
短梳角野螟的形态描述及幼虫虫龄判断
2017-06-05 14:20李小敏刘君昂欧阳博文董文统周国英
植物保护 2017年3期
李小敏, 刘君昂, 欧阳博文, 董文统, 周 斌, 周国英
(中南林业科技大学,森林有害生物防控湖南省重点实验室;经济林培育与保护省部共建教育部重点实验室, 长沙 410004)
短梳角野螟的形态描述及幼虫虫龄判断
李小敏, 刘君昂, 欧阳博文, 董文统, 周 斌, 周国英*
(中南林业科技大学,森林有害生物防控湖南省重点实验室;经济林培育与保护省部共建教育部重点实验室, 长沙 410004)
短梳角野螟Meroctenatullalis(Walker)是为害红花天料木HomaliumhainanenseGagnep的重要食叶害虫,其形态描述和幼虫准确分龄是开展害虫生物生态学及林间防控研究的重要依据。本文采用林间采样与室内饲养相结合的方法,首次对短梳角野螟卵、幼虫、蛹和成虫的形态及生活习性和生活史进行调查和描述。并通过测定头宽、头长和体长3项分龄指标,利用SPSS Statistics19.0统计软件对数据进行频次分析,结合戴氏法则和线性回归等试图探究准确划分幼虫虫龄的可靠方法。结果表明:短梳角野螟幼虫随龄期增长,幼虫颜色由浅黄绿色逐渐变为深绿色,头部由肉色逐渐变深,最后形成外围红褐色中间深肉色。通过频次分析统计,测得短梳角野螟幼虫虫龄为5龄;头宽可作为最佳分龄指标,且头宽测量指标平均值自然对数与幼虫龄期呈线性关系(y=0.181 6x-0.602 3,R2=0.999 1)。头长和体长统计检验,符合戴氏法则,但频次分析5个峰值不是很明显,且有部分区域重叠,只能作为划分虫龄的辅助指标。本研究确定了短梳角野螟各虫态的形态特征及划分幼虫龄期的最佳指标,以期为该虫林间防治提供生物学依据。
短梳角野螟; 形态特征; 幼虫虫龄; 生活史; 红花天料木
短梳角野螟Meroctenatullalis(Walker)属鳞翅目Lepidoptera草螟科Crambidae梳角野螟属MeroctenaLederer,在我国广泛分布于海南、广东、广西、云南及福建等地区;国外主要分布在越南、菲律宾、印度、斯里兰卡、印度尼西亚等东南亚国家,是红花天料木Homaliumhainanense Gagnep的一种重要食叶害虫。该虫主要以幼虫卷叶为害,喜食抽芽嫩叶,食量大,单株虫口数量可达几十到上百,为害严重时可在短时间内把整株树嫩叶吃光,不仅影响林分景观和红花天料木的健康生长,而且造成严重的死苗,从而引起经济损失。
红花天料木,又称母生、多宝树,是我国最珍贵用材树种,海南特类材,具有重要的木材及药用价值,是海南重点保护乡土树种之一,目前材源严重短缺。关于红花天料木虫害的报道甚少,以母生蛱蝶、杨扇舟蛾、慧双尾舟蛾和母生小木虱为主[1-2]。短梳角野螟于1859年在印度首次被Walker发现,刘元福[3]在对海南海峡尖峰岭林区螟蛾科进行的昆虫区系划分中对螟蛾科41属71种优势种进行了描述,首次采集到短梳角野螟标本。王平远[4]于1980年撰写的中国经济昆虫志(第二十一册 螟蛾科)中对短梳角野螟成虫形态进行了描述,但未涉及该虫各虫态生物学特性与虫龄划分等研究。
Esperk等指出昆虫分龄是开展生物学研究的基础[5-6]。为摸清短梳角野螟在红花天料木树种植区的成灾规律,适时防控其危害,作者通过林间采样与室内饲养相结合,对海南省澄迈地区红花天料木种植区短梳角野螟的形态特征、生物学特性及生活史进行了系统调查,并结合数据分析,明确各龄幼虫形态指标,划分了幼虫龄期,为进一步开展短梳角野螟生态学及林间防控研究奠定基础。
1 材料与方法
1.1 试验材料
各龄短梳角野螟幼虫在海南省澄迈县国营林场红花天料木人工林(110°32′51″ ~ 110°34′54″E, 21°43′42″ ~ 21°44′09″N)采集并分装在相同规格的养虫瓶中,置于室内温度为(28.0±1.0)℃、相对湿度75.0%±5.0%、光照L∥D=14 h∥10 h条件下饲养,羽化后以10%的蜂蜜水作为成虫的营养来源。1龄幼虫由卵孵化人工饲养。
1.2 研究方法
1.2.1 短梳角野螟生物学特性调查
2014年11月到2016年5月,在虫口密度较大的红花天料木林地内设置固定调查样地,随机选取 10株有虫红花天料木植株,每个植株按东南西北4个方位标记 4 个有虫枝条,定期在样树上调查短梳角野螟的发生情况。每间隔10 d调查1次,记录短梳角野螟产卵的位置及卵的形态、颜色、数量等;观察并记录该虫的为害特征、为害部位、虫态及生活习性等; 并通过佳多频振式杀虫灯(PS-15II DC 12V,佳多科工贸有限责任公司)调查成虫的发生情况,方法参照杨美红等[7]和周云娥等[8]。
1.2.2 短梳角野螟幼虫分龄指标及测量
本试验采用实验室饲养及林间收集的方法得到足够多的供试幼虫,于每天中午12:00选取30头幼虫进行拍摄测量。由于海南气候问题及短梳角野螟本身蜕皮不明显等问题,本文依据戴氏法则,即昆虫幼虫的体长增长是连续的,但相邻龄期幼虫的骨化程度较高部位却以一定的几何级数增长,如鳞翅目幼虫的头壳,并参照前人对昆虫龄期划分指标选择结果,最后选取(1)头宽(头壳最大宽度);(2)头长(头壳最大长度)(3)体长(从口器至臀部末端之间的距离)(图1)作为分龄指标。短梳角野螟龄期测定参照Dyar[9]方法。各指标通过使用尼康D90数码相机在标准测量尺下拍照测量。

图1 短梳角野螟幼虫分龄3个测量指标Fig.1 The three variables measured for separating instars of Meroctena tullalis larvae
1.3 短梳角野螟分龄数据分析
数据分析采用SPSS19.0和Microsoft Excel 2013等软件处理。参照文献[10]的方法对短梳角野螟幼虫的头宽、头长和体长进行频次统计、作图,并根据结果计算头宽、头长和体长的均值、标准偏差,根据戴氏法则和Crosby生长法则,应用Brooks指数、Crosby指数以及线性回归来确定短梳角野螟最佳龄期划分[9,11]。
计算公式如下:
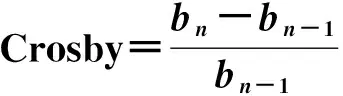
式中:Xn和Xn-1分别指第n龄幼虫和第n-1龄幼虫各自头壳宽度的均值;bn和bn-1分别指第n和n-1个Brooks指数。
2 结果与分析
2.1 短梳角野螟的形态特征
2.1.1 卵块
扁圆形,直径0.61~0.67mm,初产卵肉色,卵表面有淡黄色网纹将其覆盖,卵块一般包含十几粒至四五十粒卵,孵化前变为淡黄色,孵化后的幼虫取食叶背叶肉(图2a)。
2.1.2 幼虫
体长3~30mm,头呈半球形,1~2龄头部肉色,3龄起头部外围开始出现一圈红褐色;胸部3节,有3对胸足,胸足浅黄色,跗节末端有爪;前胸气门呈圆形,前侧毛2根;腹部10节,有4对腹足(位于第3~6腹节)和1对臀足(位于第10腹节),足为乳白色,趾钩3行,呈缺环排列;老熟幼虫身体收缩变粗,体长一般为8~12mm,体色转变黄绿色,头部外围褐色与内部肉色相差更明显(图2b~f)。
2.1.3 蛹
锥形,头端稍大,尾端尖削,长10.31~14.85 mm,初化蛹为淡黄绿色。腹部10节,腹面淡黄色,翅伸至第6腹节1/4处。臀棘圆锥形,末端带有小钩。蛹接近羽化时复眼呈黑色,圆形,翅型轮廓明显,呈茶褐色(图2g~h)。
2.1.4 成虫
成虫翅展29~34 mm。鲜黄色,下唇须褐色,基部白色,前翅有不明显的褐色亚基线,内横线褐色倾斜,中室内有一小黑点,中室端脉斑暗褐色新月形,中线褐色从Cu2脉伸向翅内缘,外横线褐色自翅前缘伸到M2脉,亚缘线褐色弯曲波纹状,外侧淡红褐色。后翅有一中室端脉褐斑,外横线褐色不明显,但在M2至Cu2脉之间有曲线,Cu2脉以下模糊不清,向翅臀角有淡红色斑及一暗色带。双翅缘毛基部各有一条黑线,缘毛白及黑色相间隔(图2i)。
2.2 短梳角野螟生活史
短梳角野螟在海南省澄迈地区一年发生8代(表1),每年4月到 12 月为该虫的为害期,有世代交替现象,完成一个世代历期1个月左右。卵期3~4 d;幼虫期9~13 d,蛹期6~7 d,成虫寿命6~8 d。老熟幼虫以蛹越冬,翌年 3月越冬蛹开始陆续羽化为成虫,4 月幼虫开始为害,至 12 月随着嫩叶的减少和温度的降低,短梳角野螟开始化蛹越冬。

表1 短梳角野螟生活史(海南澄迈)1)
1) ·:卵;-:幼虫;⊕:蛹;+:成虫;(⊕):越冬蛹; F:上旬; M:中旬; L:下旬。 ·:Egg;-:Larva;⊕:Pupa;+:Adult;(⊕):Over-wintering pupa; F:The first ten days; M:The middle ten days; L:The last ten days.
2.3 短梳角野螟生活习性
成虫全天均可羽化,以晚上居多,具趋光性。日间主要在红花天料木中上层叶背面或枝叶间隐蔽处静止,成虫羽化1 d后即可开始交配产卵,产卵集中在叶片背面,每头雌蛾一生产卵百粒以上。初孵幼虫附于叶片背面,2~3 d后开始卷叶为害,卷叶时头部左右摇摆并吐丝,借丝的拉力,将叶缘部分向里弯曲,形成卷叶,喜食嫩叶,造成叶面缺刻。老熟幼虫在卷叶中结茧化蛹。
2.4 短梳角野螟幼虫龄期的判断
本试验测定了552头短梳角野螟幼虫的头宽和体长,分别对短梳角野螟幼虫三项指标测定值进行频次分布统计分析,绘制频次分布表和频次分布图。结果显示:各测量指标均具有5个明显的峰值见图3。根据频次分布一个频次分布集中区域对应一个幼虫龄期的原则[12],可将短梳角野螟幼虫划分为5龄。由图3峰值分布可得,幼虫头宽的峰值分布明显优于头长和体长,可初步判定头宽为划分短梳角野螟的最佳指标。
根据测量值分别计算短梳角野螟各龄幼虫3项指标的平均值、变幅、标准差、变异系数、Brooks指数和Crosby指数,结果见表2。分析表2的统计结果表明,各项分龄指标变异系数均小于20%,Crosby指数均小于10%,表明将短梳角野螟幼虫划分为5个龄期是准确的,戴氏法则进一步验证了其合理性(图4)。

表2 短梳角野螟3项分龄指标测量值结果1)
1) Y1:头长; Y2:头宽; Y3:体长; 表中同一指标均值后不同字母表示经LSD多重比较在0.05水平上差异显著。 Y1:Head capsule length;Y2:Head capsule width;Y3: Body length;The lowercase letters in the same column indicate significant difference at 0.05 level.

图3 短梳角野螟头长(a)、头宽(b)和体长(c)频率分布图Fig.3 Frequency distribution of the head capsule length (a), head capsule width (b) and body length(c) in Meroctena tullalis
陈永年等[13]指出昆虫幼虫骨化结构测量值的对数值与幼虫龄期之间呈直线或修指指数曲线关系,邓侨等[14]在骆驼斯氏副柔线虫病传播媒介西方角蝇Haematobiairritans和截脉角蝇H.titillans的幼虫龄期划分中也验证了骨化结构测量值的对数值与幼虫龄期之间存在上述关系。利用SPSS 19.0对短梳角野螟幼虫3项分龄指标对数值与幼虫龄期关系进行拟合(表3、图4)。由结果可见短梳角野螟的三项指标的线性关系、指数关系及增长关系与幼虫龄期存在极显著相关关系;其中短梳角野螟3项分龄指标与龄期之间的三次线性拟合模型回归系数最大,二次线性拟合模型回归系数次之,指数与增长再次之;由表3还可以看出,幼虫头宽测量值的对数值与幼虫龄期的相关系数要优于幼虫头长和体长测量值的对数值与幼虫龄期的相关系数,即头宽可作为划分短梳角野螟的最佳指标。由图4可以看出,短梳角野螟各测量值的平均值自然对数与幼虫龄期线性相关极为显著(P<0.001),且头宽与龄期的回归系数最大(R2=0.9991),这进一步验证了头宽可作为划分短梳角野螟幼虫龄期的最佳指标,因此幼虫头长和体长可作为辅助指标,验证标准。
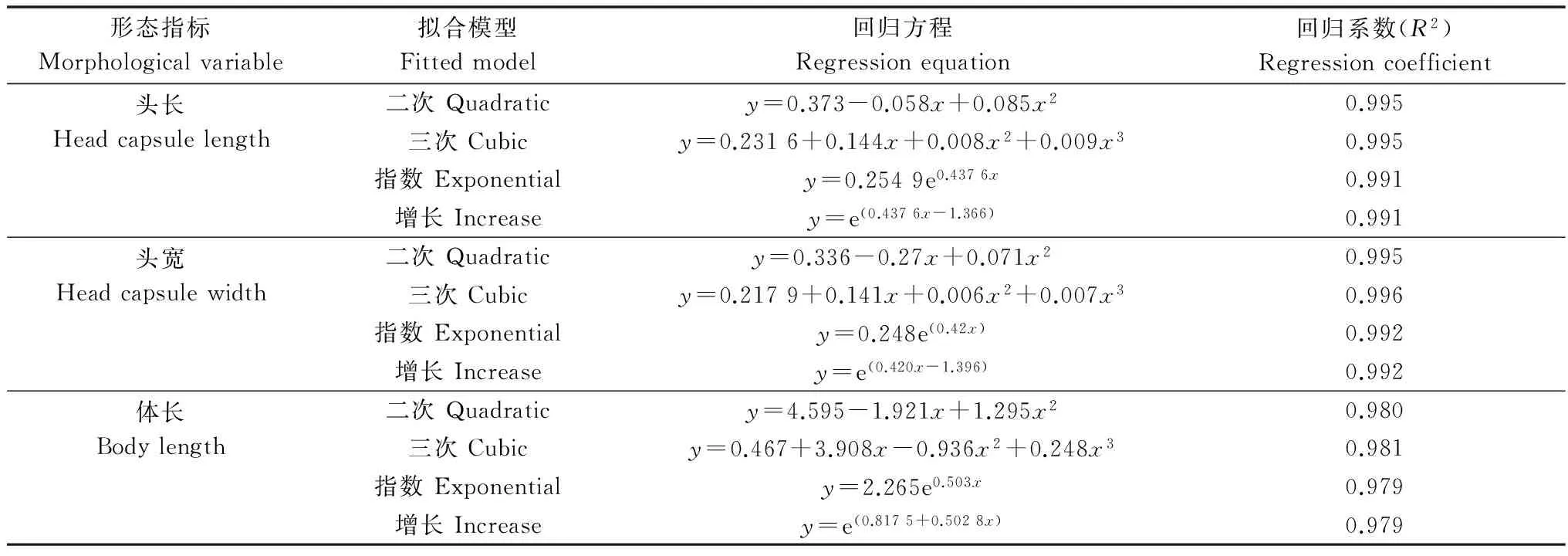
表3 短梳角野螟幼虫分龄指标与幼虫龄期的拟合结果表

图4 短梳角野螟幼虫头宽、头长及体长平均值的自然对数与虫龄的回归关系Fig.4 Regression relationships between the natural logarithm of the means of the head capsule width, head capsule length and body length of all instars of Meroctena tullalis larvae and the instar numbers
3 结论与讨论
划分幼虫龄期常用方法是对骨化结构进行测量,Dyar指出,昆虫骨化部位随周期性蜕皮而呈现跳跃式增长,故常用此骨化部位作为分龄指标的一个重要依据。董易之等、赖开平等[15-16]指出,昆虫幼虫虫龄与幼虫头宽存在一定的几何级数关系,故常用头宽作为昆虫虫龄划分的重要指标。本研究结果表明,短梳角野螟的头宽是划分幼虫龄期的最佳指标,头长和体长是分龄的辅助和验证指标。在已有研究中,李志文等[17]研究表明,头壳宽可作为油茶象CurculiochinensisChevrolat虫龄划分的可靠依据;贺春玲等[18]通过测量幼虫头壳宽度初步判定了长木蜂Xylocopatranquebarorum幼虫为4龄;魏书军[19]等发现以口片最大宽度作为柑橘爆皮虫AgrilusauriventrisSaunders分龄指标,可准确有效地将柑橘爆皮虫幼虫分为5龄;Panzavolta[20]通过头宽可有效划分栗木蠹象Pissodescastaneus的龄期。本研究以此为参考选取类似指标。陈永年等[13]指出昆虫幼虫头宽对数值与虫龄呈直线相关关系,杨宗慧等[21]指出椰子织蛾Opisinaarenosella头壳宽度与龄数呈显著指数相关关系;邓侨等[14]在对斯氏副柔线虫Parabronemaskrjabini龄期划分中得出二次和三次模型结果最好,周斌等[22]在对降香黄檀棕斑澳黄毒蛾OrvascasubnotataWalker龄期划分中得出复合曲线、增长曲线和指数模型拟合效果最好。本试验通过对短梳角野螟幼虫3项分龄指标对数值与幼虫龄期关系进行拟合,确定三次线性拟合模型结果最好;并对分龄指标平均值的自然对数值与幼虫龄期进行回归分析,得出头宽平均值的自然对数值与龄数呈线性关系,R2=0.9991,达到显著相关水平。本试验结果与已有相关报道一致。
综上所述,通过测量短梳角野螟头宽,利用本试验所得三次线性拟合方程和测量平均值的自然对数与龄期的相关系数,可准确、简便、快速地划分短梳角野螟幼虫龄期,为短梳角野螟林间防治奠定了基础。
短梳角野螟生物学特性及生活史的探究作为一项基础研究,不仅能丰富理论知识,也为鉴定红花天料木螟蛾虫害提供理论依据。关于短梳角野螟的生物学特性,王平远[4]先生曾记述其幼虫有钻蛀茎干在内部取食的习性,而本研究发现短梳角野螟卷叶为害,造成这种差异的原因还有待于更为广泛而深入的调查研究。
温度、湿度、光周期、食物量、风、饲养密度等都能影响昆虫的生理、生活史和行为等生物学特性[23]。有关环境因子对短梳角野螟生物学特性的影响,还需进一步研究。
目前,针对短梳角野螟的防治主要是化学农药,由于短梳角野螟生活隐蔽,主要以幼虫卷叶为害为主,喷洒一定浓度的农药难以取得相应的成效,因此利用自然天敌与研制微生物农药对短梳角野螟林间防治迫在眉睫。作者在试验过程中发现,短梳角野螟不仅存在寄生性天敌,并且在林间发现有相当一部分幼虫因感染病原菌而死亡。因此,对天敌的研究和分离病原菌进行微生物农药研制是对短梳角野螟进行林间防治的重要部分,需要进一步研究和完善。
[1] 顾茂彬, 陈佩珍. 母生蛱蝶的初步研究[J]. 林业科学, 1987,23(1): 105-108.
[2] 陈泽坦. 珐蛱蝶——母生树上的一种新害虫[J]. 植物保护, 2002, 28(5): 59-60.
[3] 刘元福. 海南岛尖峰岭林区昆虫区系——螟蛾科[J]. 林业科学研究, 1990, 3(6): 574-579.
[4] 王平远. 中国经济昆虫志 第二十一册 鳞翅目 螟蛾科[M]. 北京:科学出版社, 1980.
[5] Esperk T, Tammaru t, Nylin S. Intraspecific variability in number of larval instars in insects [J]. Journal of Economic Entonmology, 2007,100(3):627-645.
[6] Chen Y, Seybold S J. Application of a frequency distribution method for determining instars of the beet armyworm (Lepidoptera: Noctuidae) from widths of cast head capsules[J]. Journal of Economic Entonmology, 2013, 106(2):800-806.
[7] 杨美红, 牛辉林, 张金桐, 等. 榆木蠹蛾生物学特性观察[J]. 应用昆虫学报, 2012, 49(3): 735-741.
[8] 周云娥, 黄琼瑶, 白洪清, 等. 竹金黄镰翅野螟生物学特性研究[J]. 昆虫学报, 2008, 51(10): 1094-1098.
[9] Dyar H G. The number of molts in lepidopterous larvae [J]. Psyche, 1890, 5: 420-422.
[10]刘小虎. SPSS 12.0 for windows在农业试验统计中的应用[M]. 沈阳:东北大学出版社, 2007:39-46.
[11]Daly H V. Insect morphometrics [J]. Annual Review of Entomology, 1985, 30(1): 415-438.
[12]Hunt G, Chapman R E. Evaluating hypotheses of instar-grouping in arthropods: a maximum likelihood approach[J]. Paleobiology, 2001, 27(3): 466-484.
[13]陈永年, 潘桐. 也谈昆虫幼虫头宽及摄食量的增长规律[J]. 昆虫知识, 1988(4): 235-240.
[14]邓侨, 杨莲茹, 杨晓野, 等. 骆驼斯氏副柔线虫病传播媒介西方角蝇和截脉角蝇的幼虫龄期划分[J]. 昆虫学报, 2015, 58(3): 341-350.
[15]董易之, 陈炳旭, 徐淑, 等. 荔枝大造桥虫幼虫龄数的划分[J]. 环境昆虫学报, 2011, 33(4): 552-556.
[16]赖开平, 陆温, 刘德星, 等. 眉斑并脊天牛幼虫龄数及各龄特征[J]. 昆虫知识, 2008, 45(1): 138-140
[17]李志文, 何立红, 夏姣, 等. 油茶象幼虫虫龄的划分[J]. 昆虫学报, 2015, 58(2): 181-189
[18]贺春玲, 曹磊, 李梦. 长木蜂幼虫龄期的划分[J]. 环境昆虫学报, 2015, 37(1): 128-132.
[19]魏书军, 郑宏海, 皇甫伟国, 等. 柑桔爆皮虫幼虫龄期的划分[J]. 昆虫学报, 2006,49(2): 302-309.
[20]Panzavolta T. Instar determination forPissodescastaneus(Coleoptera: Curculionidae) using head capsule widths and lengths[J]. Environmental Entomology, 2007,36(5):1054-1058.
[21]杨崇慧, 阎伟, 李朝绪, 等. 椰子木蛾幼虫龄期的划分[J]. 植物保护, 2014, 41(2): 70-74.
[22]周斌, 周国英, 杨权, 等. 降香黄檀食叶害虫棕斑澳黄毒蛾的生物学特性及幼虫虫龄判断[J]. 昆虫学报, 2015, 58(11): 1253-1261.
[23]张晶, 胡冰冰, 李后魂, 等. 宽瓣头细蛾形态及生物学特性研究[J]. 昆虫学报, 2012, 55(5):585-595.
(责任编辑:田 喆)
Morphological characteristics and determination ofMeroctenatullalislarval instars
Li Xiaomin, Liu Jun’ang, Ouyang Bowen, Dong Wentong, Zhou Bin, Zhou Guoying
(Hunan Provincial Key Laboratory for Control of Forest Diseases and Pests;Key Laboratory ofCultivation and Protection for Non-wood Forest Trees of Ministry of Education, CentralSouth University of Forestry and Technology, Changsha 410004, China)
Meroctenatullalisis one of the most important pests which harmHomaliumhainanenseGagnep and it is very important to determine the instars of the larva and morphological characters. Here we made a detailed study on the morphological characters, the habit of each developmental stage and the life history ofM.tullalis. Through laboratory determination combined with field investigation, the three variables were head length, head width and body length, and the data were analyzed by SPSS 19.0 combined with the Dyar’s rule and linear regression to design a reliable method for determining the instars of an individual larva. The results indicated that, as the larvae grew, the body color gradually changed from light green to deep green, and the head color progressively deepened and finally became redish-brown and deep color in the middle. Statistical analysis of the body measurements showed that the head capsule width was the most important indicator of larval instars, and the natural logarithm of the means of the head capsule width was significantly correlated with the instar number (y=0.181 6x-0.602 3,R2=0.999 1). The head length and body length distribution fitted the Dyar’s law. However, due to the lack of obvious peaks, they could only used as the auxiliary indices for determining larval instars. This study described the biological characteristics of this insect and identified the best way for determination of larval instars, which may provide crucial information for developing control and prevention strategies.
Meroctenatullalis; morphological characteristics; larval instar; life history;Homalicumhainanense
2016-06-01
2016-07-11
国家林业公益性行业科研专项(201304402)
S 433.4
A
10.3969/j.issn.0529-1542.2017.03.023
致 谢: 感谢中南林业科技大学魏美才和黄建华教授对短梳角野螟的鉴定及澄迈林场工作人员对林间工作的支持和帮助。
* 通信作者 E-mail: zgyingqq@163.com
猜你喜欢
数学物理学报(2022年5期)2022-10-09
南通职业大学学报(2021年3期)2021-11-17
海河水利(2021年4期)2021-08-30
信阳农林学院学报(2021年1期)2021-04-01
林业科技(2020年3期)2021-01-21
矿业科学学报(2020年5期)2020-09-18
债券(2020年4期)2020-08-04
浙江建筑(2019年6期)2019-12-26
计算机应用(2018年12期)2019-01-08
商周刊(2018年26期)2018-12-29
