
干旱胁迫下沙芥幼苗叶片光合特性和叶绿素荧光参数的变化
2017-07-19 10:03张希吏
干旱地区农业研究 2017年3期
王 萍,张希吏,石 磊
(内蒙古农业大学农学院/内蒙古自治区野生特有蔬菜种质资源与种质创新重点实验室, 内蒙古 呼和浩特 010019)
干旱胁迫下沙芥幼苗叶片光合特性和叶绿素荧光参数的变化
王 萍,张希吏,石 磊
(内蒙古农业大学农学院/内蒙古自治区野生特有蔬菜种质资源与种质创新重点实验室, 内蒙古 呼和浩特 010019)
以一年生沙芥幼苗的叶片为实验材料,采用盆栽实验的方法,将土壤含水量设置为正常浇水为对照(70%~75%WHC,CK),轻度干旱胁迫(55%~60%WHC,LD),中度(40%~45%WHC,MD),重度(25%~30%WHC,VD)四个处理,用Li-6400 光合仪和叶绿素荧光仪FMS2(Hansatech, UK),研究干旱胁迫下沙芥光合参数、荧光参数的变化规律,探讨干旱胁迫对沙芥叶片光合生理指标的影响,旨在揭示沙芥对干旱胁迫的响应机制,为阐明沙芥的耐旱性提供理论依据。结果表明:(1) 随着干旱胁迫程度的加剧,沙芥叶片叶绿素a,b含量在各干旱胁迫处理下,比对照下降了1.30%~7.04%和19.47%~42.37%,类胡萝卜素含量下降了2.06%~9.88%。叶绿素a/b上升了33.49%~74.27%,Car/Chl上升了1.33%~15.73%。(2) 随着干旱胁迫的加剧,沙芥叶片的净光合速率、蒸腾速率、水分利用率呈现下降的趋势,胞间CO2浓度先下降后上升,气孔限制值不断下降。这表明轻度干旱胁迫下气孔限制是影响沙芥幼苗叶片Pn下降的主要因素;中度和重度干旱胁迫下,非气孔限制是沙芥幼苗叶片Pn下降的主要因素。(3) 干旱胁迫下,沙芥叶绿素荧光参数Fv/Fm、Fv/Fo、ΦPSⅡ,qP和ETR均呈下降趋势,qN在干旱胁迫下不断上升。
沙芥幼苗;干旱胁迫;光合参数;叶绿素荧光参数
光合作用是植物生长的基础,是植物生产力构成的最主要的因素,水分状况又是影响光合作用最重要的因子之一,干旱会导致叶片气孔关闭,严重时甚至损伤叶肉细胞、降低光合酶的活性、破坏叶绿体的结构[1],降低叶绿素和类胡萝卜素的含量,使植物光合速率降低[2]。植物叶绿素荧光分析技术是近年发展起来的用于光合作用机理研究和光合生理状况检测的一种新技术。与一些表现性的气体交换指标相比,叶绿素荧光参数更具有反映内在性的特点,因而被视为研究植物光合作用与环境关系的内在探针[3]。沙芥〔Pugioniumcornutum(L.) Gaertn〕为十字花科沙芥属一、二年生草本植物,别名山羊沙芥、山萝卜、沙盖、蒙名额乐孙萝邦等,为中国的特有种[4]。沙芥生长于干旱、半干旱地区的固定和半固定沙丘上,是一种典型的沙生植物[5]。它主要分布在毛乌素沙漠及其边缘地区的流动与半流动的沙丘上,在甘肃、宁夏、陕西、内蒙古等地区均有分布,它的主根发达,是防风固沙的先锋植物。目前,随着人们对野生植物沙芥生态作用和营养价值认知程度的增加,在分类、种子萌发[6]、传粉习性[7]、营养成分等方面已有一些报道[8]。迄今为止,关于土壤干旱胁迫下沙芥光合作用的研究较少,对沙芥在干旱胁迫下叶绿素荧光动力学特征的研究尚未见报道。为此,本研究通过对沙芥幼苗进行盆栽水分处理试验,以了解沙芥幼苗在干旱胁迫下光合和荧光指标的变化,揭示沙芥光合作用对土壤干旱胁迫的响应机制,为深入了解沙芥的光合生理特性提供参考。
1 实验材料
本实验材料来自于鄂尔多斯市毛乌素沙地自然环境。试验地位于内蒙古农业大学的日光温室中。于2013年4月下旬将去果皮的沙芥种子进行消毒、浸种,之后均匀摆入底部覆有滤纸的培养皿中进行种子萌发,待胚根长出后播种在PVC花盆,并置于大棚中,花盆的规格为高18 cm,内径15 cm,基质为沙∶土壤=4∶1,土壤中全氮含量为76.21 mg·100g-1,有效磷含量为28.50 mg·kg-1,速效钾含量158.0 mg·kg-1,有机质含量为1.95%,pH值为7.81。田间最大持水量12.4 g·cm-3,每盆装材料1.5 kg。每盆播种4~5粒,出苗后间苗,每盆保证幼苗2株。在幼苗长到六叶一心时进行土壤干旱胁迫处理。干旱胁迫程度按叶片萎焉程度和Hsiao[9]标准划分:对照、轻度胁迫、中度胁迫、重度胁迫处理土壤含水量分别占田间最大持水量的70%~75%、55%~60%、40%~45%、25%~30%。采用称重法,于每日早晨6∶00和傍晚18∶00对花盆进行称重,补充水分至相应的重量范围,因为花盆质量很轻为90 g,它对幼苗的影响可以省略不计,每个处理重复4次。干旱处理12 d后,于第二天清晨9∶00~11∶00进行各个梯度沙芥幼苗叶片光合特性和荧光参数的测定。
2 测定方法
2.1 土壤相对含水量的测定
土壤相对含水量(%)=土壤含水量/田间最大持水量×100%
土壤含水量(%)=(w1-w2)/(w2-w0)×100%
式中,w0=铝盒重,w1=铝盒重+风干土样重,w2=铝盒重+烘干土样重
2.2 沙芥叶片光合参数和叶绿素荧光参数的测定
沙芥叶片的光合特性利用采用Li-6400便携式光合仪在干旱处理后第二天早上9∶00~11∶00进行测定,所用光源为Li-6400配置的红蓝光LED光源,杨晓宇[10]对沙芥叶片的光饱和点和光补偿点进行研究,得出沙芥幼苗叶片的光饱和点为1 325 μmol·m-2·s-1,故此,本实验光合有效辐射设置为1 200 μmol·m-2·s-1,控制样本室内气流速率为500 μmol·s-1,参比室CO2浓度为400 μmol·mol-1,控制温度为起始时的外界环境温度,测定时,选取每种处理长势相同的植株,从外展开的第3,4片叶进行挂牌标记作为测定对象,每个处理测定8片叶。测定沙芥叶片净光合速率(Pn)、蒸腾速率(Tr)等相关指标,并计算水分利用率(WUE=Pn/Tr),整个植株的WUE=干物质量/水分损失总量×100%,气孔限制值Ls(%)=(1-Ci/Ca)×100,其中Ci为胞间CO2浓度,Ca为空气中CO2浓度。叶绿荧光参数使用FMS2(Hansatech, UK)进行测定;测定前叶片暗处理30 min。先用600 Hz测定光测定初始荧光(Fo),随后给叶片施加1 500 μmol·m-2·s-1饱和脉冲光0.7 s,测得最大荧光(Fm)。然后,打开作用光(600 μmol·m-2·s-1)。在荧光下降至施加饱和光前稳定水平之后再20 s,测得最小荧光(F0)。在仪器上直接读出参数值,如暗适应叶片可变荧光(Fv),PSⅡ最大光化学效率(Fv/Fm),实际光化学效率(ΦPSⅡ),光化学荧光猝灭系数qP,非光化学荧光猝灭系数qN,电子传递速率(ETR)。
2.3 数据分析
数据分析使用Excel和SPSS19.0统计软件进行数据处理和单因素方差分析(Duncan新复极差法,P=0.05和P=0.01)。
3 结果与分析
3.1 干旱胁迫下沙芥幼苗叶片光合色素含量和光合参数的变化
从表1可以看出,随着干旱胁迫程度的加剧,沙芥叶片叶绿素a,叶绿素b,类胡萝卜素含量逐渐下降,重度胁迫与对照相比,差异极显著(P<0.01)。叶绿素a/b的值不断上升,轻度胁迫时,与对照相比,差异不显著(P>0.05)。与对照相比,中度与重度时,差异极显著(P<0.01)。Car/chl比值先下降后上升,与对照相比,中度与重度时,差异极显著(P<0.01)。
从表2中可以看出,随着干旱胁迫的加剧,沙芥叶片的净光合速率呈先上升后下降趋势,轻度胁迫与对照相比有所上升,差异显著(P<0.05),中度和重度胁迫下,与对照相比有所下降,差异极显著(P<0.01)。从表2可以看出,沙芥叶片蒸腾速率呈下降趋势,与对照相比,轻度胁迫时差异不显著(P>0.05),中度和重度胁迫时差异极显著(P<0.01)。从表2中可以看出,随着干旱胁迫的加剧,与对照相比,水分利用率在轻度胁迫时差异不显著(P>0.05),中度和重度胁迫时下降明显,均表现出显著差异(P<0.01)。
从表2可以看出,叶片胞间CO2浓度在干旱胁迫下,呈现先下降后上升的趋势,与对照达到差异极显著(P<0.01)。与对照相比,在轻度胁迫下气孔导度值差异不显著(P>0.05),说明在轻度干旱的条件下,沙芥幼苗叶片气孔能够保持一定水平的开度,利于蒸腾和光合。然而,在中度和重度胁迫下,逐渐下降,与对照相比,差异极显著(P<0.01)。气孔限制值随着干旱胁迫的加剧呈现先上升后下降的趋势,轻度胁迫时高于对照,差异不显著(P>0.05),中度和重度干旱时低于对照,差异极显著(P<0.01)。
3.2 干旱胁迫下沙芥幼苗叶片叶绿素荧光参数的变化
Fv/Fm反映的是PSⅡ反应中心内的光能转化效率,非环境胁迫条件下叶片的荧光参数Fv/Fm极少变化,不受物种和生长条件的影响,而遭受在逆境或受伤害时这一参数变化明显;Fv/Fo则反映了PSⅡ的潜在活性,它们是表明光化学反应状况的两个重要参数[11]。从图1中可知,随着干旱胁迫程度的加剧,Fv/Fm与Fv/Fo呈下降趋势,与对照相比,Fv/Fm在轻度胁迫时,差异不显著(P>0.05);中度与重度胁迫下,差异极显著(P<0.01)。与对照相比,轻度、中度和重度胁迫下,Fv/Fm的降幅分别为5.71%,15.54%和24.70%;与对照相比,Fv/Fo在轻度胁迫时,差异显著(P<0.05);中度与重度胁迫下,差异极显著(P<0.01);与对照相比,轻度、中度和重度胁迫下,Fv/Fo的降幅分别为9.16%,19.93%和29.22%。以上表明,干旱胁迫使 PSⅡ受到了伤害,降低了 PSⅡ原初光能转化效率,使沙芥幼苗叶片PSⅡ潜在活性中心受损,光合作用原初反应过程受抑制。

表1 干旱胁迫对沙芥叶片光合色素含量的影响
注:同列数据后大、小写字母分别表示不同处理与CK间在0.01和0.05水平上有显著差异,下同。
Note: Capital and lowercase letters in the same column indicated significant difference between drought treatment and controls at 0.01 and 0.05 level, respectively. The same as below.

表2 干旱胁迫对沙芥幼苗叶片光合特性的影响
ΦPSⅡ反映叶片在光适应下PSⅡ反应中心的实际光化学效率,即光下植物叶片捕获光能中用于电子传递的比例,ΦPSⅡ的大小意味PSⅡ的实际光化学反应所利用的光能和过剩光能的多少[12-13]。从图1中可知,随着干旱胁迫程度的加剧,ΦPSⅡ逐渐下降。与对照相比,轻度胁迫时,ΦPSⅡ缓慢下降,与对照之间差异不显著(P>0.05),中度和重度胁迫时差异极显著(P<0.01)。与对照相比,轻度、中度和重度胁迫下,降幅分别为7.74%,18.42%和26.70%。以上表明,随着干旱胁迫的加剧,用于光化学反应的光能显著降低,过剩光能增多。
光化学猝灭系数qP表示PSⅡ反应中心吸收的光能用于光化学电子传递的份额,所以光化学在一定程度上反映了PSⅡ反应中心开放程度和活性状态[14]。从图1中可知,qP随着干旱胁迫的加剧不断下降,与对照相比,轻度胁迫时差异不显著(P>0.05),中度和重度胁迫,差异极显著(P<0.01)。说明干旱胁迫降低了PSⅡ反应中心开放程度,减弱了受体侧接受电子的能力。随着干旱胁迫的加剧,与对照相比,轻度、中度和重度胁迫下qP的降幅分别为6.12%,17.73%和22.80%。
qN表示叶片捕获的光能中用于非辐射能量耗散的量[15],随着干旱胁迫的加剧,qN逐渐上升,以中度胁迫下qN上升幅度最大,其次为轻度,重度胁迫上升幅度最小,与对照相比,轻度与中度达到极显著差异(P<0.01)。可见随着干旱胁迫的加剧,叶片吸收光能大部分以热量的形式耗散,以保护光合机构免受过剩光能破坏。
ETR表示表观电子传递速率(通过光系统的非环式电子传递),从图1中可知,轻度胁迫下沙芥幼苗叶片的ETR含量缓慢下降,与对照相比,差异不显著(P>0.05),与对照相比,中度和重度胁迫下,差异极显著(P<0.01)。表明轻度胁迫时,表观电子传递速率受影响较小,随着胁迫程度的加剧,电子传递速率下降。与对照相比,轻度胁迫、中度胁迫和重度胁迫下,ETR降幅分别为5.46%、17.83%和20.45%。
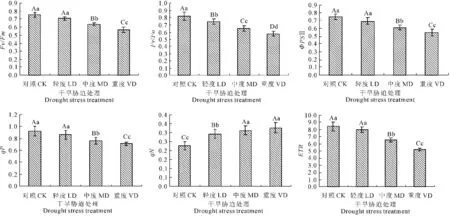
图1 干旱胁迫下沙芥幼苗叶绿素荧光参数的变化
4 讨 论
4.1 干旱胁迫下沙芥叶片光合色素和光合特性的变化
叶绿素是光合作用的光敏催化剂,与光合作用密切相关,其含量和比例是植物适应和利用环境因子的重要指标,叶绿素含量和类胡萝卜素含量的变化反映了叶片对外界光环境变化的适应情况[16]。在本研究中,叶绿素a,b的含量都在下降,但叶绿素b下降的幅度要大于叶绿素a,所以Chla/b的值升高,可减少沙芥叶片对光能的捕捉,降低光合机构受光氧化的损伤,是沙芥叶片适应干旱胁迫的光保护调节措施。随着干旱胁迫加重,沙芥幼苗的光合色素会受到损伤,导致沙芥幼苗的Chl和Car含量下降,而Car的稳定性高于Chl,所以其下降幅度小于Chl,造成沙芥幼苗中Car在光合色素中占有的比例较高,因此Car/Chl的比值会上升。这就说明给予适当的干旱胁迫,使沙芥幼苗的Car含量相对升高,就有利于保护沙芥叶片的光合机构,防止Chl的光氧化破坏。
影响植物光合作用的因素有气孔限制和非气孔限制之说。Farquhar和Sharkey认为,只有当光合速率和胞间CO2浓度变化方向相同,且气孔限制值增大,才可认为光合速率的下降主要由气孔因素引起;如果胞间CO2浓度和净光合速率变化方向相反,气孔限制值减小,则净光合速率下降归因于叶肉细胞同化能力的降低[17]。本实验结果表明在轻度胁迫时,沙芥和斧翅沙芥幼苗叶片光合速率与胞间CO2变化方向相反,而气孔限制值增大,说明在轻度胁迫下,气孔因素不会引起沙芥幼苗叶片光合速率的下降;而在中度和重度胁迫处理下,沙芥幼苗叶片光合速率明显下降,而胞间CO2浓度却上升,气孔限制值减小,说明在中度和重度干旱胁迫下,沙芥幼苗叶片光合速率下降主要原因由叶肉细胞同化能力降低等非气孔因素引起的。这与杨晓宇研究得出的结论一致[10]。随着干旱胁迫的加剧,沙芥单株幼苗的水分利用率不断下降,因为光合速率的降低会影响沙芥幼苗整个植株的生长和产量的形成,干旱胁迫越严重,沙芥植株的水分利用率越低,产量降低越明显。
4.2 干旱胁迫下沙芥叶片叶绿素荧光参数的变化
正常情况下,叶绿素吸收的光能主要通过光合电子传递、叶绿素荧光和热耗散三种途径来消耗,这三种途径存在着此消彼长的关系,光合作用和热耗散的变化能引起荧光发射的响应变化,因此,可以通过对荧光的观测来探究光合电子的传递和热耗散[18]。本试验研究结果表明,在轻度胁迫时Fv/Fm、Fv/Fo、ΦPSⅡ,qP、ETR下降幅度较小,qN上升幅度较大,说明叶片的光系统Ⅱ受到的损伤较轻,积累在PSⅡ反应中心的过剩光能可以通过热形式耗散,随着干旱胁迫的加剧,Fv/Fm、Fv/Fo、ΦPSⅡ,qP、ETR下降幅度逐渐增大,在重度胁迫下,qN上升幅度下降,说明干旱胁迫到一定程度,导致沙芥叶PSⅡ反应中心的开放程度降低,光合电子传递速率下降,使PSⅡ潜在活性受损,直接影响了光合作用的电子传递,使积累在PSⅡ反应中心的光能过剩,相关保护机制无法消耗过剩的光能,使光合机构受到过剩光能的损害,产生光抑制。
[1] 韩瑞宏,卢欣石,高桂娟.紫花苜蓿对干旱胁迫的光合生理响应[J].生态学报,2007,27(12):5229-5237.
[2] 蒙祖庆,宋丰萍,刘振兴.干旱及复水对油菜苗期光合及叶绿素荧光特性的影响[J].中国油料作物学报,2012,34(1):40-47.
[3] Krause G H, Weis E. Chlorophyll fluorescence and photosynthesis: the basics[J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1991,42:313-349.
[4] 赵一之.沙芥属的分类校正及其区系分类[J].内蒙古大学学报:自然科学版,30(2):197-199.
[5] 马毓泉.内蒙古植物志第二卷(第二版)[M].呼和浩特:内蒙古人民出版社,1990:612-616.
[6] 张卫华.沙芥种子发芽生理的研究[D].呼和浩特:内蒙古农业大学,2004.
[7] 黄修梅,郝丽珍,胡宁宝,等.中国特有种——沙芥繁育系统和传粉生物学研究[J].西北植物学报,2009,29(11):2232-2237.
[8] 贺勇鹏,杨忠仁,宋兆伟,等.沙芥属蔬菜不同苗龄植株叶片营养成分含量变化规律的研究[J].内蒙古农业大学学报,2009,30(3):87-89.
[9] Hisao T C. Plant responses to water stress[J]. Plant Physiology,1973,24:519-570.
[10] 杨晓宇.沙芥属蔬菜幼苗光合特性及其对干旱胁迫响应的研究[D].呼和浩特:内蒙古农业大学,2009.
[11] 温国胜,田海涛,张明如,等.叶绿素荧光分析技术在林木培育中的应用[J].应用生态学报,2006,17(10):1973-1977.
[12] 孙永江,付艳东,杜远鹏,等.不同温度/光照组合对‘赤霞珠’葡萄叶片光系统Ⅱ功能的影响[J].中国农业科学,2013,(6):1191-1200.
[13] 苏秀荣,王秀峰,杨凤娟.硝酸根胁迫对黄瓜幼苗叶片光合速率、PSⅡ光化学效率及光能分配的影响[J].应用生态学报,2007,18(7):1441-1446.
[14] 杨晓青,张岁岐,梁宗锁,等.水分胁迫对不同抗旱类型冬小麦幼苗叶绿素荧光参数的影响[J].西北植物学报,2004,24(5):812-816.
[15] 邢庆振,郁松林,牛雅萍,等.盐胁迫对葡萄幼苗光合及叶绿素荧光特性的影响[J].干旱地区农业研究,2011,29(3):96-100.
[16] 王满莲,韦 霄,唐 辉,等.光强对三种喀斯特植物幼苗生长和光合特性的影响[J].生态学杂志,2015,34(3):604-610.
[17] Farquhar G D, Sharkey T D. Stomatal conductance and pho-tosynthesis[J]. Ann Rev Plant Physiology, 1982,33:317-345.
[18] 张秋英,李发东,刘孟雨,等.不同水分条件下小麦旗叶叶绿素a荧光参数与子粒灌浆速率[J].华北农学报,2003,18(1):26-28.
ChangesofphotosyntheticpharacteristicsandchlorophyllfluorescenceparametersofPugioniumcornutum(L.)Gaertnseedlingsleafunderdroughtstress
WANG Ping, ZHANG Xi-li, SHI Lei
(CollegeofAgronomy,InnerMongoliaAgriculturalUniversity/InnerMongoliaAutonomousRegionKeyLaboratoryof
WildPeculiarVegetableGermplasmResourceandGermplasmEnhancement,Huhhot010019,China)
With the help of the method of a pot-cultivation experiment and the reference of different soil content with 70%~75%WHC(CK), 55%~60%WHC(LD), 40%~45%WHC(MD), 25%~30%WHC(VD) and with FMS-2 and Li-6400 photosynthetic apparatus, choosing seedling leaves ofPugioniumcornutum(L.) Gaertn as the experimental materials, this paper studies the change laws of hlorophyll fluorescence parameters ofPugioniumcornutum(L.) Gaertn under the drought stress, and the effects of drought stress on its photosynthetic index. The purpose of this paper is to reveal the mechanism ofPugioniumcornutum(L.) Gaertn dealing with the drought stress, and to illustrate its drought resistance. The results showed that: (1) With intensifying drought stress, by contrast, the content ofPugioniumcornutumleaf chlorophyll a, chlorophyll b inP.cornutum(L.) Gaertn decreased by 1.30%~7.04% and 19.47%~42.37% respectively, carotene content by 2.06%~9.88%; the value of chlorophyll a/b and Car/Chl increased by 33.49%~74.27% and 1.33%~15.73% respectively. (2) With intensifying the drought stress, there were distinct decrease trends of the net photosynthetic rate(Pn), transpiration rate(Tr), stomatal conductance(Gs) and water use efficiency(WUE) ofP.cornutum(L.) Gaertn leaves, and the carbon dioxide concentration decrease at first and then increase and the stomatal limitation(Ls) decrease, which showed thatPndecrease ofP.cornutum(L.) Gaertn leaves under light stress was mainly induced by stomatal factors, andPndecrease under moderate and severe stress was mainly induced by non-stomatal factors. (3) Under drought stress, there were decrease trends ofFv/Fm,Fv/Fo,ΦPSⅡ,qP,ETRofPugioniumcornutum(L.) Gaertn leaf, and increase trends ofqN.
Pugioniumcornutu(L.) Gaertn seedling; drought stress; photosynthetic parameters; chlorophyll fluorescence parameters
1000-7601(2017)03-0159-05doi:10.7606/j.issn.1000-7601.2017.03.25
2016-04-14
:2017-02-24
:国家自然科学基金项目(31260475,30960236,30560088);内蒙古农业大学创新(培育)团队(NDPYTD2013-3)
王 萍,副教授,硕士生导师,主要从事种质资源与种质创新研究。 E-mail: wangping@imau.edu.cn。
S637
: A
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
阅读(科学探秘)(2020年8期)2020-11-06
电脑报(2020年12期)2020-06-30
装备制造技术(2019年12期)2019-12-25
绿色科技(2019年2期)2019-05-21
科技创新与品牌(2019年12期)2019-02-06
中国果业信息(2019年1期)2019-01-05
中成药(2018年1期)2018-02-02
农民致富之友(2017年15期)2017-08-20
中国铸造装备与技术(2015年5期)2015-12-10
