
同脉缟蝇亚属种团间的系统发育关系初探
2017-06-05 14:59张梦靖申荣荣李顺德王永杰何国玮
环境昆虫学报 2017年2期
史 丽,张梦靖,申荣荣,李顺德,白 娟,王永杰,何国玮,崔 悦
(内蒙古农业大学农学院,呼和浩特 010019)
同脉缟蝇亚属种团间的系统发育关系初探
史 丽*,张梦靖,申荣荣,李顺德,白 娟,王永杰,何国玮,崔 悦
(内蒙古农业大学农学院,呼和浩特 010019)
选取2个线粒体基因COI和16S-rRNA,2个核基因Elongation factor 1-α和28S-rRNA,初步探讨同脉缟蝇亚属种团间的系统发育关系,研究表明,翅斑的减少是趋于原始的性状;基于线粒体基因COI和16S-rRNA构建的NJ、ME、MP、ML系统树中,MP树更与形态分类的研究结果一致;而基于Elongation factor 1-α和28S-rRNA基因构建的NJ、ME、MP、ML系统树反映的系统发育关系完全相同;以同脉缟蝇亚属18种为代表构建的6个种团的系统发育关系并没有呈现出一致的规律性,不能从分子水平验证根据已有的外部形态特征划分种团的可靠性。
缟蝇科;同脉缟蝇亚属;系统发育;种团
缟蝇主要营菌食性和腐食性生活,在降解有机质、保护环境、维持生态平衡中起着非常重要的作用;部分属的成虫可访花,有助于植物传粉(Silva and Mello, 2008)。缟蝇对环境变化敏感,是陆生生态系统环境变化评价的指示生物之一(Reddersen, 1995)。同脉缟蝇亚属隶属于双翅目缟蝇科Lauxaniidae同脉缟蝇亚科Homoneurinae同脉缟蝇属,是缟蝇科中具有生态和经济重要性的一大类群,物种多样性丰富。我国已知20种团,145种(Shi and Yang, 2009, 2014)。种团的建立是基于翅斑和雄性外生殖器相结合的特征进行人为划分的,本实验旨在从分子手段,检验已划分种团的可靠性,并依据分子数据,推导种团间鉴别的关键形态特征,这对于深入分析同脉缟蝇亚科同脉缟蝇属不同亚属间的系统发育关系具有重要的借鉴意义。
1 材料与方法
1.1 实验材料
野外采集的标本,直接浸泡于无水乙醇中保存,回到实验室后,更换一次无水乙醇,然后,置于-20℃冰箱中保存。标本信息见下表1。

表1 物种信息
1.2 实验方法
1.2.1 标本解剖和鉴定
在光学解剖镜下,用眼科专用手术剪刀小心剪下腹部相应部分或从胸部和腹部连接处取下整个腹部;将解剖后的腹部置于10% NaOH溶液中,8 h,待腹部大部分肌肉和脂肪溶解后取出,用清水小心漂洗,并将其置于盛有适量水或甘油的培养皿内的小脱脂薄棉层上,以待观察;在观察结束后,用解剖针将剥离的外生殖器轻轻拨入后腹部,用蘸有适量甘油的小包裹,将之转入外生殖器专用塑料小管中保存。结合相关种类鉴定资料,最终鉴定到种。
1.2.2 DNA提取
采用血液/细胞/组织基因组DNA提取试剂盒(离心柱型)TIANGEN(李轩昆,2015)。
1.2.3 PCR扩增与测序
引物合成和双向测序均由上海生物工程有限公司完成。长度在1 kb以内的片段,釆用PCR反应体系(50 μL)扩增,反应程序为:94℃预变性3 min,95℃变性30 s,48℃-55℃退火45 s,72℃延伸1-2 min,扩增35个循环,72℃终延伸7 min,4℃保存;长度在1 kb以上的片段,釆用PCR反应体系(25 μL),反应程序为:95℃预变性30 s,95℃变性10 s,48℃-55℃退火50 s,65℃延伸2-6 min,扩增45个循环,65℃终延伸10 min,4℃保存(张广宇,2014)。

表2 用于扩增分子标记的引物
1.2.4 数据处理和分析
选取缟蝇亚科Lauxaniinae双鬃缟蝇属Sapromyza和黑缟蝇属Minettia、斑腹蝇科Chamaemyiidae斑腹蝇属Chamaemyia和沼蝇科Sciomyzidae沼蝇属Sciomyza作外群,将同脉缟蝇亚科Homoneurinae同脉缟蝇亚属Homoneura的种类的序列,用Mega 7.0中的CLUSTALX进行序列比对(均用默认参数设置)。将比对后的结果转换成Mega格式,用Mega 7.0基于Kimura-2-Parameter(K2P)模型,计算两两个体之间的遗传距离,使用1000次重复检测的Bootstrap值表示树上各节点的支持率,采用邻接法(NJ)、最小进化法(ME)、最简约法(MP)构建同脉缟蝇亚属种团间系统发育树。
2 结果与分析
采用MEGA 7.0系统发育分析软件,构建同脉缟蝇亚属种团间的系统发育树。
2.1 基于COI基因的系统发育树
图1显示,系统树的最顶部聚类的是新北区分布的种类(philadelphica,fraterna,bispina,incerta,biumbrata),包括了abnormisgroup[H.(H.)philadelphica和H.(H.)incerta]与singularisgroup[H.(H.)fraterna],sauterigroup[H.(H.)bispina],quinquenotatagroup[H.(H.)biumbrata];其中,abnormisgroup[H.(H.)philadelphica]和singularisgroup[H.(H.)fraterna]聚为一支,abnormisgroup[H.(H.)incerta]与quinquenotatagroup聚为一支。中部是印度马来亚区分布的种类(dorsacerba,apicitrunctata,fengyangshanica),包括了singularisgroup [H.(H.)dorsacerba和H.(H.)fengyangshanica],calopteragroup[H.(H.)apicitrunctata],这一支的特点是纵脉R4+5上有2个以上的褐色中斑或端前斑。下部是古北区分布的种类(pictipennis,stackelbergiana,patelliformis,septentrionalis),包括了tibetensisgroup[H.(H.)pictipennis],quinquenotatagroup[H.(H.)stackelbergiana],unguiculatagroup[H.(H.)patelliformis],sauterigroup[H.(H.)septentrionalis];其中,tibetensisgroup和quinquenotatagroup聚为一支,而H.(H.)patelliformis单独为一支,可能是因为其翅前缘黑色短鬃不伸达R4+5,这一特征也为Sapromyza属的部分种类所特有。从树的顶端至底端,种团内种类的翅斑数量呈现递减的趋势。这4个系统树中,MP树更与形态分类相吻合。
2.2 基于16S-rRNA基因的系统发育树
图2显示,仅有H.(H.)apicitrunctata和H.(H.)dorsacerba是印度马来亚区分布,其余种类均为古北区分布。顶部一支包括calopteragroup[H.(H.)apicitrunctata]或singularisgroup[H.(H.)dorsacerba],sauterigroup[H.(H.)remmi],quinquenotatagroup[H.(H.)biumbrata],singularisgroup[H.(H.)dilecta];底部一支包括unguiculatagroup[H.(H.)thalhammeri和H.(H.)patelliformis],sauterigroup[H.(H.)mediospinosa]。而singularisgroup[H.(H.)dorsacerba]或calopteragroup[H.(H.)apicitrunctata]单独一支,可能是与其印度马来亚区分布有关,但证据不充分。这4个系统树中,MP树更与形态分类相吻合。
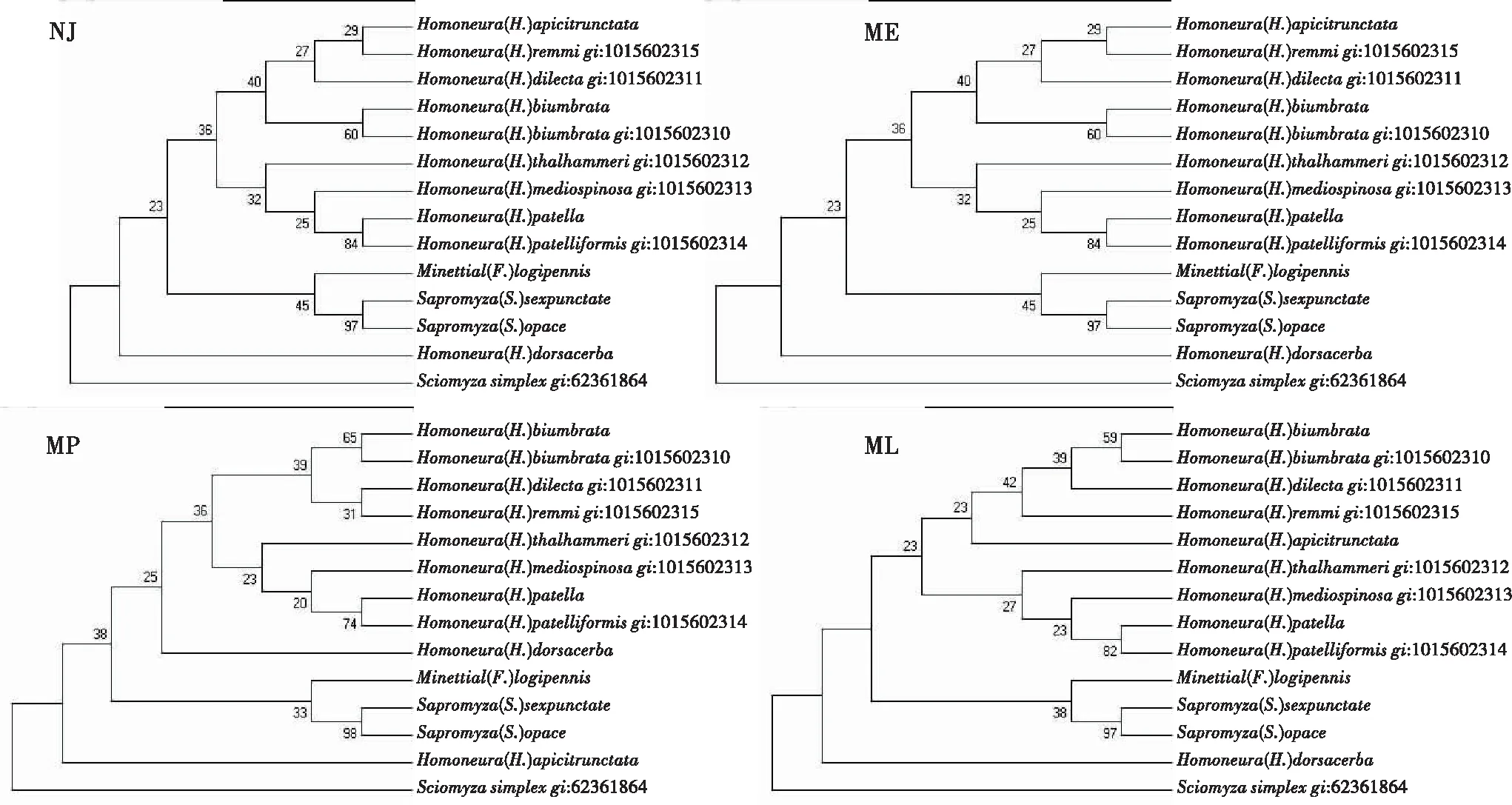
图2 基于16S-rRNA基因的系统树Fig.2 Cladistic tree based on the 16S-rRNA gene
2.3 基于28S-rRNA基因的系统发育树
图3显示,系统树顶部quinquenotatagroup[H.(H.)biumbrata]与sauterigroup[H.(H.)remmi]、singularisgroup[H.(H.)dilecta]聚为一支,singularisgroup[H.(H.)dorsacerba]和calopteragroup[H.(H.)apicitrunctata]聚为一支,而sauterigroup[H.(H.)mediospinosa和H.(H.)septetrionalis]、unguiculatagroup[H.(H.)thalhammeri和H.(H.)patelliformis]、tibetensisgroup[H.(H.)pictipennis]分别聚为一支。与COI和16S-rRNA基因的系统树相比,所选四种方法的系统树呈现的系统发育关系完全相同,且有一定的规律性,比如翅斑减少这一特征更为原始,或许在自然界中,昆虫翅斑增加,呈现拟态或保护色,更容易繁衍生息。
2.4 基于Elongation factor 1-α基因的系统发育树
图4显示,singularisgroup[H.(H.)dilecta]和quinquenotatagroup[H.(H.)biumbrata]的系统发育关系最近,calopteragroup[H.(H.)apicitrunctata]与sauterigroup[H.(H.)interstincta,H.(H.)mediospinosa,H.(H.)remmi]和unguiculatagroup[H.(H.)patelliformis]的关系最远。与28S-rRNA基因的系统树相比,所选4种方法的系统树呈现的系统发育关系也是完全相同,且有一定的规律性。但是,由于这个基因比前述3个基因更难扩增,因此,可选昆虫种类明显少。
3 结论与讨论
选取4种基因,对同脉缟蝇亚属内种团间的系统发育关系进行初探,结果表明,翅斑的减少是趋于原始的形状;基于线粒体基因COI和16S-rRNA构建的NJ、ME、MP、ML系统树中,MP树更与形态分类的研究结果一致;而基于Elongationfactor 1-α和28S-rRNA基因构建的NJ、ME、MP、ML系统树反映的系统发育关系完全相同;以同脉缟蝇亚属18种为代表构建的6个种团的系统发育关系并没有呈现出一致的规律性,不能从分子水平验证根据已有的外部形态特征划分种团的可靠性。
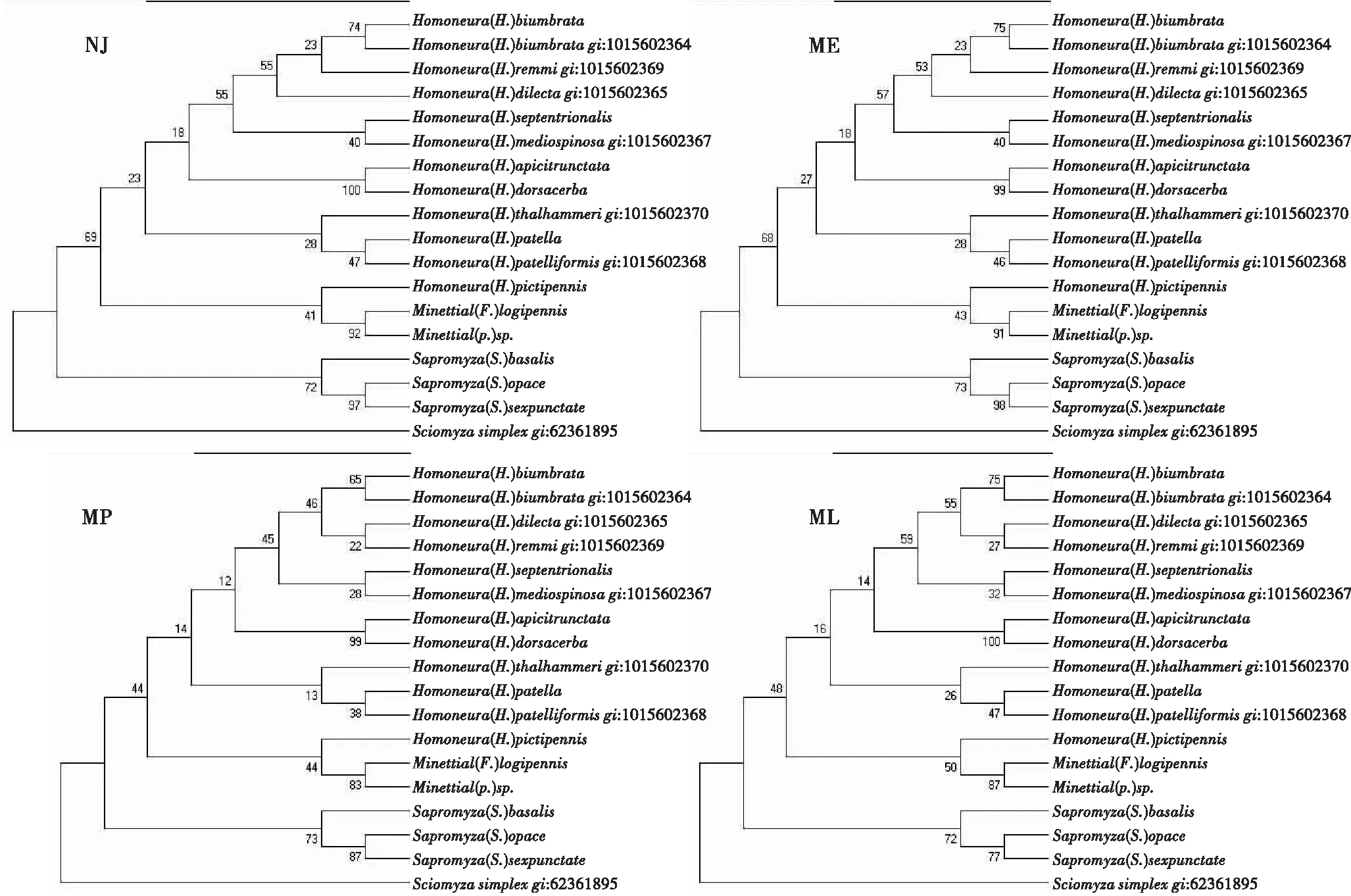
图3 基于28S-rRNA基因的系统树Fig.3 Cladistic tree based on the 28S-rRNA gene
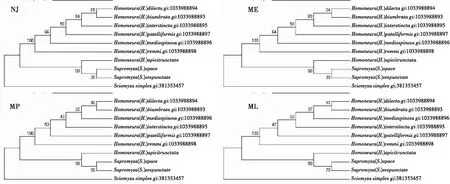
图4 基于Elongation factor 1-α基因的系统树Fig.4 Cladistic tree based on the Elongation factor 1-α gene
标本野外采集过程中,使用了乙酸乙酯,可能存在使用过量的问题,且虫体接触到了乙酸乙酯,因此,尽管实验采用了当年采集的新鲜标本,提取DNA后,测序结果并不如预期的那么理想,这大大的限制了原实验计划中的样本的选择。Elongation factor 1-α基因片段较难扩增成功,需要在引物设计和PCR扩增条件方面进行改进。文中涉及的Homoneura(H.)incerta、Homoneura(H.)fraterna、Homoneura(H.)bispina在Miller(1977)的新北区同脉缟蝇亚属种团划分的研究论著中,分别隶属于incertagroup、fraternagroup和bispinagroup中;而Homoneura(H.)dilecta、Homoneura(H.)remmi在Papp(1978)关于古北区同脉缟蝇种团划分的研究论文中,分别隶属于notatagroup和minorgroup;这两位学者的种团划分标准主要是依据雄性外生殖器各部分的特征。同脉缟蝇属是缟蝇科中最大的属,而同脉缟蝇亚属是这个属中最大的亚属,世界已知600余种,如果能选择合适的特征形状,正确划分种团,将为同脉缟蝇亚属世界区系的修订奠定基础。
References)
Chi Y, Zhi Y, Wang SD,etal. The DNA barcoding of COI gene in some species of Tachininae [J].JournalofShenyangNormalUniversity, 2011, 29 (3): 434-438. [池宇,智妍,王诗迪,等. 寄蝇亚科部分物种线粒体COI基因DNA条形码分类[J]. 沈阳师范大学学报,2011,29 (3): 434-438]
Li XQ. Studies on the Comparative Mitochondrial Genomics and Phylogeny of Cyclorrhapha (Insecta: Diptera) [D]. Beijng: China Agricultural University, 2015: 1-179. [李轩昆. 环裂亚目昆虫比较线粒体基因组学及系统发育研究[D]. 北京: 中国农业大学, 2015:1-179]
Miller RM. Taxonomy and biology of the Nearctic species ofHomoneura(Diptera: Lauxaniidae). 2. SubgenenusHomoneura[J].IowaStateJournalResearch, 1977, 52 (2): 177-252.
Papp L. Constribution to the revision of the Palaearctic Lauxaniidae (Diptera) [J].AnnalesHistorico-NaturalesMuseiNationalisHungarici, 1978, 70: 213-231.
Reddersen J. Feeding biology of fungivorous insects from Danish cereal fields [J].Pedobiologia, 1995, 39 (4): 370-384.
Semelbauer M. Molecular phylogeny of lauxaniid flies (Diptera, Cyclorrhapha) confirms non-monophyly ofSapromyzaFallén 1810 [J].InsectSystematics&Evolution, 2016, 47 (4), 389-409.
Shi L, Yang D. Notes on species groups of subgenusHomoneurafrom China with descriptions of two new species (Diptera, Lauxaniidaae) [J].ActaZootaxonomicaSinica, 2009, 34 (3): 462-471.
Shi L, Yang D. Supplements to species groups of the subgenusHomoneurain China (Diptera: Lauxaniidae:Homoneura), with description of twenty new species [J].Zootaxa, 2014, 3890 (1): 1-117.
Silva VC, Mello RL. Occurrence ofPhysoclypeusfarinosusHendel (Diptera: Lauxaniidae) in flowerheads of Asteraceae (Asterales) [J].NeotropicalEntomology, 2008, 37 (1): 92-96.
Zhao GY. DNA Barcoding of Reduviidae (Insect: Hemiptera) [D]. Beijing: China Agricultural University, 2014: 1-160. [赵光宇. 猎蝽科昆虫条形码研究[D]. 北京: 中国农业大学, 2014: 1-160]
Preliminary study on phylogeny of species groups in subgenusHomoneura
SHI Li*, ZHANG Meng-Jing, SHEN Rong-Rong, LI Shun-De, BAI Juan, WANG Yong-Jie, HE Guo-Wei, CUI Yue
(College of Agronomy, Inner Mongolia Agricultural University, Hohhot 010019, China)
Based on two mitochondrial genes (COI and 16S-rRNA) and two nuclear genes (Elongation factor 1-alpha and 28S-rRNA), the phylogenetic relationships among six species groups of subgenusHomoneurawere preliminarily studied. The results showed that decreasing in quantity of wing spots is a trend to origin; the phylogeny presented from MP could keep better accord with morphological taxonomy than others among the NJ, ME, MP and ML builted by mitochondrial genes (COI and 16S-rRNA), while all NJ, ME, MP and ML builted by the nuclear genes (Elongation factor 1-alpha and 28S-rRNA) phylogeny reflected the same phylogenetic relationships. The phylogenetic relationships from 18 representative species of 6 species groups of subgenusHomoneuracould not show some consistent regularity, so the reliability of the existing species group constructed with artificial classification could not be verified from the molecular level.
Lauxaniidae;Homoneura; phylogeny; species groups
国家自然科学基金(31260525);内蒙古农业大学学生科学创新基金(DC201506);内蒙古杰出青年培育基金(2015JQ03)
史丽,女,1976年生,教授,研究方向为昆虫系统进化与生物多样性
*通讯作者Author for correspondence, E-mail: lirui2003@imau.edu.cn
Received: 2016-12-30; 接受日期Accepted: 2017-02-17
Q963
A
1674-0858(2017)02-0351-06
史丽,张梦靖,申荣荣,等.同脉缟蝇亚属种团间的系统发育关系初探[J].环境昆虫学报,2017,39(2):351-356.
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
疯狂英语·新读写(2021年10期)2021-12-07
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
奥秘(2019年8期)2019-08-28
收藏界(2018年1期)2018-10-10
创新作文(小学版)(2018年31期)2018-05-16
商周刊(2017年7期)2017-08-22
摄影之友(影像视觉)(2017年1期)2017-07-18
小学阅读指南·低年级版(2016年6期)2016-05-14
