
2,4-滴的回顾与展望(上)
2017-07-10 13:47编译
世界农药 2017年3期
筱 禾 编译
2,4-滴的回顾与展望(上)
筱 禾 编译
(上海市农药研究所,上海 200032)
2,4-滴用于农业生产已有70多年历史,至今在世界各地的杂草防除中起着重要作用。有说法认为,2,4-滴是19世纪40年代早期由英美科学家经系列多次、独立试验发现的,但由于历史原因,早期与氯苯氧乙酸类除草剂相关的著作和专利并未及时记载。尽管对2,4-滴的发现过程还存有争议,但1945年该剂的商业化彻底改变了杂草防除,促进了杂草科学乃至整个农化行业的发展。同一时期开发了其他苯氧乙酸类(2甲4氯、2,4,5-涕、2,4-滴丁酸、2,4-滴丙酸等),2,4-滴是氯苯氧类化合物中应用最广泛的。虽然2,4-滴已被广泛地研究,但相关新研究还在继续。
1 类型与化学性质
2,4-滴纯品为具有相对非挥发性的干结晶固体。该剂仅微溶于水(44 558 mg/L),因此必须将其改性制成易于分散并与水形成合适混合物的制剂。表1比较了各种类型2,4-滴的相对性质。胺盐和酯类是已在市场上得到广泛接受的2种基本2,4-滴类型,最近引入了第3种——2,4-滴胆碱。

表1 2,4-滴的主要类型及其化学性质
1.1 胺盐
当2,4-滴酸与胺反应时,生成了2,4-滴盐。盐基剂型使2,4-滴酸活性成分具有水溶性。2,4-滴胺盐制剂包括异丙胺、三异丙醇胺、二乙醇胺和二甲胺,其中二甲胺的应用最为广泛。胺剂型易溶于水(以重量计,超过50%)并形成真正的溶液。由于胺盐更易在水中溶解,故其逐渐取代了各种矿物盐(锂、钾、钠和铵)。
当这些盐制剂被置于水中时,会解离或分离成带有负电荷的酸性部分和携带正电荷的胺部分。在硬水中,高含量Ca2+、Mg2+和其他阳离子可与带负电荷的2,4-滴酸部分结合,形成可沉淀的不溶性盐,减少了溶液中除草剂的量,堵塞了喷雾器械筛网及喷头。因此,在大多数胺制剂中加入螯合剂来减少或消除这一问题。大多数2,4-滴胺盐溶于水时形成澄清溶液,但在制剂中加入螯合剂使溶液呈较暗的琥珀色。这种颜色变化不会影响制剂的生物活性或作物的耐药性。大多数胺盐不溶于矿物油。盐可以制成液体或固体制剂。
1.2 胆碱盐
最近,陶农科公司开发了2,4-滴胆碱盐。2,4-滴的这种盐制剂比其他类型的2,4-滴具有更好的稳定性和更低的挥发性。最近在美国登记了2,4-滴胆碱+草甘膦混剂。2,4-滴胆碱的其他制剂正在开发中。
1.3 酯类
2,4-滴酸与醇反应生成酯。2,4-滴酯类有丁氧基乙酯(BEE)、2-乙基己基酯(2-EHE,以前称为异辛酯)、丙二醇丁醚酯、甲酯、异丙酯和丁酯。BEE和2-EHE已逐渐替代了上述所提及的其他酯类,是除了专用于柑橘的异丙酯之外,仅有的支持再登记的专用酯类。由具有4个或更少碳原子烷基链的醇生成的酯被认为挥发性较高,如甲酯、异丙酯和丁酯。从20世纪80年代初开始,这些最常用剂型的退市显著地减少了由蒸汽漂移引起的对敏感植物的药害。另一方面,由具有4个以上碳原子的烷基链的醇生成的2,4-滴酯类被归类为低挥发性酯。也就是说,碳链越长,挥发性越低。由长链醇生成的低挥发性2,4-滴酯类有BEE和2-EHE(具有8个碳链)。酯易溶于矿物油,但不溶于水。因此,它们被制成可用于水或油中的乳油。当将2,4-滴酯类乳油剂型与水混合时,乳化剂使微小的油状液滴悬浮并分散,将酯保持在水包油型乳液中。2,4-滴乳液呈乳白色。一旦静置,油滴可能分离,但搅拌会改变乳液。
总之,在干燥等环境中2,4-滴酯类比盐类制剂具有更高的活性,但也可能因作物的快速吸收短暂压制植物的解毒机制,而不能及时解毒,从而对作物产生更大的药害。酯类制剂被认为比盐类制剂更易被植物吸收。一旦施用,酯类制剂会快速渗入叶表面(角质层)并转化为酸。当加入到水中时,酯类制剂不会像胺类制剂那样离解或离子化,因此不会与硬水中的阳离子反应形成不溶性沉淀物。
1.4 其他剂型
多年来,已生产了多种不同剂型的2,4-滴。固体和液体的酸类制剂以及锂、钠、三酚胺等盐类制剂已经商业化,但目前使用受到限制甚至退市。
2 吸收与输导
2,4-滴的吸收与输导是许多韧皮部输导型除草剂的代表,在20世纪50年代至70年代进行了广泛的研究。许多环境、植物和应用因素都会影响2,4-滴吸收,但输导较少出现变化。下文讨论影响吸收的因素,包括对输导的任何可能影响。
2.1 环境因素
2.1.1 光
光强和光质通过控制光合作用、光合同化物运输和角质层特性来影响除草剂吸收和输导。已就光对2,4-滴吸收与输导的影响进行了少量研究。与暗中相比,有光时2,4-滴的吸收持续增加,但光强的影响似乎因物种而异。1980年有研究者发现在印第安麻(L.)中光强可能会影响输导,当光强从50通量增加至75通量时,处理叶表面的量减少14%以上。
2.1.2 温度
温度通过影响光合速率、改变角质层及许多其他植物相关过程影响除草剂吸收和转运。在一定范围内,温度越高,则光合作用越强,光合同化物产生越多,酶活性越高,韧皮部装载越多,进而可能使除草剂输导增加。植物角质层和气孔开口的物理状态影响除草剂吸收,此可以通过提高温度来改变。一般来说,温度越高,2,4-滴吸收越多,但输导不受影响。
2.1.3 湿度
在除草剂应用时,相对湿度通过影响气孔导度、除草剂液滴干燥时间等因素来影响除草剂吸收,但其对除草剂输导的影响不常见。白杨树在高湿度条件下吸收倍增,在低湿度条件下干燥后再喷湿时,2,4-滴吸收增加。湿度为70%~74%时的吸收高于湿度为34%~48%时,而在2个湿度水平下除草剂喷雾液滴的干燥时间差异不大。苔草(Schwein)在100%湿度水平时对2,4-滴吸收高于湿度为40%时,且在100%相对湿度条件时,分生组织和根茎的输导速度更快。当湿度从40%增加到90%时,葡萄(cv.)对2,4-滴的吸收增加了11%,而输导不受影响。
2.1.4 水分胁迫
由于干旱胁迫使吸收和输导下降,会影响某些除草剂的除草活性。水分或干旱胁迫下的植物可能因气孔开度减小、角质层较厚或变化使除草剂吸收水平下降。有效水分和气体交换的降低导致光合同化物产生和运输减少,进而输导可能会下降。短期水分胁迫通常对2,4-滴吸收没有影响,但会引起输导水平降低。菜豆(L.)在膨压水平为66%~88%时对2,4-滴酸的吸收相当,而当土壤水分为13%时,处理后叶片的输导减少。类似地,在不同土壤水分含量时,2,4-滴三甲胺盐的吸收没有差异,但在0.3 atm土壤张力时的转运比4 atm时高2倍。与通过土壤产生水分胁迫相反,大豆[max (L.) Merr.]中的水分胁迫是由于使用聚乙二醇(PEG)溶液所致。当使用PEG时,植物相对含水量从88%降至75%,叶片水势从-0.55 MPa降至-1.21 MPa,在模拟水分胁迫的条件下,使大豆叶中2,4-滴叶通量减少了40%。但应用PEG引起的短期水分胁迫以及缺乏预处理可能会改变叶片解剖结构,与其他水分胁迫研究相对照,或可解释这些矛盾的结果。
2.2 植物因素
2.2.1 生长阶段及环境
植物的年龄、叶龄和生长环境均对除草剂吸收和转运具有影响。叶龄通过角质层的生长和成熟影响除草剂吸收,植物生长的环境或生长条件影响除草剂的吸收和输导。由于除草剂的输导以从源到汇的方式出现,正在发育的幼叶中与成熟老叶的输导可能有所不同。叶龄和环境对2,4-滴吸收的影响因物种而异。菜豆、豌豆(L.)、甜菜(s L.)和向日葵(L.)幼叶对2,4-滴的吸收比成熟老叶多,但玉米或棉花的不同叶龄叶片没有吸收差异。幼叶叶片具有较薄的、与成熟叶片化学成分不同的蜡质层,可影响除草剂吸收。与6周龄和16周龄成熟植物相比,5周龄田旋花(L.)幼苗对除草剂的吸收较少,但输导更多。因角质层差异,田间种植葡萄比温室种植葡萄对除草剂的吸收少26%。
2.2.2 叶片和细胞因素
植物叶片的一些特征和特性会影响除草剂的吸收和输导。叶倾角、角质层、绒毛和其他因素影响植物对除草剂吸收量,叶部和茎部植物细胞的结构可改变除草剂输导模式。已进行了许多测定各种植物角质层厚度和化学成分对2,4-滴吸收影响的研究。发现2,4-滴渗透与具不同抗性水平的不同植物的角质层厚度无关。对植物角质层组成和化学研究发现,一些植物对2,4-滴的敏感性差异很大,但未发现与2,4-滴吸收有相关性。具有不同叶部特性的苜蓿()、葡萄和豌豆施用除草剂后1、3 d或7 d对2,4-滴吸收无差异。最初的理论认为2,4-滴吸收与气孔开口的数量相关。但这就要求有低于大多数除草剂制剂的表面张力,且气孔渗透对2,4-滴吸收的总体贡献不大。
2,4-滴的输导遵循韧皮部装载途径。同化物和韧皮部移动除草剂的一般运动发生在碳水化合物合成(来源)区域至储存或利用区域。源和汇之间的运输由浓度梯度调控启动,使含有溶有溶质和除草剂的水移动。除草剂和溶质可在质外体(细胞间隙)或共质体(细胞质和胞间连丝)运输,输导率和分配各异。2,4-滴在大豆中表现出强烈的共质体移动性,发现主要存在于茎部等输导区域。2,4-滴经离子捕获在植物韧皮部运输。由于韧皮部和木质部/质外体之间的pH水平差异,2,4-滴等弱酸存留于韧皮部。与质外体(pH=5.5)相比,韧皮部(pH=8.5)为碱性,使弱酸去质子化生成负离子,防止其穿过细胞膜,以离子形式存留于韧皮部。2,4-滴的转运方式与其他内吸性除草剂相似,但是特异性激素类除草剂载体的存在使植物体内的2,4-滴移动与大多数内吸性除草剂有所区别。胞间和胞内吲哚-3-乙酸(IAA)移动与2,4-滴一样主要取决于生长素内流和外输载体。在细胞内,可经液泡膜结合蛋白WAT1运输IAA到液泡中调节IAA浓度。细胞间外输载体包括PIN-formed(PIN)和三磷酸腺苷(ATP)结合盒B亚家族(ABCB)成员,内流载体包括AUX1/LAX(auxin resistant 1/like aux1)家族成员。AUX1/LAX仅与质膜结合,ABCB位于质膜和内膜,PIN载体在质膜和内质网(ER)中发现。生长素内流载体AUX1、LAX1和LAX3都可以运输IAA和2,4-滴,外输载体PIN2和PIN7可以运输2,4-滴,而PIN1则不能。与内流载体相比,可引起细胞内2,4-滴浓度增加的外输载体较少。在烟草()细胞中,由于2,4-滴没有外流,IAA的浓度为118 nM,2,4-滴浓度则达1 106 nM。离体马铃薯组织中的2,4-滴浓度为常温溶液的15倍,且处理组织中2,4-滴的外流非常慢。由于2,4-滴对生长素外输载体的亲和力低,扩散速度慢,未检测到也不可能检测到细胞中的2,4-滴外流。细胞内2,4-滴浓度增加使除草活性更高。
2.2.3 植物物种的影响
2,4-滴的吸收和输导可能受不同个体物种特性的影响。已有许多抗性与敏感性个体对2,4-滴吸收或输导关系的研究。一般来说,敏感性和吸收几无相关性,但与输导的相关性较大。
2,4-滴的吸收,物种间敏感性差异,结果并不总是相关的。敏感菜豆的吸收速度比野燕麦(L.)慢。相比之下,抗性燕麦的吸收速度比敏感大豆快,但施用24 h后的水平相等。尽管东方龙葵()具有更高的抗性,但在番茄和东方龙葵中观察到的吸收水平相近。而敏感豌豆对2种不同2,4-滴制剂的吸收比抗性东方龙葵更多。
虽然吸收和抗性之间的关系不清楚,但与敏感物种相比,抗性物种2,4-滴输导较少或分配不同。处理24 h后,总放射性活度仅有5%从抗性燕麦处理叶片中转出,而敏感性大豆则为55%,存在于大豆根部(14.5%)和生长点(22.7%)的C14比燕麦(分别为3.1%和1.6%)多。抗性和敏感物种间的代谢差异(在下文讨论)可以解释观察到的输导差异。与敏感物种相比,将2,4-滴快速转化为可在抗性物种细胞中螯合的代谢物可以抑制除草剂活性成分或代谢物输导。2,4-滴酸的中等亲脂性和弱酸性使其能够在木质部和韧皮部中移动。另一方面,2,4-滴与各种植物化合物的结合会改变这些特性,限制移动。2,4-滴的结合稍后讨论。
某些物种对2,4-滴的另一抗性表现为根系分泌物。对曼陀罗(),honeyvine milkweed [(Nutt.)Britt.]和丝路蓟()的研究表明,这些物种通过向土壤中分泌除草剂提高了自身对2,4-滴的抗性。类似的研究发现,施用2,4-滴后东方龙葵向土壤中分泌了28%,番茄仅有7%。
2.3 化学和应用因素
2.3.1 剂型
除草剂的剂型可影响除草剂的吸收和输导。前文已讨论了2,4-滴的几种剂型。一般来说,2,4-滴酯的吸收比胺类剂型更快。在大叶槭(Pursh)中,与三乙醇胺剂型相比,乙基己酯剂型的吸收最多。东方龙葵和豌豆对2,4-滴异辛基酯的吸收量分别为三乙醇胺盐(2,4-滴+2-羟乙基胺)的2.4倍和1.3倍,但24 h后,剂型间的转运无差异。与二甲胺盐等短链胺相比,向日葵对四环胺(tetracyclamine)和十二烷基胺等长链胺2,4-滴吸收更多。
2.3.2 助剂的影响
助剂可通过增加叶面湿度,减少除草剂液滴表面张力,增大叶面和液滴接触面积等来提高植物对除草剂的吸收。就2,4-滴而言,助剂可提高除草剂吸收,但未观察到对输导有明显影响。根据2,4-滴剂型,不同的助剂可以提高或降低除草剂吸收。助剂可提高大豆和玉米对几种2,4-滴剂型(胺、钠盐和异丙酯)的吸收。添加表面活性剂Armoblen 600可提高2,4-滴三乙醇胺盐的吸收,但会使异辛酯制剂的吸收降低。当添加1.0%吐温80时,honeyvine milkweed对2,4-滴酸的吸收提高了7~8倍,2,4-滴的输导增加了,但增幅不如吸收明显。类似地,添加助剂使东方龙葵对2,4-滴三乙醇胺盐的吸收提高了4.8倍,豌豆提高了1.7倍。2,4-滴三乙醇胺盐或2,4-滴异辛基酯添加助剂对输导无影响。
还进行了助剂性质对2,4-滴吸收影响的研究。与作物油浓缩物(crop-oil concentrate)相比,以有机硅氧烷和乙炔二醇乙氧基化物(acetylinic diol ethoxylate)混合物为表面活性剂时,乳浆大戟()对除草剂吸收提高了19%。环氧乙烷(EO)含量(-CH2CH2O-)与2,4-滴吸收成反比,蚕豆(L.)对含EO=3表面活性剂的除草剂的吸收(85%)比含EO=10(61%)和EO=14(50%)的高。EO值较低即EO链较短,表面活性剂分子极性较小。含C13/C15 (即每分子具13或15个碳原子的碳链,碳越多,表面活性剂疏水性越大)烷醇表面活性剂的2,4-滴的吸收高于C10烷醇,使用辛基酚助剂的吸收增幅最小。但另有研究发现,EO为6~18的表面活性剂不影响野燕麦和菜豆对2,4-滴吸收。助剂性质、叶片特性及两者之间相互作用的复杂性使助剂对2,4-滴吸收的影响难以定论。
2.3.3 应用方法
除草剂应用的目的是将活性成分输送至靶标杂草。此可能受喷雾液滴的大小和载体体积(carrier volume)等几个因素的影响。这些因素可以通过决定靶标喷雾覆盖面积,改变液滴中活性成分的浓度以及影响叶片表面液滴撞击和持留来影响除草剂吸收。减小液滴直径或可提高2,4-滴对某些杂草的防效,激素类除草剂麦草畏和2甲4氯也有相同的研究报道,但液滴直径对草甘膦的药效无影响。与许多其他内吸性除草剂类似,载体体积的减小并不影响2,4-滴的性能,但草甘膦的性能却因此提高了。尚未确定使蚕豆中2,4-滴吸收最高的最佳液滴直径、载体体积或活性成分(ae)浓度,随着2,4-滴用量的增加,吸收效率降低。类似地,西亚大蒜芥()对2,4-滴二甲胺的吸收不受液滴直径影响,但随着液滴直径增加,输导减少。而2,4-滴酸以较小的液滴(0.5 μL相对于10 μL)、较大的总量(100 μL相对于10 μL)于菜豆叶面喷雾施用时吸收最大。2,4-滴液滴浓度不影响吸收,但浓度增加8倍,输导下降10%~14%。因各研究所用方法(如喷嘴类型、喷雾压力、施用器械等)及靶标植物不同,液滴直径和载体体积对2,4-滴吸收和转运的真正影响可能难以确定。
2.3.4 pH
喷雾溶液的pH可影响除草剂的吸收和输导。pH改变对除草剂分子的离子状态有决定性影响,带电荷分子跨越角质层和细胞膜的移动比不带电荷的分子更为困难。由于分子的离子状态将取决于植物内部,故喷雾溶液的pH对吸收的影响大于输导。pH为3时菜豆和向日葵子叶叶面测得的吸收高于pH为5时,pH变化对向日葵吸收的影响更为明显。灯心草粉苞苣(L.)对2,4-滴的吸收在pH 3.5时高于pH 8.5时,向根部的输导(以植物吸收的百分比表示)则在较低pH时更多。
2.4 小结
可改变2,4-滴吸收和输导的因素很多,但仅有几个因素具有一致的影响。在较高的温度和湿度的条件下,植物对2,4-滴的吸收较多。水分胁迫不影响吸收,但会使输导下降。助剂、较低的pH和2,4-滴酯类剂型可使植物对2,4-滴的吸收增加,但通常不影响其输导。2,4-滴吸收与角质层厚度或组成以及植物敏感性几无相关性,但在敏感物种体内2,4-滴输导比抗性物种多。未来对转基因2,4-滴抗性作物中2,4-滴的生理研究,与仅有转基因不同的非转化同源系(isogenic lines)对照,将确定影响2,4-滴的吸收和输导的这些因素是否一致。
3 植物对2,4-滴的代谢
除草剂的选择性很多情况下取决于植物的代谢。植物通常将除草剂代谢为极性更大的物质和不溶残余物。有趣的是,2,4-滴在敏感和抗性物种中的代谢途径有一些共性。虽然敏感物种在某些情况下代谢2,4-滴确实比抗性物种更快,但所产生的代谢产物可能很容易被还原为母体酸。另一方面,抗性物种通常会产生对植物无毒性、不可逆的2,4-滴代谢物。敏感双子叶植物和抗性单子叶植物的2,4-滴代谢过程中产生的代谢产物相似,但是每种代谢产物的量不同,最终抗性单子叶植物中2,4-滴浓度低于双子叶植物。例如2,4-滴和IAA在双子叶植物中都通过改变羧酸基团而被代谢为不稳定的、可逆代谢物,这些代谢物可经酯化作用被还原为活化态。但2,4-滴对单子叶植物的选择性主要与无毒、稳定代谢物的形成和螯合有关,通常经苯基或杂环的修饰来完成。
本节将对2,4-滴代谢进行总结,着重阐述各种途径、所涉及的酶类、产生的代谢产物以及物种间的差异。本节讨论图1中2,4-滴的3种代谢途径,产生的代谢产物及可通过共轭进一步代谢的羟基(图中圆圈)。某些种群对2,4-滴抗性可能是对2,4-滴具有另一种代谢机制,这些种群和抗性机制将在后文进行讨论。近期已有人对2,4-滴酯代谢中的另一个环节——以羧酸酯酶对2,4-滴酯类制剂进行生物活化作了论述,本节仅讨论2,4-滴酸的代谢。
3.1 侧链裂解
已在许多植物中观察到2,4-滴侧链的裂解,但其仅为红醋栗(Syme)、苹果、草莓和欧丁香()等少数植物的主要代谢作用。侧链经单独氧化降解生成乙醇酸和2,4-二氯苯酚。2,4-滴经侧链降解会导致二氧化碳减少,因此二氧化碳的减少是确定2,4-滴进行侧链裂解代谢的指标,在几个主要利用该途径的物种中二氧化碳减少7%~33%,在玉米、大豆、棉花和其他几种物种中则为1%~2%。
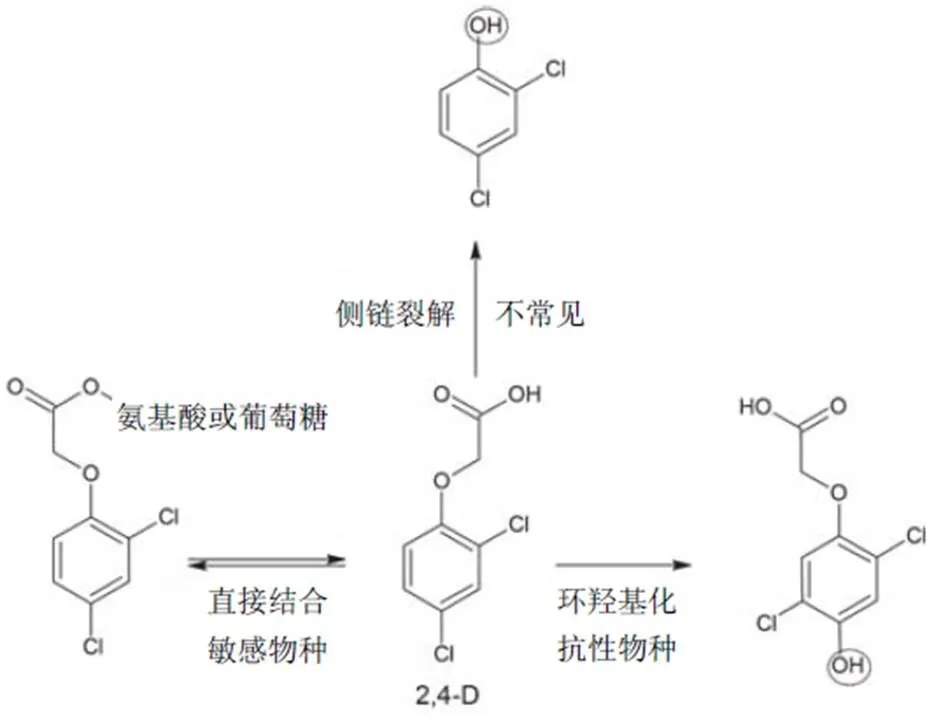
图1 高等植物2,4-滴代谢途径
3.2 直接结合(direct conjugation)
研究最为广泛的代谢机制为2,4-滴与氨基酸和葡萄糖的直接结合。氨基酸(主要是谷氨酸和天冬氨酸)和葡萄糖可与2,4-滴的羧酸基团直接结合,生成氨基酸缀合物或2,4-滴葡萄糖酯。在大豆和其他敏感双子叶植物中以生成氨基酸缀合物为主。在基因作用下IAA与氨基酸直接结合,但同样的酶不能结合作为底物的2,4-滴。氨基酸缀合物的浓度大大超过双子叶植物中生成的其他代谢物和单子叶植物中发现的量。氨基酸缀合物是双子叶植物中形成的第一代谢产物,最初产生谷氨酸缀合物,但随着时间的推移,2,4-滴-谷氨酸缀合物转化为其他代谢物,如2,4-滴-天冬氨酸缀合物和糖缀合物。2,4-滴-葡萄糖缀合物在葡糖基转移酶(GT)的作用下生成葡萄糖酯。葡萄糖是仅有的用于直接结合的糖,本节稍后再讨论在环羟基化中用于生成较大大分子的其他糖类。通常,氨基酸或葡萄糖酯缀合物多见于敏感双子叶植物中,诱导与2,4-滴类似的生长素相关活性,易水解还原为2,4-滴酸。这种活性2,4-滴和可逆2,4-滴缀合物使除草剂对这些物种发挥作用。但在双子叶植物的液泡中又可恢复为2,4-滴缀合物。这可能会使除草剂对这些物种的活性降低。
3.3 环羟基化(ring hydroxylation)
抗性单子叶植物主要通过环羟基化反应代谢2,4-滴。2,4-滴芳环上碳-4位的羟基化导致氯原子迁移或移动到碳-3或碳-5位。由参与几种除草剂代谢和解毒的细胞色素P450酶系催化反应发生环羟基化。由2,4-滴的环羟基化生成的主要代谢物有4-羟基-2,5-二氯苯氧基乙酸和4-羟基-2,3-二氯苯氧基乙酸,这些代谢物在单子叶植物中比双子叶植物中更为常见。已发现其他羟基化代谢物,但最常见的为4-羟基-2,5-滴。在羟基化后,GT酶迅速催化环羟基化代谢物生成-葡萄糖苷(图1)。糖基化后,代谢物可进一步与其他糖(包括丙二酸)结合以形成更大的结构。-丙二酰转移酶(-malonyltransferase,-MAT)催化丙二酰化(malonylation),或有助于结合物抵抗细胞消化。环羟基化代谢途径的产物比2,4-滴亲水性更大,无毒,极性更强,且不能水解还原为2,4-滴。这些无毒、不可逆环羟基化物比其他类型的代谢物在液泡等不同位置更易与木质素、果胶和纤维素等结构型聚合物(structural polymers)螯合或结合。敏感双子叶植物同样生成这些代谢物,但浓度一般极低,表明其他代谢途径的利用率更高。
3.4 小结
植物对2,4-滴的代谢主要通过直接结合和环羟基化进行,少数为侧链裂解(图1)。除草剂的选择性源于每种植物的代谢途径。2,4-滴与氨基酸或葡萄糖直接结合的植物毒性代谢产物可水解生成2,4-滴,这在敏感双子叶植物中更为常见。2,4-滴环羟基化生成一种稳定的、可被GT和O-MAT酶进一步代谢的无或部分植物毒性代谢物,此在抗性单子叶植物中较为常见。在双子叶植物和单子叶植物中,代谢物和游离的2,4-滴与结构型聚合物结合,但在液泡中仅发现代谢物。与正常品种相比,未来对转基因2,4-滴抗性作物中2,4-滴代谢的研究有望揭示这些同源系的不同代谢率、涉及的酶及产生的代谢产物。
4 作用机制
4.1 3类生长素受体
2011年加拿大科学家报道了一类新的生长素受体蛋白质,使已知的生长素受体总数达到3种。近年来,已对这3种生长素受体或辅助受体体(coreceptor systems)进行了表征,同时阐明了它们对植物生长素介导信号传导的影响。这3种生长素受体包括:⑴主要位于内质网(ER)和外层细胞膜/质外体界面(apoplast interface)的生长素结合蛋白1(ABP1);⑵位于细胞核中的生长素信号F-box (TIR1/AFB)受体蛋白同源物;以及最近的⑶ S期激酶相关蛋白2 (SKP2),其也位于细胞核中,其生长素信号传导应答表现为细胞分裂。除了亚细胞定位不同外,这3种生长素受体在细胞扩增、细胞分裂和植物生长发育调节中的初步功能作用不同。
位于质膜和内质网的ABP1蛋白在生长素感应和信号传导应答中的功能和作用已在拟南芥中得到了详细的记录和建立,但最近发现ABP1并非生长素[IAA和1-萘乙酸(NAA)]信号传导途径、生长素响应基因表达或正常生长条件下拟南芥发育调节的必要组分。此外,最近有人重新研究了可能由于拟南芥中突变型等位基因-5引起的生长素表型效应。在对-5基因系的整个基因组进行测序时,研究人员发现了可能对这些表型有影响的其他无关联位点突变(即与-5无关),从而对ABP1在生长素感应和信号传导中的实际作用和功能产生了更多怀疑。这些最新研究结果表明,应重新审视和深入研究ABP1在植物生长、发育和生长素应答基因表达(在正常和胁迫条件下)中的作用,特别是涉及双子叶杂草ABP1在人工合成激素类除草剂应答中的可能作用或激素类除草剂可能的抗性机制。但ABP1在IAA或2,4-滴信号传导中的作用尚未在拟南芥以外的双子叶植物中建立。
尽管ABP1主要位于内质网上,但是推测ABP1在细胞周围的弱酸性质外体中结合和感应生长素,其可能参与质膜上的膜电位和离子通量的快速调节,控制生长素诱导的细胞扩增。最近研究发现,细胞表面生长素感应ABP1识别位于质膜的跨膜类受体激酶——跨膜ABP1互作伴侣形成细胞表面生长素感应复合体,在胞浆(cytosol)内进行信号传导,调节已知ABP1结合生长素依赖型应答下游。虽然ABP1被认为主要作为调节细胞表面快速、非转录应答的关键因子,但其也可能影响细胞核中的生长素调节转录应答,见下文对TIR1/AFB蛋白生长素受体类的讨论。
细胞核生长素受体TIR1/AFB蛋白家族与生长素或2,4-滴结合作为“分子胶(molecular glue)”,参与了一种新的机制,即当生长素浓度高时转录抑制因子Aux/IAAs(抑制生长素应答基因表达)迅速降解,通过“抑制释放”使许多生长素响应基因转录激活。最近在研究可能参与人工合成激素类除草剂结合拟南芥TIR1/AFB蛋白受体的建立以及生长素结合差异性的结构基础方面取得了进展。例如,与TIR1相比,AFB4和AFB5蛋白具有更明显的差异(从一级结构的角度来看),AFB5与吡啶羧酸类、二氯喹啉酸和麦草畏优先结合,而TIR1、AFB1、AFB2和AFB3则优先结合2,4-滴。有趣的是,在拟南芥植物中生长素转运突变系对2,4-滴具有抗性,对麦草畏敏感,AFB突变系和均对麦草畏具有抗性,但只有-1系抗2,4-滴。此外,双突变系/对麦草畏的抗性具有叠加效应。因此,人工合成激素类除草剂对生长素转运蛋白或细胞核生长素受体的结合亲和力差异可能引起植物物种间的抗性差异或双子叶杂草中的广泛或特异性交互抗性机制差异,这取决于田间条件下选择的AFB或生长素转运蛋白的突变。
最近发现的生长素受体SKP2参与细胞周期进程中核转录因子的降解。在细胞周期中,为了引发下一个阶段,需要降解一些转录因子和其他蛋白质。SKP2a蛋白与生长素的结合增强了SKP2a与细胞分裂相关转录因子间的相互作用,从而促进其降解,同时使细胞分裂继续进行。总之,除了TIR1/AFB依赖型生长素感应机制外,SKP2还可能为细胞核中生长素的最终响应提供替代途径。
4.2 植物对IAA和2,4-滴响应的差异
在拟南芥幼苗中,IAA及其同系物2,4-滴表现抑制作用的浓度相当,但IAA和2,4-滴的剂量-反应曲线完全不同。例如,2,4-滴(低纳摩尔浓度时)对根伸长抑制的剂量-反应曲线最初比IAA的斜率大,表明双子叶植物对IAA的响应略弱于2,4-滴。植物全株(whole-plant)对IAA和2,4-滴响应差异有3种可能的解释:⑴生长素感应的差异,⑵细胞运输机制的差异和⑶生长素代谢和动态平衡(或2,4-滴的解毒反应)的差异。由于细胞运输机制和生长素感应的差异似乎不足以造成拟南芥根系对2,4-滴的敏感性高于IAA(即通常2,4-滴对于这些进程而言是较差的底物),似乎最有可能的解释是生长素代谢和维持生长素动态平衡的机制的定性和定量差异导致了植物全株对2,4-滴和IAA间敏感性的差异。例如,假设通过苯环羟基化随后葡萄糖缀合进行2,4-滴解毒赋予杂草抗性,而敏感双子叶植物只能与氨基酸或糖进行可逆缀合反应改变羧酸基团。此外,在目前已知的各种生长素受体和细胞生长素转运蛋白中,IAA是比2,4-滴更适合的底物或具有与2,4-滴相当甚至更大的结合亲和力。
直接比较放射性标记的IAA和2,4-滴的向基和向顶运输的长距离转运研究将大大有助于人们了解单个生长素内流和外输转运蛋白的总体差异以及这些差异对胞间和胞内运输、双子叶植物对天然和人工合成激素类敏感性的可能影响。如上所述,在细胞内,IAA浓度由液泡膜结合蛋白WAT1转运到液泡进行调节。细胞间生长素运输和激素类除草剂药效受前文2,4-滴输导中所述的特定外输载体的影响。对麦草畏等其他激素类除草剂具有特异性的植物细胞和组织内转运蛋白的研究报道较少,但如前所述,拟南芥生长素转运突变系对2,4-滴具有抗性,但对麦草畏敏感。
一组被称为“phytotropins”的有趣化合物可抑制细胞内IAA(由PIN生长素外输载体蛋白促进)和人工合成激素类流出。这使茎和根部分生组织的生长组织中产生异常高浓度的生长素。已发表的有关氟吡草腙(diflufenzopyr)等具phytotropin活性的除草化合物的研究成果仅报道了与人工合成激素类除草剂麦草畏、二氯喹啉酸和毒莠定的协同作用,或者缩氨基脲(semicarbazone)SCB-1对NAA细胞流出的影响。根据对IAA、NAA、麦草畏、二氯喹啉酸全株、长距离输导的比较研究,如果可以识别植物细胞中抑制苯氧乙酸类除草剂基部流出的特异性PIN抑制剂,2,4-滴必会大大增强。
4.3 生理活性
如上所述的生长素响应基因的去抑制化启动了植物体内的一系列生理响应,最终导致敏感双子叶植物死亡。有人将敏感双子叶植物的生理响应分为3个阶段:刺激、抑制和衰退(decay)。刺激阶段在几分钟内开始,其特征为除草剂施用后的合成生长素浓度直接大幅度增加,几小时内植物出现明显症状。在此阶段刺激乙烯产生,脱落酸(ABA)超累积(hyperaccumulates)以及离子通道和ATP酶代谢活化。几小时内开始出现偏上性生长、组织膨大和茎扭曲等异常、不稳定生长症状。在抑制阶段,活性氧过量产生,气孔闭合,生理防御-胁迫响应的诱导使植物生长和发育所需的氨基酸、淀粉、核酸和其他主要代谢物的总产量减少。这些不稳定的级联(cascade)事件导致植物停止生长并最终走向衰退阶段:在处理后3 d内出现因叶绿体、膜和植物维管束被破坏而造成的植物死亡和组织/细胞损伤。
最近的分子生理学研究已鉴定了番茄中几个响应2,4-滴、ABA和乙烯的转录抑制子AUX/IAA蛋白家族生长素响应基因(和)。如上所述,这些研究结果有助于进一步了解2,4-滴的作用机制以及引起敏感双子叶植物中毒的生长素、乙烯和ABA信号传导途径间的相互作用。有趣的是,转基因番茄植物中基因的过表达可诱导偏上性生长、影响气孔细胞分化和叶密度等与2,4-滴处理相似的表型,基因则不能。另外,根据根伸长测定发现,过表达(通过35S启动子)或低表达(通过RNA干扰)转基因品系对外源施用ABA的敏感性发生改变,但是乙烯反应表型没有变化。最近的这些研究结果表明,2,4-滴诱导表达的可能在番茄体内介导2,4-滴或其他人工合成激素类除草剂的下游植物毒性效应中具有重要作用,这与由几种植物激素触发的不稳定生理效应相一致。
鉴于这些复杂的植物信号机制、植物激素应答和变化的基因表达效应,亚致死剂量2,4-滴的应用或在禾本科杂草等相对不敏感物种中应用仍可因各种酶上调或下调引发许多生理响应。当2,4-滴与其他除草剂或农药联合施用时,可观察到其中一些效应。
4.3.1 禾本科杂草苗后防除中的拮抗作用
已有许多有关野燕麦、栽培燕麦和黑麦草()叶面施用2,4-滴桶混制剂(tank mixes)与乙酰辅酶A羧化酶(ACCase)抑制剂类除草剂特别是禾草灵(diclofop-methyl)存在拮抗作用的案例报道。但在小麦、野燕麦或栽培燕麦根施禾草灵和2,4-滴对根生长复合作用的研究中,并没有发现两者存在拮抗作用。2,4-滴酯类与禾草灵的拮抗作用比胺类弱,有人将这一差异部分归因于与2,4-滴胺混用时diclofop吸收较少。随后的研究发现,当haloxyfop与2,4-滴混用时,假高粱[(L.) Pers.]体内活性haloxyfop代谢较多,输导较少。有不少文献报道2,4-滴处理可增加水稻和小麦体内P450基因的表达,上调作为天然底物的小麦P450酶系[在不同位置羟基化月桂酸(lauric acid)的月桂酸羟化酶(LAH)]代谢活性或使光系统II抑制剂绿麦隆(chlortoluron)环-甲基-羟基化和-脱甲基化。有趣的是,小麦LAH也可代谢禾草灵酸(diclofop acid),导致小麦对其快速解毒,产生耐药性。虽然2,4-滴诱导小麦体内特异性LAH活性和禾草灵酸羟化活性之间没有直接的因果关系,但2,4-滴诱导同一P450的表达和活性似乎可使小麦甚至黑麦草属(spp.)和看麦娘属(spp.)等禾本科杂草体内月桂酸和禾草灵酸羟基化,从而出现拮抗作用。
2,4-滴叶面施用也可以加速ACCase抑制剂类或乙酰乳酸合酶(ALS)抑制剂类等其他除草活性成分的代谢解毒。使用ACCase抑制剂类或ALS抑制剂类除草剂苗后防除禾本科杂草时,除草剂间的这种拮抗作用是由于2,4-滴激发基于P450的代谢降低了用于防除禾本科杂草除草剂的活性,从而仿效频繁赋予禾本科杂草抗性或谷类作物耐受性的代谢机制。但2,4-滴对禾本科杂草中除草剂代谢的这种激发作用似乎是短期的,因为如果2,4-滴和苗后graminicide应用间隔24~72 h,则无明显拮抗作用。
大多数研究表明,2,4-滴可加快禾草灵酸解毒代谢为极性更大的缀合物并减少向分生组织的输导。2,4-滴除了防除禾本科杂草时与ACCase抑制剂类除草剂存在拮抗作用外,在防除多年生杂草假高粱时还与草甘膦(与2,4-滴或麦草畏桶混应用)存在拮抗作用。由于草甘膦是一种非选择性除草剂,在植物体内代谢缓慢,故上述拮抗作用的机制与草甘膦吸收和根系转运减少有关。
现阶段的研究需要更多关注2,4-滴对苗后应用的杀虫剂、杀菌剂或非激素类(nonauxinic)植物生长调节剂等其他农药活性的潜在影响,反之亦然。从理论上说,如果2,4-滴可提高P450或其他农药降解酶(类似于除草剂安全剂)的活性,则可能需要深入研究叶面施用作物化学保护剂的生物动力学,特别是在继大豆和棉花之后商业化的新颖2,4-滴抗性作物中。
4.3.2 阔叶杂草苗后防除的协同作用
与苗后防除禾本科杂草的拮抗作用相反,防除多年生杂草田旋花时2,4-滴和麦草畏与草甘膦具有协同作用。如前所述,草甘膦在植物体内的代谢不明显,而2,4-滴对双子叶杂草具有苗后防除活性,因此获得协同作用的机制可能比禾本科杂草更为复杂。有人对田旋花的研究表明,2,4-滴和草甘膦桶混制剂不仅使2,4-滴的吸收增加,2,4-滴和草甘膦向根部的输导也增加了。但此研究没有说明这些桶混制剂中是否含有助剂。还有人的田间和温室研究表明,阳离子、牛脂胺(tallow amine)表面活性剂可优化叶面吸收和草甘膦的活性,因此在桶混制剂中添加这些阳离子表面活性剂也可能增加2,4-滴或麦草畏吸收,导致在双子叶杂草中出现协同作用。
4.3.3 2,4-滴桶混制剂的作物安全性
如上所述,2,4-滴的应用使杂草中P450酶的活性增强,导致其他活性成分代谢加速。就作物而言,这是一种有利的除草剂相互作用,即2,4-滴可以通过加速除草剂解毒来保护作物,避免除草剂带来的药害,其作用机制与除草剂安全剂类似。例如,在杂草抗性研究中有机磷(OP)类杀虫剂常用于抑制代谢除草剂的P450酶,有助于判断抗性机制的类型。但从作物安全的角度来看,代谢ALS抑制剂类等除草剂的P450酶的抑制会引起除草剂协同作用,增加了作物药害和产量损失的可能性。以含有2,4-滴和烟嘧磺隆桶混制剂处理玉米幼苗(之前以有机磷杀虫剂特丁硫磷进行土壤处理),可避免特丁硫磷和烟嘧磺隆互作而引起的药害。因此,当作物体内的P450水平不足以代谢其他除草剂活性成分时,2,4-滴可充当除草剂安全剂。例如,甲磺隆与2,4-滴或麦草畏桶混应用时,对高粱[(L.) Moench ssp.]的药害减轻,但对裂叶牵牛(Jacq.)或青麻(Medik.)的防效并未下降。
(未完待续)
10.16201/j.cnki.cn31-1827/tq.2017.03.05
TQ457
A
1009-6485(2017)03-0031-08
筱禾,女,工程师。Tel: 021-64387891-201。
2016-06-01。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
空间科学学报(2021年1期)2021-05-22
生物学通报(2021年4期)2021-03-16
科学大众(2020年12期)2020-08-13
今日农业(2019年15期)2019-01-03
数码世界(2018年7期)2018-12-24
现代园艺(2017年19期)2018-01-19
中国果菜(2016年9期)2016-03-01
中国蔬菜(2015年9期)2015-12-21
营销界(2015年23期)2015-02-28
