
喀纳斯自然保护区林地和草地土壤呼吸速率差异及影响因素①
2017-04-24 09:37:19范子昂窦晓静吉春容黄有志
土壤 2017年1期
范子昂,窦晓静,邹 陈,吉春容,黄有志
(1 南京信息工程大学环境科学与工程学院,南京 210044;2 中国气象局乌鲁木齐沙漠气象研究所,中国气象局树木年轮理化研究重点实验室/新疆维吾尔自治区树木年轮生态重点实验室,乌鲁木齐 830002;3 新疆农业科学院土壤肥料与农业节水研究所,乌鲁木齐 830091;4 布尔津县气象局,新疆布尔津 836600)
喀纳斯自然保护区林地和草地土壤呼吸速率差异及影响因素①
范子昂1,2,窦晓静3,邹 陈2*,吉春容2,黄有志4
(1 南京信息工程大学环境科学与工程学院,南京 210044;2 中国气象局乌鲁木齐沙漠气象研究所,中国气象局树木年轮理化研究重点实验室/新疆维吾尔自治区树木年轮生态重点实验室,乌鲁木齐 830002;3 新疆农业科学院土壤肥料与农业节水研究所,乌鲁木齐 830091;4 布尔津县气象局,新疆布尔津 836600)
为了研究新疆喀纳斯国家自然保护区森林生态和草地生态系统与大气的相互作用,分析土壤呼吸速率的时空变异特征及与影响因子的关系,2012年及2013年5月上旬至9月上旬利用土壤碳通量测量系统LI-8150对林地和草地两种植被类型土壤的呼吸速率日动态进行了全天连续自动监测,研究了两类土壤呼吸速率在生长季各月日的变化规律。结果表明:林地和草地生态系统土壤呼吸速率在植被生长季内均呈现较明显的单峰曲线型日变化,其月最大值均出现在7月,林地土壤呼吸速率的最大值(2.43 µmol/(m2·s))显著高于草地土壤呼吸速率最大值(1.55 µmol/(m2·s)),且各月林地生态系统的土壤呼吸速率均明显高于草地,波峰出现在北京时间17:00—18:00,波谷出现在北京时间9:00—10:00,草地最小值出现在6月(0.35 µmol/(m2·s)),而林地最小值出现在5月,其呼吸速率仅 0.71 µmol/(m2·s),且整个生长季白天大于晚上,全天以呼出CO2为主。植被生长季内林地和草地土壤呼吸速率与土壤温度的变化趋势相似,具有显著的正相关性,但与土壤含水量的变化没有这样的趋势。
喀纳斯自然保护区;土壤呼吸速率;土壤温度;土壤湿度
土壤作为多孔介质其空隙内储存着气体成分(包括CO2、O2、N2O等),同时,土壤也可能吸附着大量的气体分子,土壤气体成分主要来源于土壤中一系列化学和生物化学反应的直接产生以及地面空气向土壤中的扩散,土壤中气体含量的多少及其空间分布,主要取决于土壤中的产生过程、大气向下扩散过程以及土壤气体向大气排放过程综合作用的结果[1]。土壤是陆地生态系统中主要的碳库[2–4],其有机碳总储量约1 300 ~ 2 000 Pg[5],每年通过土壤呼吸向大气中释放的CO2约占生物圈排放的20% ~ 38%[6],是土壤碳素向大气输出的主要途径,其微小变化会对大气CO2浓度产生深远影响,并可能与气候变化产生正反馈效应[7–9]。土壤呼吸已成为碳元素由陆地生态系统进入大气的最主要途径和大气CO2重要的源,因此,精确测定土壤呼吸速率,开展不同区域、不同下垫面土壤呼吸相关研究,并深入探索其与环境因子的关系、土壤生态系统物质循环与能量转化及全球陆地生态系统碳循环,可为全球碳平衡预算和全球气候变化潜在效应估计提供科学依据和基础数据[10]。
喀纳斯国家自然保护区位于新疆北部阿尔泰山中段,是中国唯一与四国接壤的自然保护区,也是我国唯一的西伯利亚泰加林分布区,成为阻滞古尔班通古特沙漠北移的重要生态屏障。区内原始植物生态系统保存完好,具有完整的垂直带谱[11],是陆地生态系统对全球气候变化响应研究的理想实验区。西伯利亚落叶松(Larix sibirica Ldb.)以及西伯利亚云杉(Picea obovata Ldb.)构成了喀纳斯森林生态系统的主体[12]。目前对喀纳斯自然保护区森林生态系统的研究主要集中在物种多样性、群落格局及演替和干扰对森林的影响,以及植被 LAI的变化特征及其与植被生物量之间关系[13–19],对该区域森林、草地混合区植物的生理生态特性及其参与陆地化学循环过程的研究未见相关报道。本研究以喀纳斯保护区内的阿勒泰山山地草原、西伯利亚落叶松、天山云杉生态区为研究对象,研究森林生态和草地生态系统与大气相互作用,分析土壤呼吸速率的时空变异特征及与影响因子的关系,为进一步探讨喀纳斯乃至整个新疆北部森林和草地生态系统碳收支平衡及其变化提供依据,为研究陆地–大气相互作用奠定基础。
1 研究区概况与研究方法
1.1 研究区概况
喀纳斯国家级自然保护区位于新疆维吾尔自治区阿勒泰地区的布尔津县境内,地理位置 48°34′ ~ 49°11′ N;86°54′ ~ 87°54′ E,北与哈萨克斯坦共和国、俄罗斯共和国接壤,东与蒙古人民共和国毗连,西与哈巴河县相望,南与布尔津林场为邻,土地总面积21.92万hm2。该林区气候寒冷湿润,冬季漫长而冷,年平均气温 –0.2℃,气温在0℃以下的时间长达5 ~ 7个月,土壤冻结深度2 ~ 3 m,1月份平均气温 –20℃,7月份平均气温15℃,植物生长期为100 ~ 200 d,年均降水量1 065.4 mm,年均蒸发量为1 097.0 mm。受第四纪冰川和北冰洋气候的影响,该区形成了特殊的自然景观和植被类型。气候、土壤和生物分布具有明显的垂直带谱,森林、草原、草甸相间交错分布。其中,海拔1 300 ~ 2 300 m范围内的亚高山寒温带针叶林——山地棕色针叶林带属于典型的泰加林区,其优势树种为西伯利亚特有种,也是我国保存较为完好的泰加林类型之一。观测场林地内主要的森林群落建群种有西伯利亚云杉(P. obovata),其伴生树种为西伯利亚落叶松(L. sibirica)、西伯利亚冷杉(Abies sibirica),林木平均胸径18.2 cm,林分密度1 441株/hm2,平均树高13.5 m,土壤平均厚度15 cm,土层下面是岩石层,坡向为西南偏西,坡度为26°。林下优势植物有:林地早熟禾(Poa nemorensi L.)、紫苞鸢尾(Iris ruthenica Ker-Gawl.)、森林草莓(Fragaria vesca L.)、白花砧草(Galium boreale L.)、石生悬钩子(Rubus saxatilis L.)等[20–21]。
1.2 研究方法
土壤呼吸日动态测定采用开路式 8通道土壤碳通量测量系统LI-8150 (LI-COR,Lincoln,NE,USA),分别采用两个呼吸气室先后对林地和草地两类土壤的呼吸速率日动态进行全天连续自动监测,一天 24时次。LI-8150利用测量室内CO2浓度的变化速率推算气室所覆盖的样地其土壤释放或吸收 CO2速度。为了保证推算结果的正确,测量室内外的浓度梯度、气压、土壤温湿度应该相似。观测试验于2012年及2013年每年5月上旬至9月上旬(喀纳斯自然保护区开放)在新疆布尔津气象局贾登禹气象站内进行。
选择两个相对较为平坦的典型样地(5 m × 5 m),在这两个不同植被土壤表面上分别设立一个全天连续自动监测气室。一个气室设置在山地草地上,选择优势自然植被为早熟禾、白花砧草、石生悬钩子集中分布的土地为研究对象;另一个气室设置在林地内,选择以西伯利亚落叶松、西伯利亚云杉、西伯利亚冷杉为优势树种森林生态系统集中分布的土地为研究对象。
为避免由于测定时基座的嵌入对土壤扰动而造成短期内土壤呼吸速率的波动,提前2 d将横截面积为371.8 cm2、高度为10 cm的测定基座(圆柱形聚氯乙烯环)嵌入两个观测样地中。实施过程中注意将测定基座削尖的一端压入土中,以减少布置土壤环对土壤的镇压作用;同时,考虑到凋落物厚度和表层根系的分布,插入土壤的测定基座深度约5.8 ~ 6.0 cm,以减少测定基座对植被根系的破坏;另外,尽可能不扰动地表的凋落物和土壤,保持土壤环在整个测定期间位置不变。在等待48 h平衡后,土壤呼吸速率基本恢复到放置前水平后,再将土壤呼吸气室分别安装在2个测定基座上。由于喀纳斯地区土壤呼吸作用较微弱,降水较多[22],为保证测量精度在测量前已对分析器进行全面校正,并设置每个呼吸气室抽取气体前的稳定时间为45 s,抽气测量时间设定为150 s,测量后抽气时间为30 s,每个整点开始依次轮流测定山地草地和林地的土壤呼吸两次取平均,两次测定期间间隔45 s。测定土壤呼吸的同时,用仪器自带的土壤温度、含水量传感器对山地草地和林地0 ~ 5 cm土层土壤平均温度和含水量进行同步测定。
1.3 数据处理
实验数据采用 Microsoft Excel 2007(Microsoft公司,美国)进行统计分析及绘图,SPSS 20.0(IBM公司,美国)进行植被土壤呼吸、土壤温度和土壤含水量的相关及回归分析。
2 结果与分析
土壤呼吸是一个十分复杂的土壤生物学过程,严格意义上讲是指未扰动土壤中产生 CO2的所有代谢作用,主要包括植物的根系呼吸、土壤微生物呼吸、土壤动物呼吸3个生物学过程和一个非生物学过程,即含碳矿物质的化学氧化作用[23]。从生物学角度看,CO2主要由微生物氧化有机物和根系呼吸产生,影响这两个组分的主导环境因子不尽相同,任何一个组分的呼吸过程改变都会使总的土壤呼吸速率发生变化,因此影响土壤微生物活动的诸因子,如土壤有机质含量、pH、温度、水分以及有效养分含量都能影响土壤呼吸作用强度,并从土壤呼吸作用强度的变化中反映出来。此外,影响根系呼吸的诸多因子:土壤质地、风速等环境因子也均会改变土壤CO2的排放速度。
2.1 不同植被类型土壤呼吸速率日变化
从同一坡向、坡度,不同植被类型5—8月土壤呼吸速率日变化观测数据(图 1)可知:草地和林地5—8 月土壤呼吸速率日变化曲线趋势基本相近,总体上均表现为单峰曲线,且5—8 月两种植被类型土壤呼吸速率曲线波峰均出现在17:00—18:00,波谷出现在 9:00—10:00。但在日变化过程中,由于受到研究区地理位置的影响,土壤呼吸速率出现日最高值的时段与其他林地有所差异。谭炯锐等[24]报道北京大兴杨树人工林土壤呼吸速率日变化最大值出现在10:30—13:00;周海霞[25]等研究发现东北温带落叶松人工林土壤表面CO2排放通量的昼夜变化呈单峰状态,最大值出现在 19:00—22:00。但本研究结果与史宝库等[26]对小兴安岭5种林型土壤呼吸时空变异的研究结果相一致。
草地土壤呼吸速率日变化曲线波峰最大值出现在7月为1.55 µmol/(m2·s),8月次之为1.36 µmol/(m2·s),5月和6月相差不大分别为1.13和1.14 µmol/(m2·s);林地土壤呼吸速率日变化曲线波峰最大值也出现在7月为2.43 µmol/(m2·s),8月、5月、6月逐渐减小,6月最小仅1.76 µmol/(m2·s),但仍比整个观测时间段草地出现的波峰最大值高0.21 µmol/(m2·s)。
草地土壤呼吸速率日变化曲线波谷的最大值也出现在7月为0.8 µmol/(m2·s),8月、5月、6月逐渐减小,最小值出现在6月为0.35 µmol/(m2·s);林地土壤呼吸速率日变化曲线波谷的最大值也出现在 7月为1.28 µmol/(m2·s),8月、6月相差不大,5月最小,其呼吸速率仅0.71 µmol/(m2·s)。
从5—8月各月土壤呼吸速率日平均值看,草地从大到小依次为7月、8月、5月、6月,林地从大到小依次为7月、8月、6月、5月,且整个观测时间林地土壤呼吸速率日平均值最小值大于草地土壤呼吸速率日平均值最大值,通过配对样本T检验,发现所得观测值具有明显差异(P<0.01)。这是因为林地内的西伯利亚落叶松长期落叶使林地内土壤有机质含量高于草地,致使林地土壤呼吸速率值明显大于草地。另外,7月同一坡向、坡度,草地和林地土壤呼吸速率明显高于其他3个月(P<0.01),主要是该地区7月气温明显高于其他3个月造成的。速率的主要因子,但不是唯一的控制因子,它主要是通过对土壤微生物代谢和植物根系生长的影响来调控土壤呼吸作用。在本研究中,土壤CO2释放的日变化趋势与土壤温度变化趋势具有一定的一致性(图2和图3)。
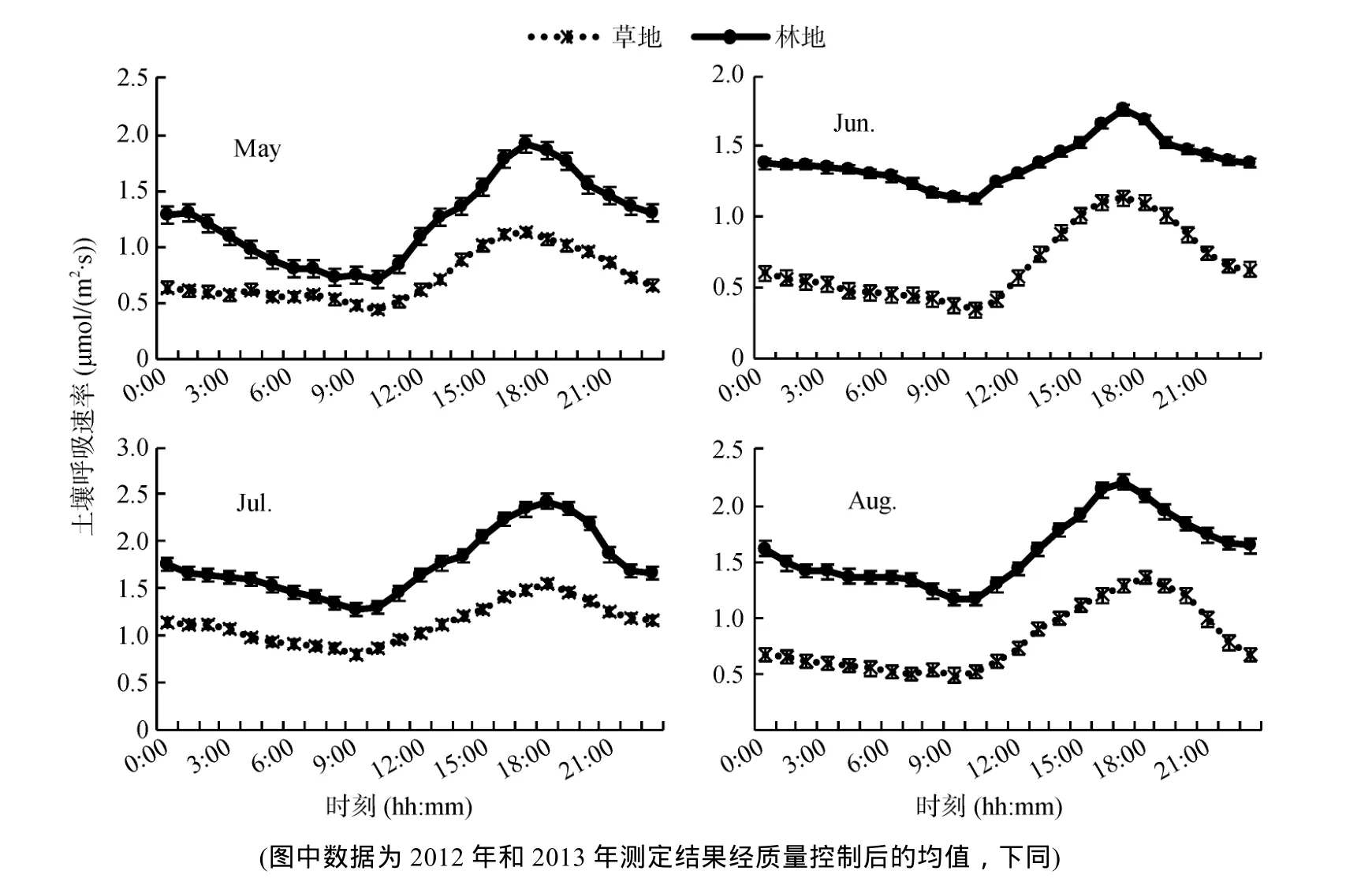
图1 林地、草地土壤5—8月土壤呼吸速率日变化Fig. 1 Daily changes of soil respiration rates under forest and grassland ecosystems from May to August

图2 草地5—8月土壤温度日变化Fig. 2 Daily changes of soil temperature under grassland ecosystem from May to August

图3 林地5—8月土壤温度日变化Fig. 3 Daily changes of soil temperature under forest ecosystem from May to August
图2和图3显示,同一坡向、坡度,草地和林地5—8月0 ~ 5 cm土层土壤平均温度昼夜变化曲线在总体上均呈单峰曲线型,波峰均出现在18:00左右,波谷均出现在09:00左右。由相关分析可知(表1),与草地、林地0 ~ 5 cm土层平均温度与土壤呼吸速率变化曲线相近,两者间具有显著的正相关性(P<0.05),且 R2值均有较大值,综合说明土壤 CO2释放的昼夜变化趋势与土壤温度变化趋势具有高度一致性。这与周小刚等[27]发现的土壤温度能够很好地解释农田和草地生态系统之间土壤呼吸的差异,及代快等[28]在研究旱作春玉米农田土壤温度对土壤呼吸速率的影响时,得到的月动态峰值出现在7月,通过耕作措施改变土壤温度是影响旱作农田 CO2排放的重要因素等结果并无矛盾。
从草地0 ~ 5 cm土层土壤温度与土壤呼吸速率日变化趋势(图 1、图 2)可以发现:日出后随土壤温度的快速上升,及18:00后土壤温度的缓慢下降,土壤呼吸速率在相应时段也呈相同的变化趋势,两者的最大值在同一时段出现,但是最小值并不同步,5月和6月先与土壤呼吸速率出现最小值,到7月同时出现,至8月稍晚出现最小值。这可能与不同月份日出时间不同,及草地0 ~ 5 cm 土层湿度的不同有关。对土壤呼吸速率与0 ~ 5 cm土层平均温度进行拟合,发现线性和指数模型能较好地描述两者之间的关系,各月指数模型拟合的决定系数 R2分别为 0.867、0.963、0.879、0.832,均达到极显著水平(P<0.01)。
从林地0 ~ 5 cm土层土壤温度与土壤呼吸速率日变化趋势(图1、图3)及二两者的回归关系(表1)可以发现,林地土壤0 ~ 5 cm土层平均温度日变化趋势与土壤呼吸速率变化趋势有较好的一致性,都是日出前最低,日出后迅速上升至18:00左右达最高;但两者的整体极值出现时刻不一致,土温的最小值只有7月与土壤呼吸速率同时刻出现,其他都比土壤呼吸速率早1 h出现;最大值除6月土温早1 h出现,其他基本同时出现。谭炯锐等[24]研究表明,北京大兴杨树人工林土壤温度是影响土壤呼吸速率的主要因素;丁金枝等[29]和刘颖[30]等报道不同植被的土壤呼吸速率与土壤温度之间呈显著的指数相关。这些与本研究结果相一致。从二者的指数模型(表 1)可以看出,林地土壤呼吸速率日动态变化和0 ~ 5 cm表层土壤温度的昼夜波动趋势较为吻合,且两者间具有显著的正相关性(P < 0.01),但林地各月R2除8月外均小于草地,通过图2、图3及表1可以发现林地下垫面对0 ~ 5 cm表层土壤温度的调温效果好于草地。
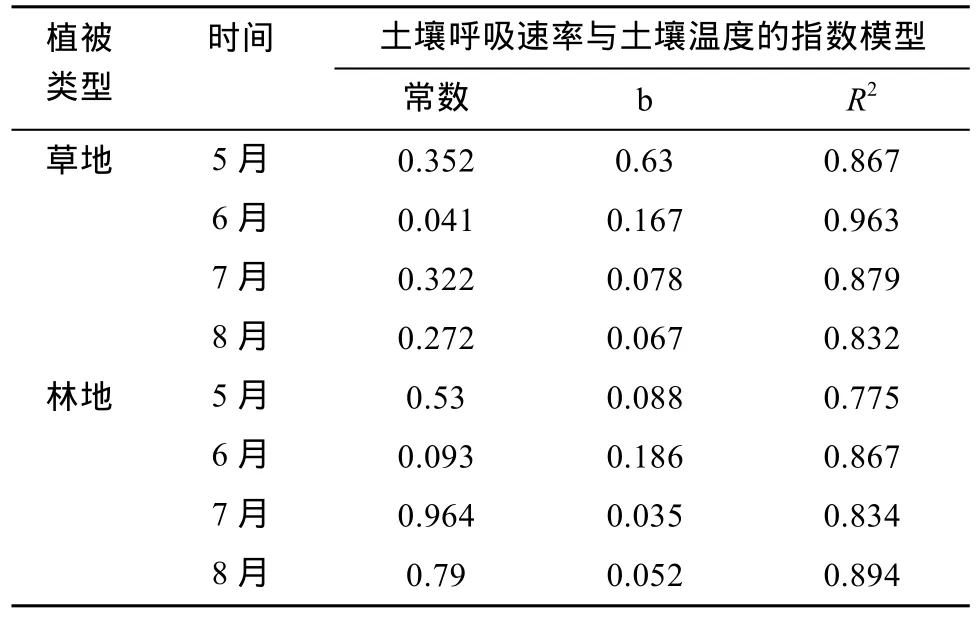
表1 土壤呼吸速率与0 ~ 5 cm土层温度之间的函数关系Table 1 Functional relations between soil respiration rates and 0–5 cm soil temperatures under forest and grassland ecosystems
2.3 不同植被类型土壤含水量日变化
土壤水分是影响陆地生态系统 CO2通量的重要环境要素,对植被的生长、根系分布、微生物活性等与土壤呼吸密切相关的生物因子起控制作用[7]。诸多研究显示,水分对土壤呼吸的影响具有复杂性与不确定性,不同生态系统土壤水分与呼吸间分别存在正相关、负相关及不相关关系:如马骏和唐海萍[31]在内蒙古农牧交错区研究不同土地利用方式下土壤呼吸速率变化时,发现土壤含水量是影响土壤呼吸速率的另一个重要因素,但土壤含水量与土壤呼吸速率间的相关性不显著,土壤含水量不能解释土壤呼吸速率的变化。王春燕等[32]结果显示,橡胶林土壤呼吸速率及其土壤湿度的相关性不显著,土壤水分与土壤呼吸速率没有直接的关系。陈骥等[33]在青海湖北岸研究高寒草甸草原非生长季的土壤呼吸对温度和湿度的响应中,得出非生长季土壤温度和土壤湿度对土壤呼吸的解释率相当,一定的土壤含水量是保证土壤呼吸对温度响应的必要条件,并且土壤湿度对土壤呼吸的解释率较土壤温度更高。龚斌等[34]研究发现,各个森林类型的土壤呼吸作用与土壤湿度的相关性不大,究其原因主要是降雨量过大造成土壤孔隙度变小,影响了土壤呼吸作用的活跃度。
由图4可知,在本研究中5—8月,草地、林地0 ~ 5 cm土层土壤平均含水量变化趋势除7月稍有浮动外,其他各月变化均比较平缓,没有明显的日变化,且各时期草地土壤平均含水量为 5月>8月>6月>7月,林地土壤平均含水量为5月>6月>8月>7月。这可能与草地土壤表面在 5月上旬左右积雪已全部融化,而林地土壤表面在5月下旬仍有部分积雪未融化有关。相关分析显示:草地、林地土壤土壤含水量与土壤呼吸速率两者间相关性不明显。
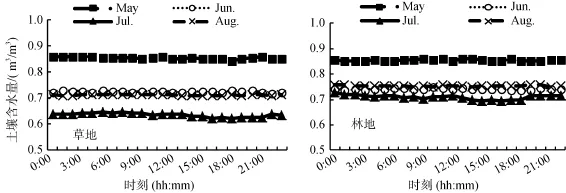
3 结论
1) 喀纳斯自然保护区林地和草地两种植被类型土壤呼吸速率的日变化和季节变化均呈单峰曲线,其日最大值出现在北京时间 16:00—18:00,月最大值出现在7月。不同植被类型土壤在相同的植被生长季内土壤呼吸速率之间存在差异,研究期内林地土壤呼吸速率最大值为2.43 µmol/(m2·s),林地的土壤呼吸速率显著高于草地土壤呼吸速率最大值(1.55 µmol/(m2·s))。
2) 研究区草地和林地两种植被土壤CO2释放的日变化趋势与0 ~ 5 cm土层土壤平均温度变化趋势具有极显著相关性,呈单峰抛物线型,峰值均出现在18:00左右,最低值均出现在10:00左右。5—8月,草地和林地土壤温度的月动态峰值分别出现在 6月和7月。
3) 研究区草地和林地土壤温度和土壤含水量对土壤呼吸的影响是同时进行的,但土壤呼吸速率在植被生长季的变化趋势大体与土壤温度相似,土壤含水量没有类似的趋势。根据相关分析可知,草地、林地土壤土壤含水量与土壤呼吸速率两者间相关性不明显。
[1] 刁一伟, 郑循华, 王跃思, 等. 开放式空气CO2浓度增高条件下旱地土壤气体CO2浓度廓线测定[J]. 应用生态学报, 2002, 13(10): 1249–1252
[2] Schlesinger W H, Andrews JA. Soil respiration and the global carbon cycle[J]. Biogeochemistry, 2000, 43: 7–20
[3] 黄湘, 李卫红, 陈亚宁, 等. 塔里木河下游荒漠河岸林群落土壤呼吸及其影响因子[J]. 生态学报, 2007, 27(5): 1951–1959
[4] 米迎宾, 杨劲松, 姚荣江, 等. 不同措施对滨海盐渍土壤呼吸、电导率和有机碳的影响[J]. 土壤学报, 2016, 53(3): 437–441
[5] 康熙龙, 张旭辉, 张硕硕, 等. 旱地土壤施用生物质炭的后效应——水分条件对土壤有机碳矿化影响[J]. 土壤, 2016, 48(1): 145–150
[6] Raich J W, Potter C S. Global patterns of carbon dioxide emissions from soils[J].Global Biogeochemical Cycles, 1994, 9: 23–36
[7] Rustad L E, Huntington T G, Boone R D. Controls on soil respiration:implications for climate change[J]. Biogeochemistry, 2000, 48: 1–6
[8] 高艳红, 刘立超, 贾荣亮, 等. 沙坡头人工植被演替过程的土壤呼吸特征[J]. 生态学报, 2012, 32(8): 2474–2482
[9] 孟祥利, 陈世平, 魏龙, 等. 库布齐沙漠油蒿灌丛土壤呼吸速率时空变异特征研究[J]. 环境科学, 2009, 30(4): 1152–1158
[10] Raich J W, Schlesinger W H. The global carbon-dioxide flux in soil respiration and its relationship to vegetation and climate[J]. TellusB,1992, 44(2): 81–99
[11] 潘文斗. 中国喀纳斯保护区科学考察[M]. 北京: 中国林业出版社, 1989: 4–7
[12] 叶高, 刘华, 白志强, 等. 喀纳斯自然保护区3种天然林分土壤呼吸速率的动态变化[J]. 东北林业大学学报, 2014, 42(3): 77–84
[13] 赖晶晶, 张世熔, 刘艳婷, 等. 基于多元分析的岷江下游土壤有机碳密度空间分布及影响因素研究[J]. 土壤, 2016, 48(1): 151–156
[14] Zhou W J, Wang K R, Zhang Y Z, et al. Phosphorus transfer and distribution in a Soybean-citrus inter-cropping system[J]. Pedosphere, 2006,16(4): 435–443
[15] Liu X Y, Yang Z P, Ma Z G, et al. Land use and land cover pattern in Kanas Natural Reserve based on terrain factors. Arid Land Geography, 2011, 5(34): 851–857
[16] 贺明荣, 冷寿慈. 桃粮间作对土壤养分状况及土壤生物活性的影响[J]. 土壤通报, 1994, 25(4): 188–189
[17] 潘晓玲, 张宏达. 哈纳斯自然保护区植被特点及植物区系形成的探讨[J]. 干旱区资源与环境, 2011, 25(10): 92–97
[18] 李偲, 海米提×依米提,李晓东. 喀纳斯自然保护区森林生态系统服务功能价值评估[J]. 东北林业大学学报, 2003, 31(6): 102–103
[19] 刘新宇, 杨兆萍, 马忠国, 等. 基于地形因子的喀纳斯自然保护区土地利用/覆被现状格局分析[J]. 干旱区地理, 2011, 5(34): 851–857
[20] 张荟荟, 潘存德, 巴扎尔别克·阿斯勒汗, 等. 喀纳斯旅游区森林群落类型及其物种多样性随海拔梯度的变化[J]. 新疆农业科学, 2008, 45(2): 225–231
[21] 刘翠玲, 潘存德, 吴晓勇, 等. 新疆喀纳斯树种丰富度垂直格局特征[J]. 生态学杂志, 2009, 28(2): 203–208
[22] 陈全胜, 李凌浩, 韩兴国, 等. 水分对土壤呼吸的影响及机理[J]. 生态学报, 2003, 23(5): 972–978
[23] 韩兴国, 李凌浩, 黄建辉. 生物地球化学概论[M]. 北京:高等教育出版社, 1999
[24] 谭炯锐, 查同刚, 张志强, 等. 土壤温湿度对北京大兴杨树人工林土壤呼吸的影响[J]. 生态环境学报, 2009, 18(6): 2308–2315
[25] 周海霞, 张彦东, 孙海龙, 等. 东北温带次生林与落叶松人工林的土壤呼吸[J]. 应用生态学报, 2007, 18(12): 2668–2674
[26] 史宝库, 金光泽, 汪兆洋. 小兴安岭 5 种林型土壤呼吸时空变异[J]. 生态学报, 2012, 32(17): 5416–5428
[27] 周小刚, 张彦军, 南雅芳, 等. 黄土区农田和草地生态系统土壤呼吸差异及其影响因素[J]. 环境科学, 2013, 34(3): 1026–1033
[28] 代快, 蔡典雄, 王燕, 等. 不同耕作措施对旱作春玉米农田土壤呼吸影响的研究——土壤温度对土壤呼吸速率的影响[J].中国土壤与肥料, 2010(6): 64–69
[29] 丁金枝, 来利明, 赵学春, 等. 荒漠化对毛乌素沙地土壤呼吸及生态系统碳固持的影响[J]. 生态学报, 2011, 31(6): 1594–1603
[30] 刘颖, 韩士杰, 胡艳玲, 等. 土壤温度和湿度对长白松林土壤呼吸速率的影响[J]. 应用生态学报, 2005, 16(9): 1581–1585
[31] 马骏, 唐海萍. 内蒙古农牧交错区不同土地利用方式下土壤呼吸速率及其温度敏感性变化[J]. 植物生态学报, 2011, 35(2): 167–175
[32] 王春燕, 陈秋波, 袁坤, 等. 橡胶林土壤呼吸速率及其与土壤温湿度的关系[J]. 土壤学报, 2013, 50(5): 974–982
[33] 陈骥, 曹军骥, 魏永林, 等. 青海湖北岸高寒草甸草原非生长季土壤呼吸对温度和湿度的响应[J]. 草业学报, 2014, 23(6): 78–86
[34] 龚斌, 王风玉, 张继平, 等. 中亚热带森林土壤呼吸日变化及其与土壤温湿度的关系[J]. 生态环境学报, 2013, 22(8): 1275–1281
Differences in Soil Respiration Between Forestland and Grassland Ecosystems and Influential Factors in Kanas Nature Reserve
FAN Zi’ang1,2, DOU Xiaojing3, ZOU Chen2*, JI Chunrong2, HUANG Youzhi4
(1 College of Environmental Science & Engineering, Nanjing University of Information Science & Technology, Nanjing 210044, China; 2 Institute of Desert Meteorology, China Meteorological Administration, Key Laboratory of Tree-ring Physical Chemic Research of China Meteorological Administration, Key Laboratory of Tree-ring Ecology of Uigur Autonomous Region, Urumqi 830002, China; 3 Institute of Soil and Fertilizer & Agricultural Sparing Water, Xinjiang Academy of Agricultural Sciences, Urumqi 830091, China; 4 Buerjin Meteorological Bureau, Buerjin, Xinjiang 836600, China)
In order to disclose the interaction between forest and the grassland ecosystems with atmosphere in Kanas National Nature Reserve in Xinjiang and the spatial and temporal variation of soil respiration rate (SRR) as well as the influential factors, LI - 8150 soil carbon flux measurement system was used to continuously and automatically monitor daily soil respiration rates under the two vegetation ecosystems in the growing season. The results showed that the diurnal variation of SRR under both forest and grassland ecosystems in the growing season presented obvious unimodal type curve, the monthly maxima of SRR both appeared in July, maximum of SRR under forest ecosystem (2.43 µmol/(m2·s)) was significantly higher than that under grass ecosystem (1.55 µmol/(m2·s)). Monthly SRR under forest ecosystem was obviously higher than that under grass ecosystem. The peak and bottom of SRRs occurred at 17:00–18:00 and at 9:00–10:00, respectively. Minimum of SRRs occurred in June under grass ecosystem (0.35 µmol/(m2·s)) and in May under forest ecosystem (0.71 µmol/(m2·s)). SRR was higher during the day than in the evening. CO2was the main exhalant ingredient throughout the day. SRRs of woodland and grassland ecosystems had similar change trend and significant positive correlation with soil temperature in the growing season, but not with soil moisture content.
Kanas Nature Reserve; Soil respiration rate; Soil temperature; Soil moisture
S154.4
A
10.13758/j.cnki.tr.2017.01.028
国家自然科学基金项目(41675152)、中央公益性基本科研业务费项目(IDM2016006)和公益性行业(气象)科研专项(GYHY(QX) 201506001)资助。
* 通讯作者(zoucheng@idm.cn)
范子昂(1982—),男,福建上杭人,助理研究员,主要从事树木年轮与气候变化方面研究。E-mail: franz1326@163.com
猜你喜欢
小读者(2020年4期)2020-06-16 03:33:52
金桥(2019年12期)2019-08-13 07:16:34
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
中国三峡(2017年1期)2017-06-09 11:09:41
河北林业科技(2016年5期)2016-11-08 03:13:26
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
学生天地(2016年10期)2016-04-16 05:14:44
大气科学(2015年5期)2015-12-04 03:04:44
中国土地科学(2014年5期)2014-03-01 03:25:40
