm-1~2800 cm-1、1450 cm-1~1400 cm-1、1800 cm-1~1550 cm-1、1350 cm-1~1300 cm-1和 1100 cm-1~1000 cm-1等五个频率区间:杨絮纤维和柳絮纤维同时存在着:νasCH2、νCH、νsCH2、νC=O、酰胺峰 Ⅰ(νC=O)、酰胺峰 Ⅱ(δN-H+ νC-N)、δOH和 νC-O等红外吸收模式。研究发现,采用传统的红外光谱技术(包括:一维红外光谱和二阶导数红外光谱)无法有效的区分杨絮纤维和柳絮纤维,而采用同步二维红外光谱技术可以明显的区分杨絮纤维和柳絮纤维,并进一步研究了其光谱差异机理。
[1] 尹传青,张洪亭. 杨絮纤维形态结构与应用[J]. 山东纺织经济,2013,37(9): 37-38.
[2] Wartelle L H, Bradow J M, Hinojosa O, et al. Quantitative Populus tomentosa fiber maturity measurements by X-rayfluorescence spectroscopy and advanced fiber information system[J]. J Agric Food Chem, 1995, 43(5): 1219-1223.
[3] 蔡玉兰,王东伟. 天然竹纤维的固态核磁共振谱表征[J]. 纤维素科学与技术,2009,17(1): 1-6.
[4] Muto A, Bhaskar T, Tsuneishi S, et al. Activated carbon monoliths from phenol resin and carbonized populus tomentosa fiber for methane storage[J]. Energy Fuels, 2005, 19(1): 251-257.
[5] 王戈,黄安民,胡小霞,等. 竹原纤维和苎麻纤维鉴别方法的探讨[J]. 光谱学与光谱分析,2010,30(9): 2365-2367.
[6] 何莉萍,周海业,李新起,等. 氨基硅油改性苎麻纤维的研究[J]. 湖南大学学报(自然科学版),2012,39(9): 72-75.
[7] 刁均艳,潘志娟. 黄麻、苎麻及棕榈纤维的聚集态结构与性能[J]. 苏州大学学报(工科版),2008,28(6): 39-43.
[8] 刘伟,刘晓洪,殷肖海,等. 热处理对苎麻纤维结晶度的影响[J]. 纺织科技进展,2009(4):50-51.
[9] 陈敏. 香蕉纤维与亚麻、苎麻纤维的特征差异[J]. 宁波大学学报(理工版),2010,23(3): 109-112.
[10]李卫东,彭丽桦,王新厚. 竹原纤维和苎麻纤维鉴别方法的探讨[J].上海纺织科技,2012,40(8): 46-48.
[11]翁诗甫. 傅里叶变换红外光谱分析[M]. 北京:化学工业出版社,2010: 291-299.
[12]王正熙. 聚合物红外光谱分析和鉴定[M]. 成都:四川大学出版社,1989: 221-223.
[13]Maekawa H, Ballano G, Tonioilo C, et al. Linear and two-dimensional infrared spectroscopic study of the amide Ⅰ and amide Ⅱ modes in fully extended peptide chains[J]. J Phys Chem B, 2011, 115 (18): 5168-5182.
[14]Noda I. Two-dimensional infrared (2D IR) spectroscopy: theory and applications[J]. Applied Spectroscopy, 1990, 44(4): 550-551.
[15]Maekwa H, Ge N. Comparative study of electrostatic modes for amide Ⅰ and amide Ⅱ modes: linear and two-dimensional infrared spectra[J]. J Phys Chem B, 2010, 114 (3): 1434-1446.
[16]Wang J P. Ab inito-bases all-mode two-dimensional infrared spectroscopy of sugar molecule[J]. J Phys Chem B, 2007, 111(31): 9193-9196.
[17]王维,隋丽丽,李中秋,等. 氮化硅 νSi-N伸缩振动的衰减全反射红外光谱研究[J]. 理化检验(化学分册),2016,52(11): 1241-1245.
2017-03-21
河北省科技厅科学技术研究与发展计划(12222802),石家庄市科学技术研究与发展计划课题(151500182A)。
于宏伟(1979-),男,博士,副教授,研究方向:纺织材料的结构研究。
TS102,O434.3
A
1008-5580(2017)03-0017-08
猜你喜欢
作文与考试·小学低年级版(2021年15期)2021-09-13
音乐天地(音乐创作版)(2021年4期)2021-07-12
应用数学(2020年2期)2020-06-24
数学物理学报(2018年6期)2019-01-28
中央民族大学学报(自然科学版)(2018年3期)2018-11-09
儿童故事画报·自然探秘(2017年7期)2018-03-14
诗林(2016年5期)2016-10-25
火花(2015年7期)2015-02-27
红蜻蜓·低年级(2014年5期)2014-07-29
