
美洲商陆的入侵机制研究
2017-05-12 14:04刘辉谢晓华何平何梨
绿色科技 2017年7期
刘辉+谢晓华+何平+何梨

摘要:指出了植物的入侵机制一直是生态学上的研究热点,为了解美洲商陆的入侵机制和验证氮分配假说是否具有通用性,通过对不同林窗环境下入侵植物美洲商陆与本地植物(芒、铁芒萁、皱叶狗尾草)的光合特征及光合氮分配特征进行了研究。结果表明:①美洲商陆具有最高的叶氮含量,最大的单位质量光合速率,最大的叶绿素浓度;②美洲商陆的最大净光合速率显著高于铁芒萁与皱叶狗尾草,叶磷含量明显高于本地植物;③美洲商陆氮在光合机制中的分配低于本地C4植物,高于本地C3植物,各林窗中美洲商陆的光合速率、氮分配均相对稳定。可见,美洲商陆入侵符合氮分配假说,同时也具有较大的生态适应性,但美洲商陆的最大净光合速率与光合氮分配仍低于本地C4植物。
关键词:美洲商陆;入侵机制;光合特征;氮分配
中图分类号:Q945
文献标识码:A 文章编号:1674-9944(2017)07-0001-06
1 引言
入侵是指外来植物在本地成功定居、繁殖,甚至取代本地植物成为优势种群的一类现象。入侵植物所带来的危害受到极大的重视[1];入侵植物如何进行成功的入侵和它们的入侵机制一直是生态学研究和争论的热点;有研究提出入侵植物具有通用的基因型,有较强的生态适应性[2]、广泛的生态位[3];入侵植物具有高的光合速率[4],生长迅速,植株高大;入侵植物具有高的氮含量、高的光合氮分配策略[5]。基于这些研究,各学者提出了几大入侵机制假说:入侵进化和基因通用型假说、EICA假说、氮分配假说[6~8]。其中氮分配假说依赖于最佳的防御理论,天敌和竞争者造成的选择压力均会影响入侵植物氮的分配进化策略[9]。然而,氮分配假说是否具有通用性,入侵植物所体现出的优势是否优于本地植物,针对这些问题的研究甚少。
美洲商陆(Phytolacca ameyicana),商陆科商陆属,是一种产自北美洲东部的有强烈气味的高大草本植物,是我国很早发现的一种入侵植物[10]。为更好地了解商陆的入侵机制,选取美洲商陆及本地伴生种芒(Miscanthus sinensis)、铁芒萁(Dicranopteris linearis)、皱叶狗尾草(Setaria plicata)为研究对象;比较入侵植物与本地植物在不同光环境下的光合特征、光合氮分配的差异,探讨商陆的入侵机制,并验证氮分配假说是否具有通用性。也为治理入侵植物和防止入侵植物暴发提供一定的参考和理论依据。
2 材料与方法
2.1 研究区域概况
研究地点位于四川省宜宾市高县来复镇毛颠坳林区,地理位置介于东经104°33′25″~104°33′32″,北纬28°35′56″~28°36′08″,海拔高度400~450 m;属典型的中亚热带湿润季风气候,降雨量较大;年均温18.1 ℃。地带性植被类型为亚热带常绿阔叶林。现有林地是1972年人工造林后形成的马尾松人工纯林,行间距2.5 m×2.5 m,郁闭度0.6~0.8。土壤类型为山地黄壤,成土母岩为黄色砂岩,土壤基本理化性质见表1。林下植被盖度约30 %,主要灌木有青冈(Quercus serrata)、梨叶悬钩子(Rubus pirifolius)、野桐(Mallotus japonicus)、铁仔(Myrsine africana)、展毛野牡丹(Melastoma normale)等;主要草本有美洲商陆(Phytolacca americana)、蕨(Pteridium aquilinum)、芒(Miscan- thus sinensis)、芒萁(Dicranopteris dichotoma)、淡竹叶(Lophatherum guacile)等[11]。
2.2 样地设置及样品采集
试验选取年龄相同(1972年造林,平均株高16.2 m)、环境条件(坡向、坡度等)一致的马尾松純林样地,随机设置20 m×20 m(SG)、35 m×35 m(LG)的人工采伐林窗各4个,将树干残体及灌草植物的地上部分清出林窗,但未进行掘根处理。各林窗间距10 m以上。
2014年4月,于各林窗中央及其接近林下(US)和全伐旷地(OS)分别随机选取一年生草本植物美洲商陆、芒、铁芒萁、皱叶狗尾草各3株健康成熟叶片,进行光合参数测定。并采集相应叶片进行相关指标测定。
2.3 研究方法
在晴朗天气的上午9∶00~11∶30,采用LI-6400便携式光合仪测定植物叶片光响应曲线。条件控制:空气流速为500 μmol/s,叶片温度设置为25±1 ℃,相对湿度为60%±5%。在测定前,叶片在500~1500 μmol/(m2/s)人工光源条件下诱导20 min,然后测定不同光合有效辐射(PAR)梯度下的净光合速率Pn,每个PAR梯度平衡时间180 s。拟合Pn-PAR曲线求得饱和光强[12]。最大净光合速率(Pmax)是饱合光量子通量(PPFD)与380 μmol/mol CO2浓度下所得的净光合速率。在饱合光量子通量下,CO2浓度梯度为400、300、200、150、100、50、20、0、400、400、600、800、1000、1200 μmol/mol,按照各梯度测得的Pn与胞间二氧化碳浓度(Ci)值绘制Pn-Ci曲线,求得叶片光合潜力Pmax,最大羧化速率(Vcmax)和最大电子传递速率(Jmax)[13]。
每个样品叶片洗净后扫描叶片面积(SLA),然后放置于75 ℃的烘箱中,48 h后称重并计算比叶面积SLA;光合色素含量测定采用95%乙醇浸提法;用半微量凯氏定氮法测定叶片总氮含量;钼锑抗比色法测定叶磷含量。叶片氮利用效率PNUE=Pmax/NA。单位质量的光合速率Am=Pmax×SLA。
氮在光合机制中的分配系数计算[14,15]。
叶氮在羧化系统(主要是Rubisco)中的分配系数Pc,其计算式为:
Pc =Vcmax/(6.25×Vcr×NA)(1)
叶氮在生物力能学组份(光合电子传递和光合磷酸化载体蛋白组份)中的分配系数PB按照,其计算式为:
PB=Jmax/(8.06×Jmc×NA)(2)
叶氮在捕光色素蛋白复合体(PSI、PSⅡ、LHCⅡ)中的分配系数PL,其计算式为:
PL=CC/(NM×CB)(3)
叶氮在光合机制中的分配系数PT,其计算式为:
PT=PC+PB+PL(4)
式(1)、(2)、(3)、(4)中,NA是单位面积的氮含量(g/m2);NM为单位质量的氮含量(g/g);CC为叶绿素浓度(mmol/g);CB是结合在捕光色素蛋白复合体中的叶绿素,为5.79 mmol/g N;Vcr是单位Rubisco蛋白的固定活性;Jmc为单位细胞色素f(Cyt f)的最大电子传递速率;Vcr与Jmc的测定参见innemets和Tenhunen[16]。
2.4 数据处理
数据处理使用Excel 2003和SPSS 17.0,采用多因素单变量方差分析 (Univariate Multiv-ariate ANOVA)、单因素方差分析(One-way ANOVA),各参数的相关性分析采用Pearson双尾检验。
3 结果与分析
3.1 4个植物的最大净光合速率和单位重量的光合速率比较
不同植物间的最大净光合速率(Pmax)存在极显著差异(表2),芒[21.67~31.66 μmol/(m2/s)]的Pmax显著高于其他3个植物,美洲商陆的Pmax显著高于铁芒萁与皱叶狗尾草。芒的Pmax随着林窗面积增大而增大,除林下環境外,各环境中的芒的Pmax无显著差异。美洲商陆Pmax也表现为随着光环境增强而增大,大林窗中Pmax显著高于小林窗,但林下与小林窗、大林窗与旷地均无显著差异。铁芒萁和皱叶狗尾草的Pmax在不同林窗中均无显著差异(图1)。
在4个植物间,单位重量的光合速率(Am)存在显著性差异(表2),并按美洲商陆、芒、皱叶狗尾草、铁芒萁依次显著降低,其中美洲商陆的Am值最高[0.6652~0.8795 μmol/(s/g)],林下、小林窗和大林窗中,美洲商陆的Am均显著高于其他3个植物。美洲商陆、铁芒萁和皱叶狗尾草三者的Am随着光环境增大均呈逐渐减小的趋势;美洲商陆与皱叶狗尾草的Am在各林窗环境下有显著差异。铁芒萁的Am从小林窗到旷地显著降低,林下的Am与小林窗无显著差异。芒在各林窗中的Am表现出与其他植物相反的变化趋势,随着光环境增强而增大,林下与小林窗间有显著差异,旷地与大林窗差异不显著(图2)。
3.2 4个植物的叶氮含量与叶磷含量比较
不同植物间的叶氮含量存在极显著差异(表2),美洲商陆的叶氮含量最高(0.034~0.055 mg/g),显著高于芒、铁芒萁和皱叶狗尾草;芒的叶氮含量显著低于其余3个植物。各林窗环境中,美洲商陆、芒、铁芒萁均有显著性差异;铁芒萁与皱叶狗尾草在小林窗和大林窗中差异不显著,在旷地环境中有显著差异。四种植物在不同光环境梯度下的氮含量均呈显著变化,其中美洲商陆和铁芒萁的叶氮含量随着光环境增大而减小,芒的叶氮含量呈先增大与减小,而皱叶狗尾草的叶氮含量随光环境呈先减小后增大的趋势(图3)。
4个植物间的叶磷含量存在极显著差异(表2),美洲商陆的磷含量最高并显著高于其他3个植物,其次是皱叶狗尾草和铁芒萁,磷含量最低的植物是芒。林下环境,美洲商陆磷含量显著高于芒与铁芒萁;小林窗中,美洲商陆磷含量显著高于其余3个植物;大林窗中,美洲商陆与皱叶狗尾草的磷含量差异不显著,但皱叶狗尾草高于芒与铁芒萁;旷地中3个本地植物磷含量无显著差异。在不同林窗中,4个植物磷含量均有显著变化,随着林窗面积的增大,商陆、芒、皱叶狗尾草的磷含量呈先升高再降低的趋势(图4)。光环境梯度与物种差异的交互作用对叶磷含量的影响显著(表2)。
3.3 4个植物的比叶面积和叶绿素浓度比较
4个植物间的比叶面积(SLA)间存在极显著差异(表2)。4个林窗梯度下,美洲商陆的SLA均最高且显著高于芒、铁芒萁和皱叶狗尾草;芒的SLA显著最低。美洲商陆和皱叶狗尾草两者的SLA随着林窗面积增大而显著减小。铁芒萁随林窗面积增大而减小,但林下与小林窗的SLA无显著差异。相反,芒的SLA却呈随林窗面积增大而显著增大的变化(图5)。林窗大小与物种差异的交互作用对比叶面积的影响显著(表2)。
叶绿素浓度在4个植物间有显著差异(表2)。除林下的美洲商陆外,4个植物的叶绿素浓度(CC)均随着林窗面积增大而降低。从小林窗到旷地环境,美洲商陆的叶绿素浓度均高于其他3种植物,并与铁芒萁呈显著关系。林下环境中美洲商陆的叶绿素浓度显著小于芒与铁芒萁(图6)。
3.4 4个植物植物的光合氮分配系数特征
芒的PC值显著高于其余3个植物,美洲商陆和铁芒萁两者的PC值均相对较低。在各林窗梯度下,PC值均呈芒>皱叶狗尾草>美洲商陆>铁芒萁。芒PC值随林窗面积增大而增大,林下与小林窗有显著差异,大林窗与旷地有显著差异。铁芒萁表现出同芒相同的变化趋势,芒在林下环境中PC显著小于其他林窗。皱叶狗尾草在各林窗环境中PC值无显著差异。美洲商陆的PC值在林下环境中最大,在旷地最小;林下与小林窗中的PC显著高于大林窗与旷地环境(图7)。
芒的PB值显著高于其余3个植物,铁芒萁的PB值最小,显著小于美洲商陆。各林窗中,芒、美洲商陆、铁芒萁三者的PB依次显著减小。皱叶狗尾草PB在小林窗中与美洲商陆无显著差异,在大林窗中显著高于美洲商陆,旷地中却显著低于美洲商陆。在不同林窗面积梯度下,芒与美洲商陆的PB均随林窗面积增大而增大,且林下PB值显著小于大林窗及旷地环境。铁芒萁在各林窗中PB无显著差异。皱叶狗尾草的PB呈先增大后减小的趋势,旷地环境PB值显著小于林窗(图8)。
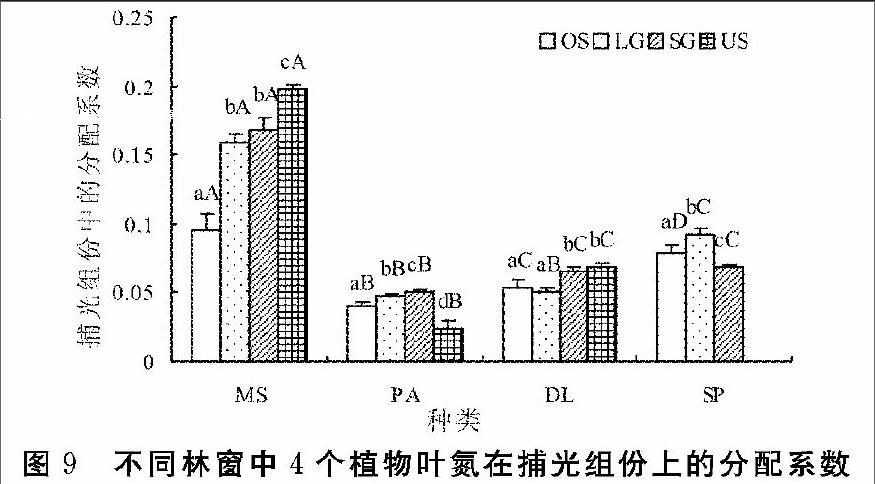
氮分配到捕光色素蛋白复合体组份(PL)在4个植物间,在美洲商陆的PL最小,显著小于其余3个植物;芒的PL显著高于美洲商陆、铁芒萁与皱叶狗尾草。随林窗面积增大,芒PL呈减小的趋势,芒在林下的PL显著高于林窗,在旷地的PL显著小于林窗。铁芒萁的PL在林下与小林窗中无显著差异,大林窗与旷地间也无显著差异,但大林窗PL显著小于小林窗。美洲商陆在林
下的PL显著小于其他林窗环境,在小林窗到旷地梯度下,PL呈显著减小的趋势。皱叶狗尾草从小林窗到旷地,PL呈先增大后减小,各林窗梯度下PL均有显著差异(图9)。
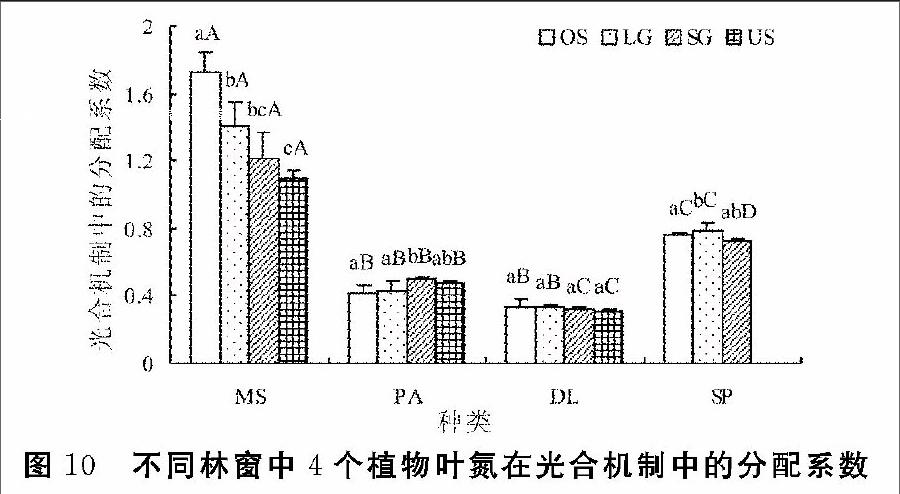
在4个植物间,芒的PT值仍显著高于其余3个植物;皱叶狗尾草PT显著高于美洲商陆与铁芒萁。林下和小林窗中,铁芒萁PT显著小于美洲商陆,大林窗和旷地中PT小于美洲商陆,但差异不显著。随林窗面积增大,芒的PT值显著增加。美洲商陆的PT在小林窗中显著高于大林窗和旷地。铁芒萁PT随林窗面积增大而增大,但无显著差异。皱叶狗尾草的PT在大林窗中显著高于小林窗与旷地,在小林窗与旷地间无显著差异(图10)。
3.5 4个植物的光合氮利用效率
4个植物间芒的PNUE最大,并显著高于其余3个植物。美洲商陆与皱叶狗尾草的PNUE无显著差异,两者均显著高于铁芒萁。随林窗面积增大,芒的PNUE呈增大的趋势,旷地的PNUE显著高于小林窗与林下,小林窗显著高于林下。铁芒萁的PNUE在各林窗中无显著差异。皱叶狗尾草呈先增大后减小的趋势,大林窗PNUE显著高于小林窗与旷地,小林窗与旷地间无显著差异。美洲商陆的PNUE在小林窗、大林窗、旷地三者间无显著差异,显著高于林下环境(图11)。
3.6 4个植物的各参数间的交互作用及相关系数
在4个植物中,林窗面积及物种差异对各项参数造成的差異及林窗面积与物种差异对参数的交互作用见表2,在PC、PB、PL、PT、NM、Am、SLA、PNUE、P、CC参数中,物种类型的不同与林窗面积的不同均对各参数造成极显著差异,其交互作用也达到极显著差异。林窗梯度对植物最大净光合速率造成显著差异,林窗梯度与物种类型对最大浄光合速率所起的交互作用无显著差异。
Pearson相关性分析表明,不同光环境下四种草本植物PC、PB、PL、PT、Am、SLA呈极显著的正相关关系;CC和NM、P、PNUE、Pmax呈极显著的正相关关系;NM与PC、PL、PT、Am呈极显著的负相关关系;P与PC、PT呈显著负相关关系,与PL呈极显著负相关关系,PL与PNUE、Pmax呈极显著负相关关系(表3)。
4 讨论
植物能成功入侵定伴随着超越本地植物的一些优点,有研究认为,入侵植物具有生物学基础和较宽的生态位、对环境的耐受范围较大,所以能成功入侵。如三裂蟛蜞菊(Wedelia trilobata)[17]、五爪金龙(Ipomoeacairica)[18]等入侵植物具有较大的光合生态位。同时,成功入侵的植物具有基因多样性,植物随生长环境的改变,表观基因组差异也较大,赋予了植物对环境的快速适应[19],促进入侵的成功。EICA假说提出,植物成功入侵是因为在新的环境中没有了天敌;减少了在防御组份上的资源分配,将更多的资源分配于植物生长组份中,表现为生长迅速,植株高大等,促进入侵。冯玉龙认为氮是植物分配的主要资源,入侵植物在防御组份上分配的氮减少,而将更多的氮分配于光合机构组份中。在该研究中,美洲商陆在4个植物间叶密度最小,SLA最大,氮含量最高;与多数入侵植物的研究结果一致[20,21]。美洲商陆的Pmax、PNUE和光合氮分配低于芒和皱叶狗尾草,因为芒与皱叶狗尾草两者是具有很强固氮能力的C4植物。美洲商陆的Pmax与PNUE均显著高于铁芒萁,体现出入侵植物的优势,与杜鹃与悬钩子的结果相同[22,23]。美洲商陆的Am显著高于其他3个物种,超越了植物在碳同化结构上的差异,成为入侵成功重要的因素之一。
入侵植物具有高于本地植物的氮含量[24],并将更多的氮分配于生长机制中。美洲商陆在4个植物间具有最高的氮含量;在光合氮分配组份上,美洲商陆的PB与PT显著高于铁芒萁、PL低于其余3个植物。碳同化结构相同的入侵植物与本地植物比较,美洲商陆通过高的叶氮向光合机构分配比例来取得入侵优势;氮向光合组份上分配的量增加,影响着PNUE的增加,美洲商陆入侵机制符合光合氮分配假说,与紫茎泽兰入侵机制相同[25]。美洲商陆的磷含量显著高于其他3个物种,磷的含量与植物的生命活动周期有关[26,27]。美洲商陆生长速率快,与本地植物的生长周期形成一个时间差,在本地植物生长初期占据较多的资源,在较短的时间内完成生命周期,这也成为美洲商陆成功入侵的一个重要原因。
5 结语
通过对不同林窗中4个植物的光合特征比较,发现美洲商陆具有较强的环境适应性,与薇甘菊(Mikania micrantha)相同[28]。在林下环境,美洲商陆叶绿素含量较小,通过增大比叶面积来获取更多资源,旷地环境中光资源丰富,表现出SLA显著降低。在不同的环境中,美洲商陆的光合氮分配、Pmax、PNUE等最为稳定。因此美洲商陆的较强适应性也促进其成功入侵。
通过分析得出,35 m×35 m环境中的美洲商陆入侵能力最强;美洲商陆具有较强的环境适应性、较宽的生态位,光合氮分配较高。但美洲商陆的光合速率、氮利用效率及光合氮分配仍未超越本地C4植物。入侵植物的入侵机制比较复杂,本研究也仅从光合特征及光合氮分配入手研究美洲商陆的入侵机制,入侵机制一直以来受生态学专家的密切关注及研究,希望能尽早攻克植物入侵带来的难题。
参考文献:
[1]闫淑君,洪 伟,吴承祯.生物入侵对福建生态安全的影响[J].福建林学院学报,2006,26(3):275~280.
[2]Christian C E.Consequences of a biological invasion reveal the importance of mutualism for plant communities[J].Nature,2001,413:635~639.
[3]王俊峰,冯玉龙,梁红柱.紫茎泽兰光合特性对生长环境光强的适应[J].应用生态学报,2004,15(8):1373~1377.
[4]Pattison R R,Goldstein G,Ares A.Growth,biomass allocation and photosynthesis of invasive and native Hawaiian rainforest species[J].Oecologia,1998,117(4):449~459.
[5]王满莲,冯玉龙,李 新.紫茎泽兰和飞机草的形态和光合特性对磷营养的响应[J].应用生态学报,2006,17(4):602~606.
[6]翁伯琦,林 嵩,王义祥.空心莲子草在我国的适应性及入侵机制[J].生态学报,2006,26(7):2373~2381.
[7]冯玉龙,廖志勇,张 茹,等.外来入侵植物对环境梯度和天敌逃逸的适应进化[J].生物多样性,2009,17(4):340~352.
[8]黄 伟,王 毅,丁建清.入侵植物乌桕防御策略的适应性进化研究[J].植物生态学报,2013,37(9):889~900.
[9]Feng Y,Lei Y,Wang R,et al.Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant [J].Proceedings of the National Academy of Sciences,2009,106(6):1853~1856.
[10]周 兵,闫小红,肖宜安,等.外来入侵植物美洲商陆的繁殖生物学特性及其与入侵性的关系[J].生态环境学报,2013(4):567~574.
[11]谭 波,吴福忠,杨万勤,等.不同林龄马尾松人工林土壤节肢动物群落结构[J].应用生态学报,2013,24(4):1118~1124.
[12]Farquhar G D,Ehleringer J R,Hubick K T.Carbon isotope discrimination and photosynthesis[J].Annual Review of Plant Biology,1989,40(1):503~537.
[13]胡启鹏,郭志华,李春燕,等.不同光环境下亚热带常绿阔叶树种和落叶阔叶树种幼苗的叶形态和光合生理特征[J].生态学报,2008,28(7):3262~3270.
[14]Feng Y L,Auge H,Ebeling S K.Invasive Buddleja davidii allocates more nitrogen to its photosynthetic machinery than five native woody species[J].Oecologia,2007,153(3):501~510.
[15]Niinemets U,Bilger W,Kull O,et al.Responses of foliar photosynthetic electron transport,pigment stoichiometry,and stomatal conductance to interacting environmental factors in a mixed species forest canopy[J].Tree Physiol,1999,19(13):839~852.
[16]Niinemets U,Kull O,Tenhunen J D.An analysis of light effects on foliar morphology,physiology, and light interception in temperate deciduous woody species of contrasting shade tolerance[J].Tree Physiol,1998,18(10):681~696.
[17]宋莉英,孫兰兰,舒 展,等.干旱和复水对入侵植物三裂叶蟛蜞菊叶片叶绿素荧光特性的影响[J].生态学报,2009,29(7):3713~3721.
[18]吴彦琼,胡玉佳.外来植物南美蟛蜞菊、裂叶牵牛和五爪金龙的光合特性[J].生态学报,2004,24(10):2334~2339.
[19]Schmitz R J,Schultz M D,Urich M A,et al.Patterns of population epigenomic diversity[J].Nature,2013,495(7440):193~198.
[20]Baruch Z,Goldstein G.Leaf construction cost,nutrient concentration,and net CO2 assimilation of native and invasive species in Hawaii[J].Oecologia,1999,121(2):183~192.
[21]Gulías J,Flexas J,Mus M,et al.Relationship between Maximum Leaf Photosynthesis,Nitrogen Content and Specific Leaf Area in Balearic Endemic and Non-endemic Mediterranean Species[J].Annals of Botany,2003,92(2):215~222.
[22]Mcdowell S C.Photosynthetic characteristics of invasive and noninvasive species of Rubus(Rosaceae)[J].American Journal of Botany,2002,89(9):1431~1438.
[23]Niinemets ,Valladares F,Ceulemans R.Leaf‐level phenotypic variability and plasticity of invasive Rhododendron ponticum and non‐invasive Ilex aquifolium co-occurring at two contrasting European sites[J].Plant,Cell & Environment,2003,26(6):941~956.
[24]邓 雄.入侵植物薇甘菊与其本地同属种蔓泽兰的叶构建因子比较研究[J].中山大学学报(自然科学版),2010,49(1):80~82.
[25]Feng Y L,Lei Y B,Wang R F,et al.Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant[J].Proceedings of the National Academy of Sciences,2009,106(6):1853~1856.
[26]Niinemets .Global-scale climatic controls of leaf dry mass per area,density,and thickness in trees and shrubs[J].Ecology,2001,82(2):453~469.
[27]胡啟鹏,郭志华,李春燕,等.不同光环境下亚热带常绿阔叶树种和落叶阔叶树种幼苗的叶形态和光合生理特征[J].生态学报,2008,28(7):3262~3270.
[28]祖元刚,张衷华,王文杰,等.薇甘菊叶和茎的光合特性[J].植物生态学报,2006,30(6):998~1004.
