
普通烟草RLP类受体蛋白家族成员的鉴定与进化、表达分析
2017-05-10 03:38:57李晓旭段奇佳郭永峰
中国烟草科学 2017年2期
李 奎,李晓旭,刘 成,段奇佳,郭永峰*
普通烟草RLP类受体蛋白家族成员的鉴定与进化、表达分析
李 奎1,2,李晓旭1,2,刘 成1,3,段奇佳1,郭永峰1*
(1.中国农业科学院烟草研究所,青岛 266101;2.中国农业科学院研究生院,北京 100081;3. 云南省烟草公司曲靖市公司,云南曲靖 655000)
类受体蛋白(receptor-like proteins, RLPs)作为一类细胞表面受体,广泛存在于高等植物中,参与调控植物生长发育和抗逆等过程。本研究以普通烟草()TN90数据库为基础,构建隐马尔科夫模型进行检索,鉴定了普通烟草RLP家族成员;采用邻接法构建系统发育树进行系统进化分析;利用GSDS对RLP家族成员的基因结构进行了分析;利用普通烟草TN90转录组数据,分析基因家族成员的表达情况;最后对普通烟草RLP家族成员进行GO注释分析。结果表明,在普通烟草中共鉴定出70个RLP家族成员,各成员的氨基酸序列长度和等电点差异较大;系统进化与基因结构分析表明,普通烟草家族成员划分为6个亚家族,基因结构较为简单;转录组分析结果显示,基因家族成员在不同组织和发育时期的表达有较大差异,个别成员在叶片衰老时期表达量较高;GO注释结果表明,RLPs在植物的生长发育、逆境响应等过程中发挥作用。本研究结果将为普通烟草基因功能分析和利用奠定基础。
普通烟草;基因家族;生物信息学分析
植物细胞表面受体以一种复杂的方式转导细胞内外各种信号。植物类受体蛋白(receptor-like proteins, RLPs)是一类重要的细胞表面受体[1],从结构来看,RLPs由一个单向跨膜域、两个eLRR域以及一个短尾细胞质域组成,eLRR域中通常含有许多潜在的糖基化位点,表明RLPs可能是锚定于细胞膜上的糖蛋白受体[2]。
RLP蛋白在植物的生长与发育过程中发挥着重要作用[3]。拟南芥鉴定出57个基因[4],杨树鉴定出80个基因[5],拟南芥CLV2和其玉米中的功能同源蛋白FEA2调控着分生组织的维持与分化以及相关器官的发育[6-7]。TMM是调控拟南芥气孔发育的关键因子[8]。RLP类受体蛋白也参与植物激素的信号传导,如AtRLP41调控拟南芥叶片对ABA的敏感性[4],AtRLP44参与激活油菜素内酯信号途径[9]。
前期研究表明,多数RLP蛋白参与植物抗病过程[10]。包括苹果抗黑星病菌蛋白HcrVf2[11]、番茄抗叶霉菌Cf类蛋白[12]、番茄抗黑白轮枝菌Ve类受体蛋白[13]、油菜茎基溃疡病菌小种特异性抗性蛋白LepR3[14]等。在拟南芥中也发现了抗病蛋白,如AtRLP01能够识别来自黄单胞菌的PAMP信号ReMAX,参与抗病反应[15]; AtRLP52参与调控拟南芥对二孢白粉菌的防卫响应[16];AtRLP51参与NPR1介导的植物免疫反应[17];AtRLP42能够识别真菌PAMP信号多聚半乳糖醛酸酶(PG)信号[18]。
RLP蛋白生物学功能研究主要集中在拟南芥,如CLV2、TMM[19]等,以及其他诸如苹果HcrVf2蛋白、番茄Cf和Ve蛋白及小麦TaRLP1.1蛋白[20],而烟草RLP蛋白的研究还未见报道。研究普通烟草基因结构、表达模式及其所编码蛋白的理化性质,有利于进一步研究其在烟草生长发育过程中的功能。本研究采用生物信息学方法从普通烟草基因组中鉴定了70个基因,并分析了家族成员的进化关系、基因结构、表达情况等,用Blast2GO进行GO注释,对其生物学功能进行预测,为开展普通烟草基因的功能研究奠定了基础。
1 材料与方法
1.1 普通烟草RLP家族成员的鉴定及序列分析
根据关于AtRLPs的文献[4],从TAIR数据库(http://www.arabidopsis.org/)下载了拟南芥RLP家族蛋白全长序列。在茄科植物数据库(http://solgenomics.net/)下载普通烟草TN90蛋白序列。用构建的隐马尔科夫模型检索普通烟草TN90蛋白序列数据库,获得RLP候选序列,然后用InterPro(http://www.ebi.ac.uk/interpro/)工具剔除不含LRR的蛋白,得到普通烟草的RLP。利用ExPASy Proteomics Server(http://web.expasy.org/protparam/)工具对RLP家族成员的氨基酸长度、分子量大小、蛋白质疏水性、理论等电点等进行分析。
1.2 普通烟草RLP家族系统发育分析
参照文献[21]中方法,将预测的普通烟草RLP与拟南芥RLP蛋白全长序列进行比对。根据比对结果,用MGEA5重建邻接发育树。
1.3 普通烟草基因结构分析
在茄科植物数据库(https://solgenomics.net/),下载普通烟草TN90全基因组数据,用Perl程序分析所提取的基因的结构信息,最后使用GSDS(http://gsds.cbi. pku.edu.cn/)工具对普通烟草基因结构进行可视化展示。
1.4 普通烟草基因表达分析及GO注释分析
在NCBI SRA(http://www.ncbi.nlm.nih.gov/sra)下载普通烟草TN90转录组(RNA-seq)数据,将表达数据标准化后用R语言pheatmap程序绘制热图(heatmap),将其表达数据可视化。用Blast2GO对普通烟草RLP蛋白序列进行GO(Gene Ontology)注释分析,利用WEGO工具对注释结果进行进行可视化展示。
2 结 果
2.1 普通烟草RLP家族成员鉴定及其理化性质分析
基于WANG等[4]对拟南芥RLP家族成员的分析,在普通烟草全基因组中鉴定出70个家族基因,将其命名(表1)。对普通烟草RLP家族成员进行蛋白序列和理化性质分析发现,RLP家族成员间氨基酸的长度差异较大,介于119aa(NtRLP63)~1136aa(NtRLP53),蛋白分子量则介于13.40 kD~83.55 kD。NtRLP家族成员蛋白的理论等电点从4.49(NtRLP67)到9.26(NtRLP38)不等,变化范围较大,其中第II亚组中除了NtRLP51理论等电点(PI=8.56)大于7.0在碱性范围内,其他成员的理论等电点小于7.0(PI<7.0),均在酸性范围内,说明这些成员富含酸性氨基酸;第III亚组中,NtRLP19(PI=7.82)富含碱性氨基酸,其余成员的理论等电点小于7.0(PI<7.0),表明其富含酸性氨基酸;第VI亚组除了NtRLP07(PI=7.18)富含碱性氨基酸外,其他成员富含酸性氨基酸(PI<7.0),说明第II、III和VI亚家族绝大部分成员的蛋白由较多的酸性氨基酸组成。NtRLP家族中有26个成员蛋白疏水性为正值,说明其为疏水性蛋白;44个成员蛋白疏水性为负值,表明其为亲水性蛋白。
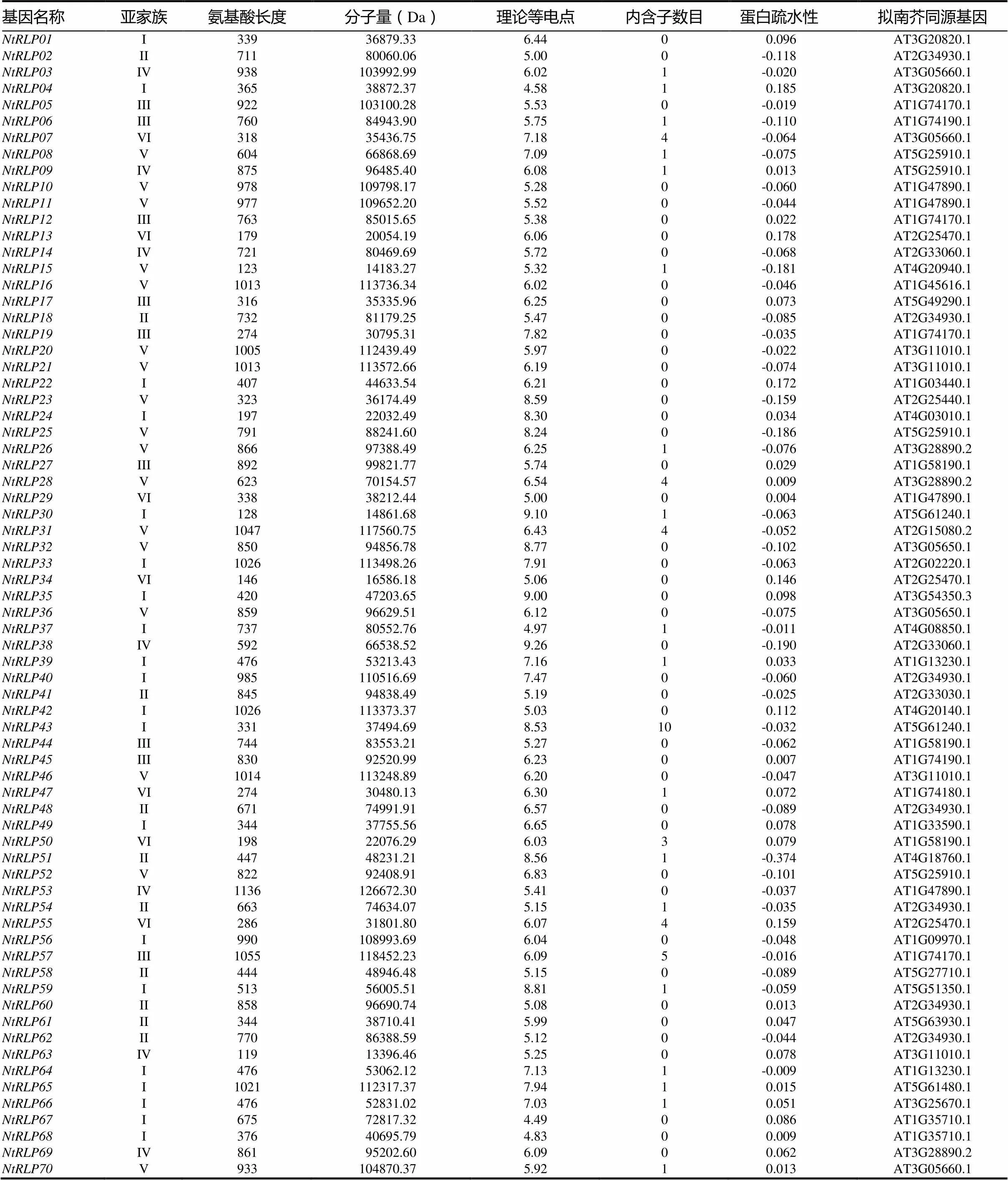
表 1 普通烟草RLP家族成员理化性质分析
2.2 普通烟草RLP家族系统进化与基因结构分析
以拟南芥RLP家族为参考,根据序列比对的结果,利用MEGA重构了邻接树(图1)。系统进化分析显示,普通烟草70个RLP家族成员可划分为6个亚家族,第I~VI亚家族的成员个数分别为20、10、9、7、17、7。在6个亚家族中,第VI亚家族较其他5个亚家族亲缘关系较远,是一个较为独立的分支,而又与该家族其他成员的亲缘关系较远,是比较特殊的一个。
利用GSDS构建普通烟草基因内含子-外显子结构图(图2)。基因结构分析显示,在家族70个成员中,45个成员没有内含子,17个只有1个内含子,5个成员有4个内含子,剩余基因所内含子数目不等。第I亚组中,有10个内含子,是家族中最多的;在第III亚家族中,除、外,其余成员均没有内含子。整体来看,基因家族成员结构较为简单。
2.3 普通烟草基因家族的基因表达分析
基于TN90转录组(RNA-seq)数据,将普通烟草基因的表达数据标准化后用R语言进行可视化展示(图3)。图3显示,普通烟草基因在根、茎、幼叶、成熟叶、衰老叶等5个组织中都有表达。其中,、两个基因在根、茎、幼叶、成熟叶、衰老叶5个组织中表达量都较高,说明其可能在整个发育过程中都起作用。进一步分析发现,少数的基因的表达具有一定的组织特异性。特别是、、、四个基因在成熟叶中的表达量较高,表明它们可能参与调控营养生长向生殖生长过程的转换。而仅在根部表达,可能调控根部的发育;、、只在茎部表达,在其他四个组织没有表达,表明其与茎部的发育密切相关。有些基因的表达具有一定的规律性,如、随着生长发育的进行表达量逐渐升高,在衰老叶片中表达量达到最高,而、只在衰老叶片中表达,据此可以推测它们可能参与调控叶片的衰老进程。
2.4 普通烟草RLP家族GO注释分析
我们对普通烟草RLP蛋白序列从细胞组成(Cellular Component)、分子功能(Molecular Function)和参与的生物学过程(Biological Process)三个方面进行GO注释(图4)。Go注释结果表明,在细胞组成和分子功能方面,普通烟草的RLP蛋白在细胞表面发挥结合和转运作用,这与RLP蛋白作为一类细胞表面受体的功能相吻合。RLP家族成员的生物学功能呈多样化,主要参与细胞物质代谢、酶代谢、多细胞组织形成、生长发育等过程。
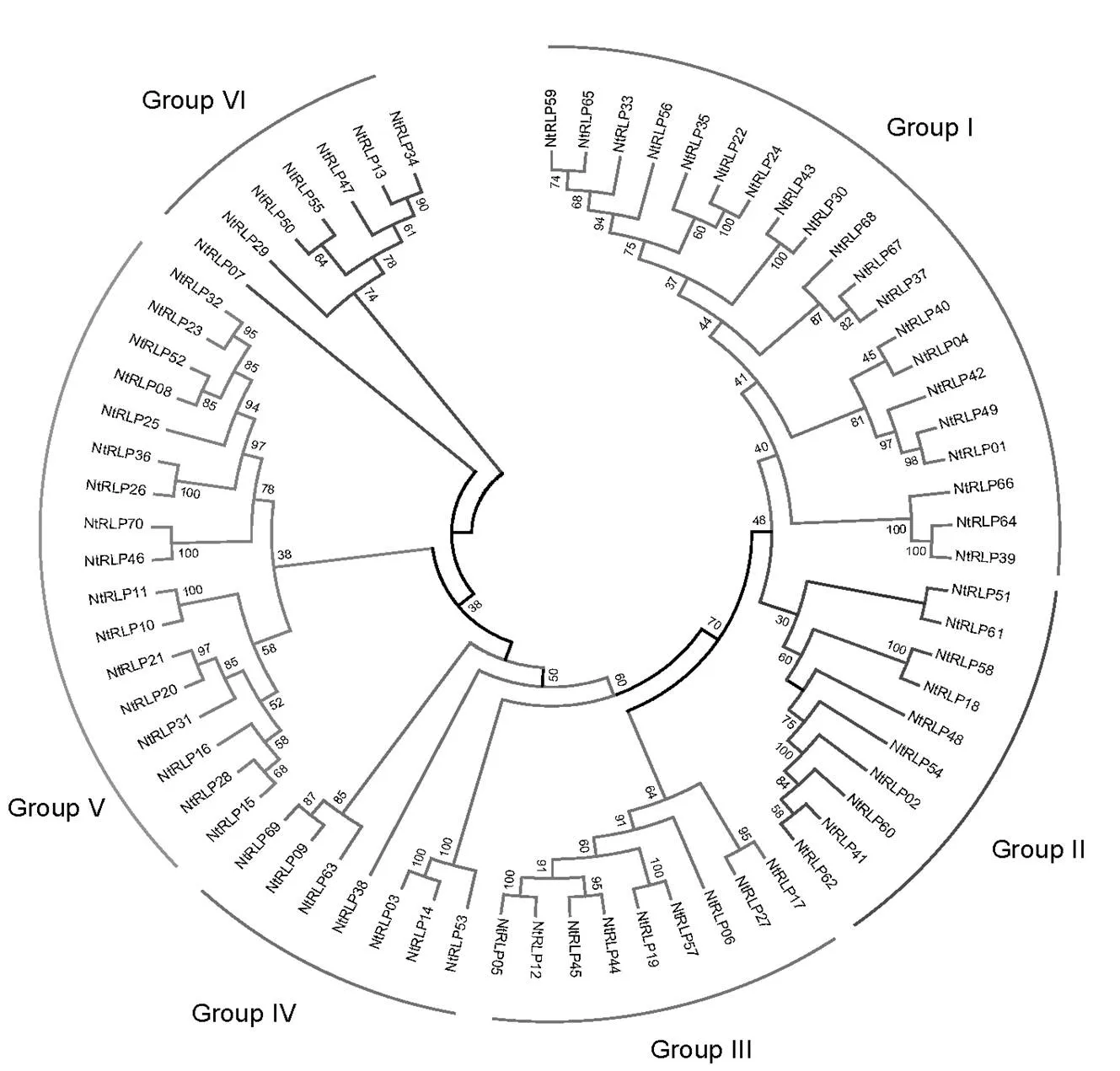
图 1 普通烟草RLP家族成员邻接进化树

注:T1,根;T2,茎;T3,幼叶;T4,成熟叶;T5,衰老叶。
3 讨 论
RLP类受体蛋白广泛参与植物生长发育、逆境胁迫响应、植物激素信号传导等过程。拟南芥CLV2与CRN结合形成传导CLV3信号受体复合体,调控拟南芥茎顶端分生组织发育[22],小麦受体类蛋白TaRLP1.1参与小麦抗条锈病响应过程[20]。SOBIR1与LerR3、AtRLP30等蛋白相互作用,参与调控植物抗病过程[23-24]。
本研究从普通烟草基因组中鉴定出70个基因。系统发育分析表明,普通烟草的家族成员被分成了个6亚家族。第IV亚组中,只有有内含子,其余成员没有内含子;第VI亚组成员基因结构差异较大。GO注释表明,RLP在细胞表面发挥结合与转运功能,这与其作为受体蛋白相吻合。WANG等[4]的研究表明可能参与调控拟南芥叶片衰老过程。我们利用Arabidposis eFP Broswer(http://bbc.botany.utoronto.ca)数据库分析家族成员表达模式,发现、、等在衰老叶片中表达水平非常高,可能参与拟南芥叶片衰老调控过程。普通烟草基因表达模式分析发现,与同源的具有类似的衰老组织表达特异性,随着叶片落黄其表达量显著上调,衰老叶片中表达量达到最高,其可能参与普通烟草叶片衰老的调控过程。特别需要指出的是,,两个基因只在衰老的叶片中表达,这表明他们同样可能参与普通烟草叶片衰老的调控。本研究对普通烟草基因家族进行了全基因组鉴定、表达模式分析和功能注释等分析,但其具体的生物学功能还有待于利用分子生物学方法进行进一步验证。
[1] WU J B, LIU Z J, ZHANG Z, et al. Transcriptional regulation of receptor-like protein genes by environmental stresses and hormones and their overexpression activities in Arabidopsis thaliana[J]. Journal of Experimental Botany, 2016, 67(11): 3339-3351.
[2] TÖR M, LOTZE M T, HOLTON N. Receptor-mediated signaling in plants: molecular patterns and programmers[J]. Journal of Experimental Botany, 2009, 60(13): 3645-3654.
[3] WANG G D, FIERS M, ELLENDORFF U, et al. The diverse roles of extracellular leucine-rich repeat- containing receptor-like proteins in plants[J]. Critical Reviews in Plant Sciences, 2010, 29: 285-299.
[4] WANG G D, ELLENDORFF U, KEMP B, et al. A genome-wide functional investigation into the roles of receptor-like proteins in Arabidopsis[J]. Plant Physiol, 2008, 147(2): 503-517.
[5] PETRE B, HACQUARD S, DUPLESSIS S, et al. Genome analysis of poplar LRR-RLP gene clusters reveals RISP, a defense-related gene coding a candidate endogenous peptide elicitor[J]. Frontiers in plant science, 2014, 5(111): 1-10.
[6] WANG G D, ZHANG Z, ANGENEN G C, et al. New aspects of CLAVATA2, a versatile gene in the regulation ofdevelopment[J]. Journal of Plant Physiology, 2011, 168(4): 403-407.
[7] TAGUCHI-SHIOBARA F, YUAN Z, HAKE S, et al. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize[J]. Genes & Development, 2001, 15(20): 2755-2766.
[8] NADEAU J A, SACK F D. Control of stomatal distribution on the Arabidopsis leaf surface[J]. Science, 2002, 296(5573): 1697-1700.
[9] WOLF S, VAN DER DOES D, LADWIG F,et al. A receptor-like protein mediates the response to pectin modification by activating brassinosteroid signaling[J]. PNAS, 2014, 111(42):15261-15266.
[10] KRUIJT M, DE KOCK M J D, DE WIT P J G M. Receptor-like proteins involved in plant disease resistance[J]. Molecular plant pathology, 2005, 6(1): 85-97.
[11] BELFANTI E, SILFVERBERG-DILWORTH E, TARTARINI S, et al. The HcrVf2 gene from a wild apple confers scab resistance to a transgenic cultivated variety[J]. Plant Biology, 2004, 101: 886-890.
[12] THOMMA B P H J, VAN ESSE H P, CROUS P W, et al. Cladosporium fulvum (syn. Passalora fulva), a highly specialized plant pathogen as a model for functional studies on plant pathogenic Mycosphaerellaceae[J]. Molecular Plant Pathology, 2005, 6(4): 379-393.
[13] KAWCHUK L M, HACHEY J, LYNCH D R, et al. Tomato Ve disease resistance genes encode cell surfacelike receptors[J]. Proceedings of the National Academy of Sciences, 2001, 98(11): 6511-6515.
[14] LARKAN N J, LYDIATE D J, PARKIN I A, et al. The Brassica napus blackleg resistance gene LepR3 encodes a receptor-like protein triggered by the Leptosphaeria maculans effector AVRLM1[J]. New Phytologist, 2013, 197: 595-605.
[15] JEHLE A K, LIPSCHIS M, ALBERT M, et al. The receptor-like protein ReMAX of Arabidopsis detects the microbe-associated molecular pattern eMax from Xanthomonas[J]. The Plant Cell, 2013, 25: 2330-2340.
[16] RAMONELL K, BERROCAL-LOBO M, KOH S, et al. Loss-of-function mutations in chitin responsive genes show increased susceptibility to the powdery mildew pathogen Erysiphe cichoracearum[J]. Plant physiology, 2005, 138(2): 1027-1036.
[17] ZHANG Y, YANG Y, FANG B, et al. Arabidopsis snc2-1D activates receptor-like protein-mediated immunity transduced through WRKY70[J]. The Plant Cell, 2010, 22: 3153-3163.
[18] ZHANG L, KARS I, ESSENSTAM B, et al. Fungal endopolygalacturonases are recognized as microbe- Associate molecular patterns by the Arabidopsis receptor-like protein RESPONSIVENESS TO BOTRYTIS POLYGALACTURONASES1[J]. Plant physiology, 2014, 164: 352-364.
[19] LYU Y, YANG N, WU J, et al. New insights into receptor-like protein functions in Arabidopsis[J]. Plant signaling & behavior, 2016, 11: 7, e1197469.
[20] JIANG Z, GE S, XING L, et al. RLP1. 1, a novel wheat receptor-like protein gene, is involved in the defence response against Puccinia striiformis f. sp. tritici[J]. Journal of experimental botany, 2013, 64(12): 3735-3746.
[21] 李晓旭,刘成,李伟,等. 番茄WOX转录因子家族的鉴定及其进化、表达分析[J]. 遗传,2016,38(5):444-460.
[22] BLECKMANN A, WEIDTKAMP-PETERS S, SEIDEL C A, et al. Stem cell signaling in Arabidopsis requires CRN to localize CLV2 to the plasma membrane[J]. Plant physiology, 2010, 152: 166-176.
[23] MA L, BORHAN M H. The receptor-like kinase SOBIR1 interacts with Brassica napus LepR3 and is required for Leptosphaeria maculans AvrLm1-triggered immunity[J]. Frontiers in plant science, 2015, 6(6): 1-10.
[24] ZHANG W, FRAITURE M, KOLB D, et al. Arabidopsis receptor-like protein30 and receptor-like kinase suppressor of BIR1-1/EVERSHED mediate innateBIR1-1/ EVERSHED mediate innate immunity to necrotrophic fungi[J]. The Plant Cell, 2013, 25(10): 4227-4241.
Genome-wide Identification and Expression Analysis of theGene Family in
LI Kui1,2, LI Xiaoxu1,2, LIU Cheng1,3, DUAN Qijia1, GUO Yongfeng1*
(1. Tobacco Research Institute of Chinese Academy of Agricultural Sciences, Qingdao 266101, China; 2. Graduate School of Chinese Academy of Agricultural Sciences, Beijing 100081, China; 3. Qujing Tobacco Company of Yunnan, Qujing, Yunnan 655000, China)
Receptor-like proteins (RLPs) as an important class of cell surface receptors, are present in many plant species. RLPs have been implicated in multiple biological processes including plant development and stress responses. In this study, 70 RLP family genes were identified in. Phylogenetic analysis was performed using the neighbor-joining method and the results showed that thesefamily members could be classified into 6 subfamilies. Transcriptome analysis revealed that the expression patterns ofwere distinct in different tissue types and at different developmental stages, with a number of genes being up-regulated in senescing leaves. GO analysis suggested that the RLP family members could be potentially involved in a number of biological processes including developmental regulation and stress responses. The results of this study could be valuable resources for future investigation of the functions ofs in.
;family; bioinformatic analysis
S572.03
1007-5119(2017)02-0063-06
10.13496/j.issn.1007-5119.2017.02.011
中国农业科学院基本科研业务费增量重点项目“烟草叶片衰老的分子调控机制研究”(2013ZL024)
李 奎(1989-),男,硕士研究生,研究方向:烟草分子育种,E-mail:lk928346616@126.com。
,E-mail:guoyongfeng@caas.cn
2016-11-17
2017-02-19
猜你喜欢
奥秘(创新大赛)(2023年3期)2023-05-06 01:48:20
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01 08:10:30
生物工程学报(2019年6期)2019-07-10 08:38:38
生物学通报(2019年1期)2019-02-15 16:33:43
生物学通报(2018年12期)2018-10-10 06:52:36
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
浙江中西医结合杂志(2017年2期)2017-01-12 18:23:59
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
