
黄土高原典型草原优势植物凋落物分解及养分释放对氮添加的响应
2017-05-15 12:36:17文海燕
生态学报 2017年6期
文海燕, 傅 华, 郭 丁
兰州大学草地农业科技学院,草地农业生态系统国家重点实验室, 兰州 730020
黄土高原典型草原优势植物凋落物分解及养分释放对氮添加的响应
文海燕*, 傅 华, 郭 丁
兰州大学草地农业科技学院,草地农业生态系统国家重点实验室, 兰州 730020
利用原位分解袋法研究了黄土高原典型草原优势植物长芒草(Stipabungeana)和阿尔泰狗娃花(Heteropappusaltaicus)凋落物的养分释放过程对氮添加的响应,试验周期为1 a。设置6个氮添加水平,分别为N0(0)、N1(1.15 g N m-2a-1)、N2(2.3 g N m-2a-1)、N3(4.6 g N m-2a-1)、N4(9.2 g N m-2a-1)和N5(13.8 g N m-2a-1),氮素类型为尿素((NH2)2CO)。结果表明:(1)氮添加处理两年显著改变了长芒草和阿尔泰狗娃花凋落物的初始化学性质。随着氮梯度的增加,凋落物的N(氮)含量逐渐增加,木质素含量先增加后下降,C/N(碳氮比)和木质素/N降低,C(碳)、P(磷)和C/P(碳磷比)没有显著的差异。(2)氮处理对长芒草和阿尔泰狗娃花凋落物的分解速率的影响不显著。长芒草和阿尔泰狗娃花凋落物C含量随分解时间整体为降低过程,N和P含量总体上为增加过程,且整个分解过程中N含量各处理间差异显著。(3)氮处理对长芒草和阿尔泰狗娃花凋落物C和P的分解基本无影响,两种元素都呈现释放过程。氮处理对凋落物的N残留率有显著的影响,在N1—N3(1.15—4.6 g/m2)处理下的长芒草凋落物N残留率高于其他处理,且呈现富集过程;而阿尔泰狗娃花凋落物中的N呈现富集-释放过程。在土壤养分贫瘠的黄土高原典型草原,适量的氮输入可以促进系统的固氮。
氮添加;草原;凋落物;养分释放;黄土高原
工业革命以来由于化石燃料的燃烧和化肥的施用,使生态系统中氮的输入量增加了近3到5倍,且呈持续上升的趋势[1]。大多数生态系统均受氮素限制,所以氮沉降会直接影响生物圈的氮循环,并且通过氮输入的增加影响植物生产力和生态系统碳蓄积能力,从而使生态系统的结构和功能发生改变[2]。
凋落物是分解者亚系统的重要组成部分,将生产者和消费者两个环节联结起来。凋落物动态影响着植物萌发、群落结构和演替,在改善生态环境及土壤理化性状、能量流动和营养循环过程中起着重要作用[3]。近年来,国内外许多学者对模拟氮沉降条件下的凋落物的分解过程进行了研究,发现氮沉降增加对植物凋落物分解的影响并不一致[4- 6]。植物凋落物的分解是形成土壤有机质的初期过程,影响着土壤的呼吸过程且凋落物分解所释放的各种养分对植物的生长具有十分关键的调控作用。因此氮沉降增加对凋落物分解过程的影响,将在一定程度上影响着土壤肥力及生态系统初级生产力。
草原生态系统是陆地生态系统中最重要、分布最广的生态系统类型之一,由于草原生态系统的脆弱性及其对气候变化响应的敏感性,决定了它在陆地生态系统的元素和能量循环研究中的独特地位及重要性。然而,由于氮沉降问题的提出以及后续科学研究主要集中于欧洲和北美,因此氮沉降研究主要关注森林生态系统;对氮沉降的增加如何影响草原生态系统凋落物的分解过程仍不十分清楚。因此,本研究以在黄土高原典型草原已实施两年模拟氮沉降的样地为研究对象,研究了氮添加梯度下两种优势植物长芒草(Stipabungeana)和阿尔泰狗娃花(Heteropappusaltaicus)凋落物的分解速率和元素残留率的变化规律,以期阐明氮沉降对黄土高原典型草原凋落物分解过程的影响,为该区草原生态系统的管理和氮沉降的深入研究提供基础资料。
1 材料与方法
1.1 试验地概况
研究地点位于甘肃省榆中县兰州大学黄土高原国际地面气候与环境监测站围封草地内,地处35°57′N,104°09′E,该地区地貌为黄土高原残塬梁峁沟壑,属于大陆性半干旱气候,海拔为1965.8 m,年均降水量为382 mm,年均蒸发量1343 mm,年均气温6.7℃,年日照时数大约2 600 h,无霜期90—140 d。土壤类型为灰钙土,植被类型为黄土高原半干旱草原,主要植物有长芒草(Stipabungeana)、阿尔泰狗娃花(Heteropappusaltaicus)、赖草(Leymussecalinus)等,各物种的重要值见表1。
1.2 试验设计
选择地势平缓地形(海拔、坡向)一致的地段,选取面积为4 hm2的围封草地(2005年开始围封)。小区面积4 m×5 m,6个氮素添加处理,每处理5个重复,30个小区,完全随机区组设计。氮素梯度分别为N0(0)、N1(1.15 g N m-2a-1)、N2(2.3 g N m-2a-1)、N3(4.6 g N m-2a-1)、N4(9.2 g N m-2a-1)和N5(13.8 g N m-2a-1)。于2009年开始实施氮素梯度处理,氮肥选用尿素((NH2)2CO),施加时间为每年的6月底。为减少氮素的损失,选择在下雨时进行氮添加。添加的具体方法:将尿素溶解于5 L水中,用洒壶均匀喷洒在每个小区内,洒壶喷洒等量水在对照样地。

表1 试验地群落的物种组成及其重要值
重要值= (相对密度+相对高度+相对盖度)/3
为了保证试验材料的来源,2010年12月22日从30个处理小区内分别齐地面收集长芒草和阿尔泰狗娃花的立枯物,作为供试凋落物材料[7]。将凋落物在室温下晾至恒重后,将植物剪成5 cm长的片段,准确称取2 g,分别装进10 cm×10 cm的尼龙网袋中(网眼大小为1 mm2)。每个小区每种凋落物制成4个分解袋,为试验准备分解袋240个。
2011年1月12日,将各氮处理生产的凋落物放在各原处理的小区中用竹签固定;分别于2011年的4月12日(已分解3个月)、7月12日(已分解6个月)、10月12日(已分解9个月)和2012年1月12日(已分解12个月),分4次每个小区每种植物取一袋。在实验室中,用手捡出混进分解网袋中的其它植物的材料、小石头和钻进网袋的小动物等。从分解网袋中取出凋落物后,将其装在信封中放于烘箱中在65 ℃下烘48 h称重,由此得到分解一段时间后的残留重量。称重后将凋落物粉碎用于分析其成分含量。
1.3 样品分析
碳(C)、氮(N)采用元素分析仪(FlashEA1112, USA),磷(P)采用微波消解-钼蓝比色法,木质素含量采用Detergent法[8]。
1.4 数据分析
(1) 凋落物干物质残留率(%)
干物质残留率=Mt/M0× 100%
式中,M0为凋落物的初始干重(g),Mt是t时刻凋落物分解袋中剩余样品的干重(g)。
(2) 凋落物分解速率(k)
按照Olson指数衰减模型[9]模拟凋落物分解过程,计算凋落物分解系数(k):
Mt/M0=e-1kt
式中,k为凋落物分解系数,t为分解时间。
(3) 元素残留率(R)
R=(Ct×Mt)/(C0×M0)× 100%
式中,Ct为t时刻凋落物元素含量(g/kg),C0为初始元素含量(g/kg)。
数据采用SPSS 16.0软件进行统计分析,选用单因素方差分析(ANOVA)判断氮添加处理间各凋落物各性状的差异,LSD法进行多重比较。采用Excel制图。
2 结果与分析
2.1 凋落物的初始化学性质(干基)
氮添加处理2年后,长芒草和阿尔泰狗娃花凋落物的初始化学性质有明显的差异(表2)。随着氮梯度的增加,长芒草和阿尔泰狗娃花凋落物的N含量逐渐增加,木质素含量呈现先增加后下降的趋势,C、P和C/P(碳磷比)没有显著的差异,C/N(碳氮比)和木质素/N降低。长芒草凋落物的N、P和木质素含量低于阿尔泰狗娃花凋落物。

表2 凋落物的初始化学性质(干基)
小写字母表示氮处理间的比较,字母不同者表示差异显著(P<0.05)
2.2 氮添加对凋落物干物质残留率和分解速率的影响
随着分解时间延长,长芒草和阿尔泰狗娃花凋落物的干物质残留率均呈逐渐下降趋势(图1)。分解12个月后,两种凋落物的干物质残留率没有显著的差异。

图1 氮处理对凋落物干物质残留率的影响(*P<0.05)Fig.1 Percentages of mass remaining in the different nitrogen (N) treatments for the two litters (*P<0.05)

处理Treatments年分解常数kAnnualdecompositionrate/a-1长芒草Stipabungeana阿尔泰狗娃花HeteropappusaltaicusN00.31±0.05a0.59±0.09aN10.32±0.04a0.60±0.04aN20.33±0.07a0.68±0.03aN30.32±0.03a0.57±0.07aN40.39±0.04a0.60±0.07aN50.37±0.05a0.57±0.06a
分解常数k值的生态学意义是k值越大,凋落物的分解速率越快[10]。从表3可以看出,阿尔泰狗娃花凋落物的年分解速率均快于长芒草凋落物的年分解速率,它们的分解过程都可以很好的用Olson的指数方程进行模拟;长芒草和阿尔泰狗娃花k值的范围分别为0.31—0.38和0.57—0.68,各处理间两种植物凋落物都无显著的差异。
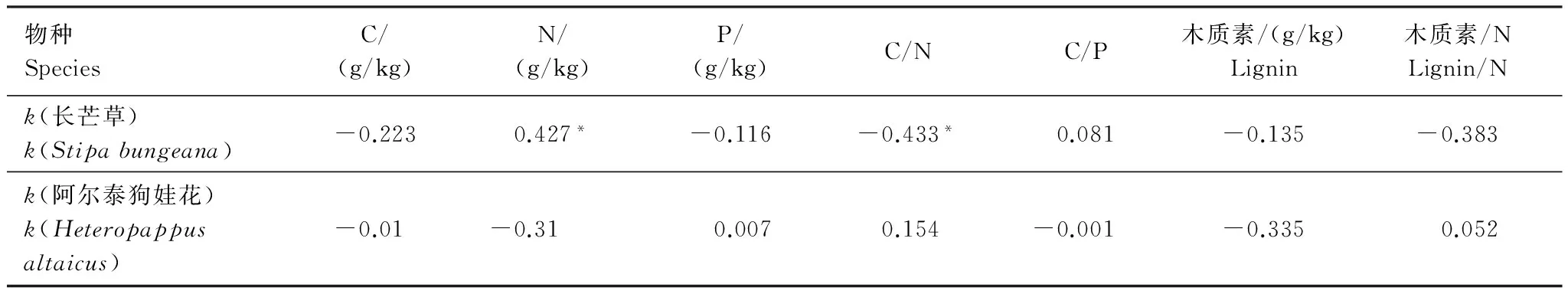
表4 凋落物年分解常数(k)与其初始化学性质(干基)的相关性
从表4可以看出,长芒草凋落物的分解速率与其初始N含量呈显著正相关关系(P<0.05),与C/N呈显著负相关关系(P<0.05);阿尔泰狗娃花凋落物的分解速率与其初始化学组成没有显著的相关关系。
2.3 氮添加对凋落物分解过程C、N、P含量的影响
从图2可以看出,长芒草凋落物的C含量呈现分解前3个月增加,3—9个月急剧减少,9—12个月略有增加的趋势;N含量在整个分解过程中总体呈现上升的趋势,且分解过程中各处理间都有显著的差异(P<0.05);P含量在0—3月各处理都降低,3—9月增加,9—12月各处理变化不一致。

图2 氮处理对长芒草分解过程C、N、P含量的影响Fig.2 Effects of nitrogen (N) treatments on carbon (C), N and phosphorus (P) contents of Stipa bungeana
从图3可知,阿尔泰狗娃花凋落物的C含量呈现上升-下降-上升的趋势;N含量0—3月各处理都增加,3—6月减少,6—12月各处理变化不一致。阿尔泰狗娃花P含量呈现“M”形状的变化规律,各处理只在分解9月和12月有显著的差异(P<0.05)。综合图2和图3可以发现,总体来看长芒草和阿尔泰狗娃花的C含量呈下降的趋势,N和P含量呈现增加的趋势。

图3 氮处理对阿尔泰狗娃花分解过程C、N、P含量的影响Fig.3 Effects of nitrogen (N) treatments on carbon (C), N and phosphorus (P) contents of Heteropappus altaicus
2.4 氮添加对凋落物分解过程C、N、P元素残留率的影响
由表5可知,凋落物分解12个月后,氮处理对长芒草凋落物C和P残留率基本无显著影响;氮处理对长芒草凋落物的N残留率有显著的影响(P<0.05)。分解1年后,在N1—N3处理下的凋落物N残留率高于其他处理,且呈现富集过程。

表5 氮处理对长芒草分解过程元素残留率的影响
从表6可以看出,氮处理对阿尔泰狗娃花凋落物分解过程中C和P残留率基本无显著影响,分解3个月和6个月时N3—N5处理凋落物N残留率显著高于对照(P<0.05),且3个月时呈富集过程,分解后期各处理间无显著差异。

表6 氮处理对阿尔泰狗娃花分解过程元素残留率的影响
3 讨论
3.1 凋落物的初始化学组成对氮添加的响应
凋落物的初始化学组成对氮添加有着不同的响应,挪威云杉叶片凋落物的N含量与N的施用量成正比,凋落物的N含量从对照的4.2 mg/g加到高N处理的18.3 mg/g,木质素含量从300 mg/g增加到407 mg/g[11]。但是对两个土壤养分状况有差异的草地进行了12 年的施氮肥试验,发现施氮肥几乎对凋落物的化学组成没有影响[12]。也有研究发现施氮肥6个月后,3 种高草草原植物叶片凋落物的木质素含量稍微下降[13]。本研究结果表明,随着氮梯度的递增,长芒草和阿尔泰狗娃花的初始N含量都显著增加(P<0.05),C和P含量没有显著的影响(表2)。而两种凋落物的木质素含量随着氮梯度的增加呈现先增加后下降的趋势(表2),有研究表明氮素能够影响木质素合成关键酶的活性,适量的氮会增加小麦和水稻茎的木质素含量,而过多的氮则降低了茎秆木质素合成相关酶的活性和木质素含量[14- 16]。
3.2 氮处理对凋落物分解过程的影响
影响凋落物分解的主要因素有内在和外在因素,内在因素是指凋落物自身的物理和化学性质;外在因素是指凋落物分解过程发生的外部环境,包括参与分解的异养微生物、土壤动物、土壤理化性质、水热条件等外界环境[17]。一般认为,氮沉降改变了凋落物的化学组成成分,进而影响凋落物的分解速率[18]。而本研究发现氮处理虽然增加了凋落物的初始N含量,改变了凋落物的C/N和木质素含量(表2),但是氮处理下只有长芒草凋落物的初始N含量,C/N与其分解速率有显著的相关关系(P<0.05),阿尔泰狗娃花的初始化学组成和其分解速率并无显著的相关关系(表4),因此研究结果表明凋落物的分解速率与其初始化学组成之间的相关关系因研究的凋落物类型的不同而存在差异,并且氮处理引起的外在因素的改变可能会改变凋落物的分解规律。
通过对黄土高原典型草原进行为期一年的模拟氮沉降试验,研究结果表明,凋落物的C含量总体呈现降低的趋势,但是氮处理对分解一年后的两种凋落物的C含量和释放过程均没有显著的影响。在一些森林、湿地和草地生态系统中发现,氮添加促进了微生物的活性[19];而其它研究发现抑制了微生物的活性[20],或者影响不明显[21]。这些结果很不一致,其部分原因可能是由于微生物群落的组成差异以及酶系统对氮增加的响应差异所引起的。Waldrop等[22]认为,氮添加对含顽拗凋落物(Recalcitrant Litter)较多、以白腐真菌为主要分解者的生态系统的影响是逆向的,而对含有易分解凋落物、维持不同真菌种群(如软腐真菌)的生态系统的影响是正向的。因此,氮处理对凋落物分解过程中C含量的影响是非常复杂的,既决定于生态系统分解者的种类和活性,也受到气候、植物种及环境条件等多种因素的影响。
尽管在试验中,氮处理对长芒草凋落物的年分解速率没有显著的影响(表3),但是N1—N3处理(1.15—4.6 g/m2)却显著增加了N素的固定(表5)。有研究者认为积累在凋落物中的N是一种“过度摄取”,并不是所有固定的N都是分解者微生物所必需的。在富含N的底物降解的过程中,每释放出一单位的CO2,微生物都需要固定更多的N,所以在分解过程中会一直表现出对N的积累,直至易分解的C被完全分解[23]。微生物所固定的N,其中一部分会被单宁沉淀,固定在木质素或酸-不溶性物质中,而不是被固定在微生物量中[24]。尽管养分的固持暂时打破了分解过程中养分的矿化,却为低养分或退化生态系统中养分的固定提供了一个重要的机制[25- 26]。长芒草是黄土高原典型草原的优势种,在群落中的重要值为0.401(表1),其生物量占到总生物量的50%以上[27],所以在该生态系统低浓度的氮沉降增加可以增加系统的固N量。
4 结论
1a的凋落物分解试验表明,氮添加对长芒草和阿尔泰狗娃花凋落物的分解速率无显著的影响。两种凋落物的C含量随分解时间整体为降低过程,N含量都表现为增加趋势,且整个分解过程中各处理间差异显著,P含量总体上为增加过程。氮添加对两种凋落物C和P的分解过程基本无影响,两种元素呈现释放过程。但是氮处理对长芒草凋落物的N残留率有显著的影响,经过1a的分解,N1—N3处理下的长芒草凋落物中N残留率高于其他处理,且呈现富集过程。因此,在土壤养分贫瘠的黄土高原典型草原,适量的氮输入可以促进系统的固氮。氮沉降对草原生态系统的影响是一个长期的过程,本试验仅仅是对1a的数据进行分析,初步揭示了凋落物的分解过程对氮沉降的响应。因此,要确切了解氮沉降对黄土高原典型草原凋落物分解的影响还需长期的定位监测。
[1] Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, FreneyJ R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science,2008, 320(5878): 889- 892.
[2] 吕超群, 田汉勤, 黄耀. 陆地生态系统氮沉降增加的生态效应. 植物生态学报,2007, 31(2):205- 218.
[3] 蒋有绪. 川西亚高山冷杉林枯枝落叶层的群落学作用. 植物生态学与地植物丛刊,1981, 5(2): 89- 98.
[4] 莫江明, 薛璟花, 方运霆. 鼎湖山主要森林植物凋落物分解及其对N沉降的响应. 生态学报,2004, 24(7): 1413- 1420.
[5] Tu L H, Hu H L, Hu T X, Zhang J, Liu L, Li R H, Dai H Z, Luo S H. Decomposition of different litter fractions in a subtropical bamboo ecosystem as affected by experimental nitrogen deposition. Pedosphere, 2011, 21(6): 685- 695.
[6] Bragazza L, Buttler A, Habermacher J, Brancaleoni L, Gerdol R, Fritze H, Hanajík P, Laiho R, Johnson D. High nitrogen deposition alters the decomposition of bog plant litter and reduces carbon accumulation. Global Change Biology, 2012, 18(3): 1163- 1172.
[7] Olofsson J,Oksanen L. Role of litter decomposition for the increased primary production in areas heavily grazed by reindeer: a litterbag experiment. Oikos,2002, 96(3): 507- 515.
[8] Goering H K, Van Soest P J. Forage Fiber Analyses. Washington:U.S. DeptAgric Handbook,1970:379- 379.
[9] Olson J S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology,1963, 44(2): 322- 331.
[10] 窦荣鹏. 亚热带9种主要森林植物凋落物的分解及碳循环对全球变暖的响应[D]. 杭州: 浙江农林大学,2010.
[11] Berg B, Matzner E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environmental Reviews,1997, 5(1): 1- 25.
[12] Aerts R, DeCaluwe H, Beltman B. Plant community mediated vs. nutritional controls on litter decomposition rates in grassland. Ecology,2003, 84(12): 3198- 3208.
[13] Kemp P R, Waldecker D G, Owensby C E, Reynolds J F, Virginia R A. Effects of elevated CO2and nitrogen fertilization pretreatments on decomposition on tallgrass prairie leaf litter. Plant and Soil,1994, 165(1): 115- 127.
[14] 魏凤珍, 李金才, 王成雨, 屈会娟, 沈学善. 氮肥运筹模式对小麦茎秆抗倒性能的影响. 作物学报,2008, 34(6): 1080- 1085.
[15] 杨世民, 谢力, 郑顺林, 李静,袁继超. 氮肥水平和栽插密度对杂交稻茎秆理化特性与抗倒伏性的影响. 作物学报,2009, 35(1): 93- 103.
[16] 陈晓光, 石玉华, 王成雨, 尹燕枰,宁堂原,史春余,李勇,王振林. 氮肥和多效唑对小麦茎秆木质素合成的影响及其与抗倒伏性的关系. 中国农业科学,2011, 44(17): 3529- 3536.
[17] Moretto A S, Distel R A, Didoné N G. Decomposition and nutrient dynamic of leaf litter and roots from palatable and unpalatable grasses in a semi-arid grassland. Applied Soil Ecology,2001, 18(1): 31- 37.
[18] 马川, 董少锋, 莫江明. 鼎湖山马尾松林凋落物分解对凋落物输入变化的响应. 生态环境学报, 2012, 21(4): 647- 653.
[19] Neff J C, Townsend A R, Gleixner G, Lehman S J, Turnbull J, Bowman W D. Variable effects of nitrogen additions on the stability and turnover of soil carbon.Nature,2002, 419(6910): 915- 917.
[20] Carreiro M M, Sinsabaugh R L, Repert D A, Parkhurst D F. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition. Ecology,2000, 81(9): 2359- 2365.
[21] Hobbie S E,Vitousek P M. Nutrient limitation of decomposition in Hawaiian forests. Ecology,2000, 81(7): 1867- 1877.
[22] Waldrop M P, Zak D R, Sinsabaugh R L, Gallo M, Lauber C. Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecological Applications,2004, 14(4): 1172- 177.
[23] Fog K. The effect of added nitrogen on the rate of decomposition of organic matter.Biological Reviews,1988, 63(3): 433- 462.
[24] Gallardo A, Merino J. Nitrogen immobilization in leaf litter at two Mediterranean ecosystems of SW Spain. Biogeochemistry,1992, 15(3): 213- 228.
[25] McGroddy M E, Silver W L, De Oliveira Jr R C. The effect of phosphorus availability on decomposition dynamics in a seasonal lowland Amazonian forest.Ecosystems,2004, 7(2): 172- 179.
[26] Liu P, Huang J H, Han X G, Sun O J, Zhou Z Y. Differential responses of litter decomposition to increased soil nutrients and water between two contrasting grassland plant species of Inner Mongolia, China. Applied Soil Ecology,2006, 34(2/3): 266- 275.
[27] 程积民, 贾恒义, 彭祥林. 施肥草地群落生物量结构的研究. 草业学报,1997, 6(2): 22- 27.
Influence of nitrogen addition onStipabungeanaandHeteropappusaltaicuslitter decomposition and nutrient release in a steppe located on the Loess Plateau
WEN Haiyan*, FU Hua, GUO Ding
CollegeofPastoralAgricultureScienceandTechnology,StateKeyLaboratoryofGrasslandAgro-ecosystems,LanzhouUniversity,Lanzhou730020,China
A rise in nitrogen (N) deposition can increase net primary productivity in many terrestrial ecosystems. Litter decomposition in terrestrial ecosystems is an important component of global carbon (C) budgets and is greatly affected by the increase in N deposition that has been recorded worldwide. Grassland covers 46% of the global terrestrial area and can have an important effect on global C, and nutrient cycling and balances. We hypothesized that litter decomposition and nutrient release may respond to N deposition differently based on the quality of the litter substrate. To investigate the effect of increased N deposition on nutrient release during litter decomposition, we conducted a one-year field experiment that simulated N deposition in a steppe located on the Loess Plateau. There were six N treatments included in the experiment. These were 0 g N m-2a-1, 1.15 g N m-2a-1, 2.3 g N m-2a-1, 4.6 g N m-2a-1, 9.2 g N m-2a-1, and 13.8 g N m-2a-1. After 2-years of N treatment, the N contents in theStipabungeanaandHeteropappusaltaicuslitters increased and the C/N and lignin/N ratios gradually declined. Furthermore, the lignin first increased, but then decreased. None of the N treatments had a significant effect on the decomposition rates of the two litters after one year ofin-situdecomposition. Our results indicated that there were significant correlations between the litter decomposition rates and the N content and C/N ratio forStipabungeanalitter. The C content decreased over time, but the N and P contents increased as the two litters decomposed. The N treatments significantly influenced the N contents of the two litters during decomposition. The N treatments also influenced the percentage N remaining in the two litters. Afterin-situdecomposition for one year, the percentage N remaining inStipabungeanaunder the 1.15, 2.3, and 4.6 g N m-2a-1treatments were higher than for the other treatments and this percentage rose as the process progressed. The N release pattern inHeteropappusaltaicusshowed that N accumulated during the early stages and was released during later stages. The N treatments had no significant effects on C and P decomposition, and the C and P release patterns were similar in the two litters. Therefore, increased N deposition could contribute to the accumulation of C in a grassland ecosystem.
nitrogen addition; steppe; litter; nutrient release; Loess Plateau
国家自然科学基金资助(31602001);草地农业生态系统国家重点实验室开放基金资助(SKLGAE201401);兰州大学中央高校基本科研业务费专项资金资助(lzujbky-2016-13)
2015- 11- 11;
2016- 06- 12
10.5846/stxb201511112280
*通讯作者Corresponding author.E-mail: whaiyan@lzu.edu.cn
文海燕, 傅华, 郭丁.黄土高原典型草原优势植物凋落物分解及养分释放对氮添加的响应.生态学报,2017,37(6):2014- 2022.
Wen H Y, Fu H, Guo D.Influence of nitrogen addition onStipabungeanaandHeteropappusaltaicuslitter decomposition and nutrient release in a steppe located on the Loess Plateau.Acta Ecologica Sinica,2017,37(6):2014- 2022.
猜你喜欢
情感读本·道德篇(2020年6期)2020-08-06 14:32:28
做人与处世(2020年3期)2020-03-07 05:23:55
语文世界(小学版)(2020年11期)2020-01-08 02:19:30
孩子(2019年8期)2019-08-26 05:43:32
微型小说选刊(2016年15期)2017-01-18 09:51:00
语文世界(小学版)(2016年1期)2016-02-17 20:54:59
新疆地质(2015年3期)2015-12-10 05:08:23
工友(2015年11期)2015-08-08 01:20:48
文物鉴定与鉴赏(2015年6期)2015-07-06 17:55:26
杂文选刊(2014年10期)2014-09-18 08:07:27
