
表层水温结构变化对东南太平洋秘鲁鳀渔场的影响
2017-04-14 09:16:50陈芃陈新军
海洋学报 2017年4期
陈芃,陈新军
(1. 上海海洋大学 海洋科学学院,上海 201306;2.上海海洋大学 大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306;3.上海海洋大学 国家远洋渔业工程技术研究中心,上海 201306;4.远洋渔业协同创新中心,上海 201306)
表层水温结构变化对东南太平洋秘鲁鳀渔场的影响
陈芃1,4,陈新军1,2,3,4*
(1. 上海海洋大学 海洋科学学院,上海 201306;2.上海海洋大学 大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306;3.上海海洋大学 国家远洋渔业工程技术研究中心,上海 201306;4.远洋渔业协同创新中心,上海 201306)
秘鲁鳀(Engraulisringens)是栖息于东南太平洋沿岸的小型中上层鱼类,掌握其渔场变化及其与海洋环境因子的关系有利于企业把握该渔业的生产情况。研究结合2005-2014年渔汛期间秘鲁各港口出港的船数及其所获得的秘鲁鳀渔获量和表层水温数据(海表面温度,sea surface temperature,SST),以单位捕捞努力量的渔获量(catch per unit effort,CPUE)和捕捞努力量(Effort)数据构建的渔场指数(fishing ground index,FGI)为渔场指标,对秘鲁鳀的渔场类型进行分类,探讨水温结构变化对秘鲁鳀渔场的影响。方差分析表明:渔场指数在不同渔汛阶段(渔汛前期、中期和末期,P<0.01)和不同捕捞区域(北部、中部和南部,P<0.01)都有着极显著的差异。以表层水温大于20℃的海水是否入侵到近岸为标志,可以将秘鲁鳀渔场分成两种类型:大于20℃的海水没有入侵到沿岸(A型渔场)和入侵到沿岸(B型渔场)。研究表明,在渔汛前期和中期阶段,A型渔场的出现有利于渔场的形成,其中渔汛前期沿岸19℃或20℃等温线的出现以及渔汛中期沿岸18℃或19℃等温线的出现可以作为中心渔场形成的指标。
秘鲁鳀;东南太平洋;表层水温结构;渔场
1 引言
秘鲁鳀(Engraulisringens)是一种重要的经济型鱼类,栖息于东南太平洋近岸,其渔业为世界上产量最大的单鱼种渔业[1—2]。主要的捕捞国家为秘鲁和智利,其中秘鲁为首要的捕捞国家,2000年以来其产量占到了世界总产量的80%以上[3]。由于其近岸分布的特性,在秘鲁境内一般仅由本国的船队进行捕捞作业[1—2,4]。我国是世界上最大的鱼粉进口国,秘鲁鳀由于其优良的质量在我国的进口鱼粉的比例最高[5],而秘鲁鳀的产量波动直接影响到了鱼粉市场的状况[6—7],因此,结合海洋环境情况分析秘鲁鳀的渔场情况有助于为我国鱼粉进口企业提供技术支持。国内外学者对秘鲁鳀资源渔场与海洋环境的关系作了许多研究,例如:Yez等[8]分析了资源丰度与海域上升流指数与湍流系数的关系;iquen等[9]探究了厄尔尼诺及拉尼娜事件对其资源的影响;Gibson等[10]研究了海域氧含量与秘鲁鳀离岸远近的关系。但是这些环境因子较难被实时监测和获取,而海表面温度(sea surface temperature,SST)作为最易获取的卫星遥感产品,监测技术也最为成熟,已在诸多渔场分析中得到成功应用[11—13]。目前在秘鲁沿岸水温状况与秘鲁鳀的渔场分析方面,主要由秘鲁国家海洋局(Instituto del Mar del Perú,IMARPE)在其网站上(http://www.imarpe.pe/imarpe2/)发布短期的渔场总结报告,其中表层水温与渔场的关系主要依靠于历史的经验总结为主,并没有具体量化的研究。对于秘鲁鳀渔场,沿岸表层水温结构变化是如何影响其渔场变动的?可否找出关键的等温线作为中心渔场的指标?为此,研究结合2005-2014年渔汛期间秘鲁各港口出港的船数及所获得的秘鲁鳀渔获量和表层水温(SST)数据,引入渔场指数(fishing ground index,FGI)的概念,分析水温结构变化对秘鲁鳀渔场的影响,以期为我国鱼粉进口企业把握该渔业生产情况提供技术支撑。
2 材料与方法
2.1 数据来源
秘鲁鳀的生产数据来源于秘鲁国家海洋局IMARPE网站(http://www.imarpe.pe/imarpe2),图1为2005-2014年秘鲁各港口(8°~15°S)渔汛期间(4-8月)每日出港的大型工业围网渔船的总船数及其所获得的渔获量(数据未包含每条渔船作业的具体位置)。SST数据来源于美国国家海洋大气局NOAA的OceanWatch网站 (http://oceanwatch.pifsc.noaa.gov/las/servlets/dataset),时间分辨率均为周,空间分辨率分别为0.1°×0.1°。空间范围为7°~15°S,75°~85°W。
2.2 分析方法
2.2.1 渔场分析
(1)捕捞区域和渔汛阶段划分
秘鲁鳀主要栖息于沿岸30海里50 m水深内的海域[8],同时渔船都为当日出海捕捞当日回港[14],捕捞位置离港口不远,渔场为近岸渔场。因此研究主要探索近岸水温结构对渔场的影响,以港口的位置作为渔场的位置。同时,渔场空间上南北分布的不同可能会导致渔场间存在差异。为此,从8.5°S纬度线开始,向南每隔2°纬度线划分捕捞区域,分成北部(8.5°~10.5°S)、中部(10.5°~12.5°S)和南部(12.5°~14.5°S)3块捕捞区域,这3块区域包含了研究中所有的港口(图1)。
时间上,以每年4月1日所在周数为第一周,渔汛总共可以持续18~19周。研究同时探究渔场时间上的差异:以1~6周为渔汛前期;7~12周为渔汛中期;13~19周为渔汛末期,分析各个时期的渔场变化。
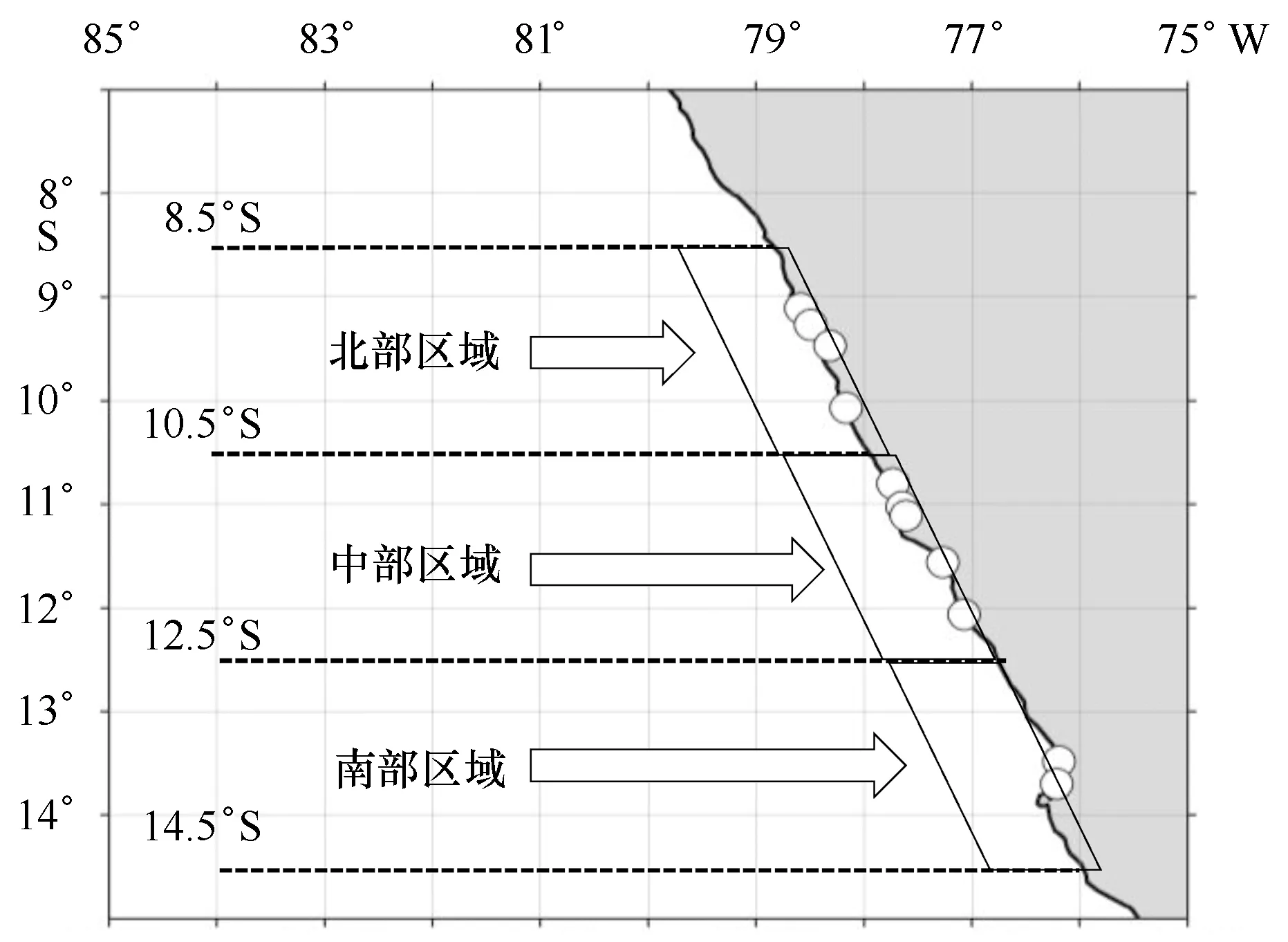
图1 秘鲁鳀港口分布及捕捞区域划分Fig.1 Port distributions in Peruvian coast and the classification to fishing areas白色圆点代表港口的位置,从上至下分别是:钦博特、萨曼库、卡斯马、瓦尔梅、苏普、维格塔、瓦乔、昌凯、卡亚俄、坦博德莫拉和皮斯科White dots represent the position of port from up to down: Chim-bote, Samanco, Casma, Huarmey, Supe, Vegueta, Huacho, Chancay, Callao, Tambo de Mora and Pisco
(2)渔场指数
根据渔场的定义[15],渔场必须满足两个特性:较高的渔业资源丰度(fishery abundance)和较好的作业可行性(fishery availability)。这两个指标通常使用单位捕捞努力量的渔获量(catch per unit effort,CPUE)和捕捞努力量(Effort)来表示[16-17]。但是以往的研究中,往往只选取单一的因子进行分析。这经常会出现以下两种情况:第一,CPUE很高但是捕捞努力量较低,这可能是由于海况条件不好不利于作业因此海域不具有良好的作业可行性;第二,捕捞努力量很高但是CPUE较低,这来源于虽然海域资源情况不好,但是渔民或船队可能为了收回成本或者完成捕捞计划聚集在某个区域从事捕捞作业。此外,研究中经常只使用名义的CPUE或者名义的捕捞努力量进行研究,很大的一个原因就在于数据的缺乏而无法进行标准化。但是即使是名义的CPUE或者是名义的捕捞努力量,它们都在一定程度上反映了作业海域的资源丰度和作业可行性,因此笔者认为,为了能够相对真实地反映渔场的状况,可以结合CPUE和捕捞努力量构造渔场指数来更为真实地反映渔场状况。为此引入渔场指数的概念,公式如下:
(1)
式中,Effort和CPUE为一周某个捕捞区域内所有港口的总出港船数和单位捕捞努力量的渔获量(总渔获量除以总出港船数),CPUEmax和Effortmax为当年CPUE和Effort的最大值。渔场指数的值在0~1之间,其值越接近于1表示渔场情况越好。
(3)方差分析和多重比较
使用方差分析[18]来评价渔场时间和空间上的差异性,即验证了渔汛阶段和捕捞区域划分的可行性。方差分析之前,利用Levene检验[19]检验渔场指数的方差是否齐性。使用最小显著差数法(least significant difference method,LSD)[18]对方差分析结果进行多重比较,以探究不同捕捞区域和不同渔汛阶段间渔场指数的差异。数据分析使用SPSS20.0软件。
2.2.2 分析渔场与水温结构关系
利用Arcgis10.2作出沿岸水温结构图,等温线插值使用Arcgis10.2中的Contour工具。观察并描述沿岸等温线分布情况,以水温结构不同归纳不同海况类型的渔场。使用t检验[18]来探究不同类型渔场间渔场指数的差异性。
可以看出,渔场指数与前人的栖息地指数对渔场的研究类似,因此,参照前人的研究,将渔场指数大于0.6作为中心渔场的指标[20],对比中心渔场和等温线出现情况的关系,以寻找能代表中心渔场的关键等温线。
3 结果
3.1 渔场时空差异分析
Levene检验表明,渔场指数方差是齐性的(F=1.57,df1=8,df2=290,P=0.13),因此可以进行方差分析。方差分析表明:渔场指数在不同渔汛阶段(F=1 273.97,df1=3,df2=294,P<0.01)和不同捕捞区域(F=50.26,df1=2,df2=294,P<0.01)都有着极显著的差异。最小显著差异法比较表明(表1):时间上,渔汛前期的渔场指数要极显著地高于中期和后期(P<0.01),同时渔汛中期阶段的渔场指数也要极显著地高于后期(P<0.01);空间上,北部区域的渔场指数要显著(P<0.05)和极显著(P<0.01)地低于中部和南部区域,而中部区域的渔场指数也要显著地低于南部区域(P<0.05),这同时也验证了捕捞区域和渔汛阶段划分的可行性。从图2也可以看出:时间上,3块区域随着时间的推移,渔场指数都在变小;空间上,除了末期的中部区域的渔场指数稍小于北部以外,各渔汛阶段北部渔场的渔场指数要低于中部和南部区域,而渔汛前期和中期中部区域和南部区域的渔场指数基本相等,到了渔汛末期南部区域的渔场指数要大于中部。
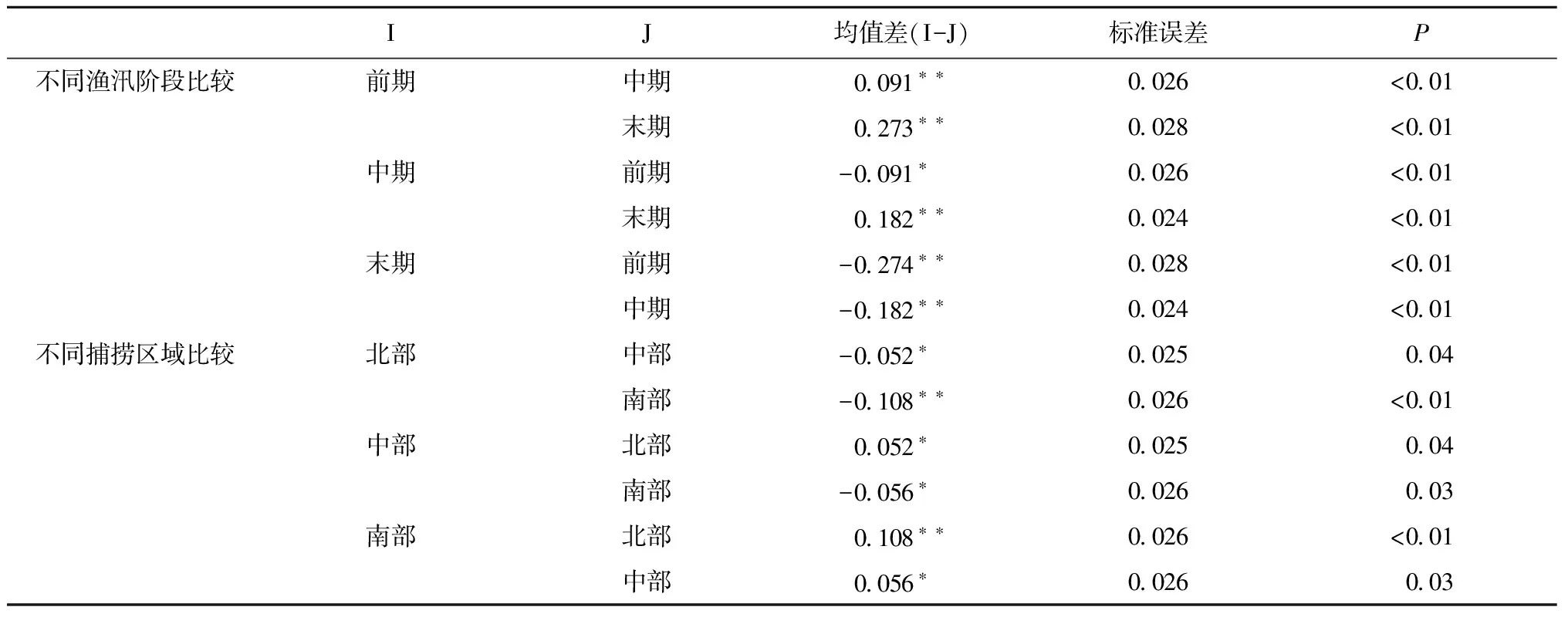
表1 不同渔汛阶段和不同区域间秘鲁鳀渔场指数最小显著差异法比较结果
注: *表示在0.05显著性水平上差异显著;**表示在0.01显著性水平上差异极显著。
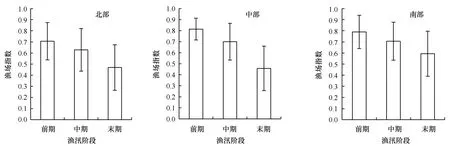
图2 秘鲁鳀渔场指数变化状况Fig.2 Variations of anchoveta (Engraulis ringens) fishing ground index图中误差线表示标准差 The error bars in the picture represent the standard deviation
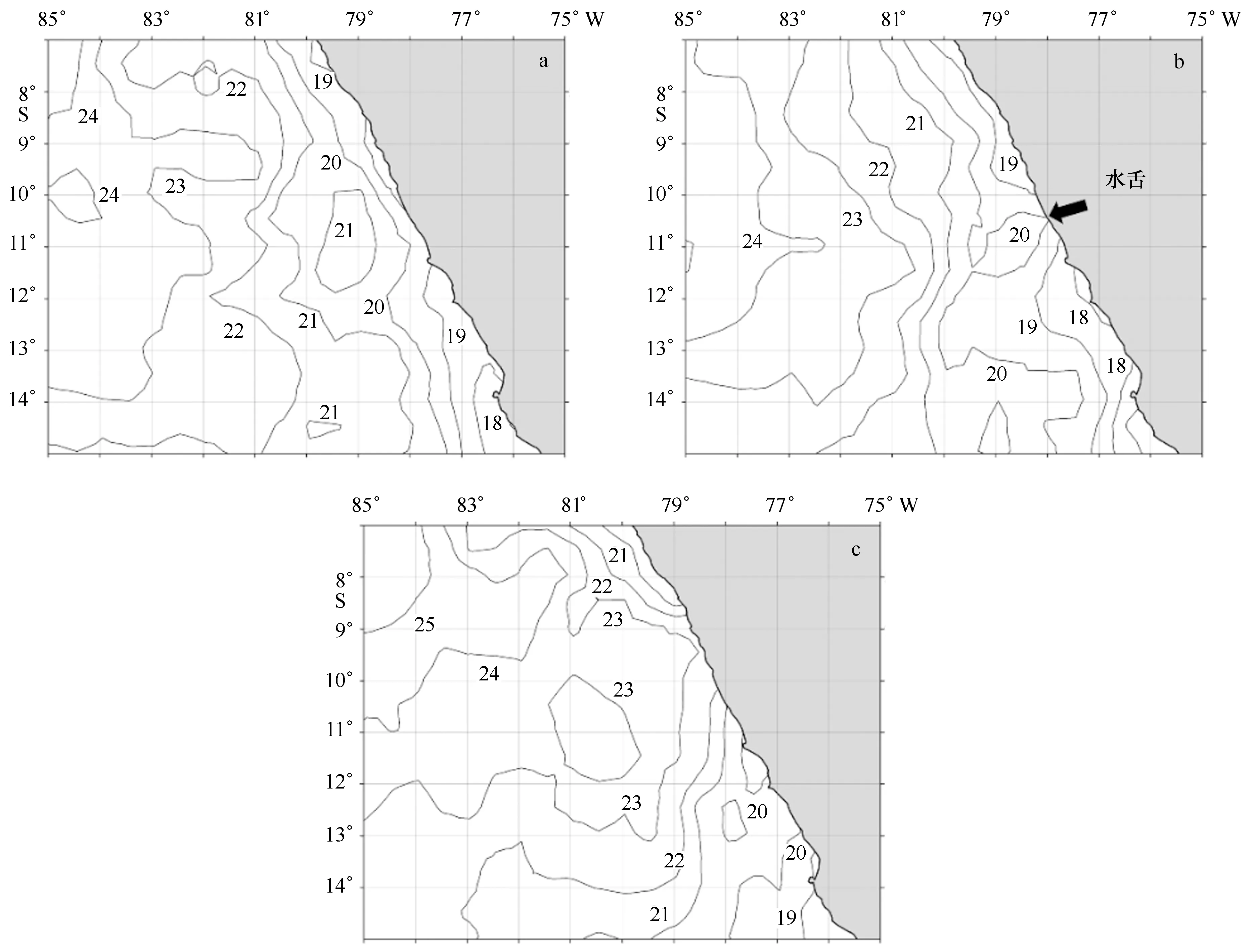
图3 秘鲁沿岸表层水温(单位:℃)结构典型范例(图中等温线经过平滑处理)Fig.3 Typical examples of the surface temperature(unit:℃) structure along the Peruvian coastal area (the isotherms in the pictures are after smoothing process)
3.2 秘鲁鳀渔场类型划分
正常情况下(图3a),秘鲁沿岸水温由近岸向外海升高;以20℃等温线为冷暖区域的边界,可以看出,外海是广阔的暖水水域,而近岸则是狭长的冷水水域,冷水区域的等温线通常与沿岸的走向保持相同(如图3a中的20℃和19℃等温线以及图3b中的19℃和18℃等温线)。暖水区域能以两种方式入侵到岸界,第一种:以水舌(water meander,图3b)的方式向近岸入侵;第二种:整体向东部移动,沿岸的冷水区域不存在。
因此,以暖水是否入侵岸界为标准,通过判断沿岸水温结构确定沿岸附近的温度情况,对北部、中部和南部的秘鲁鳀渔场划分为以下两种类型(图4)。
A型渔场:沿岸表层水温小于20℃区域的南北跨度(L<20℃)大于1°纬度。
B型渔场:沿岸表层水温大于20℃区域的南北跨度(L>20℃)大于1°纬度。
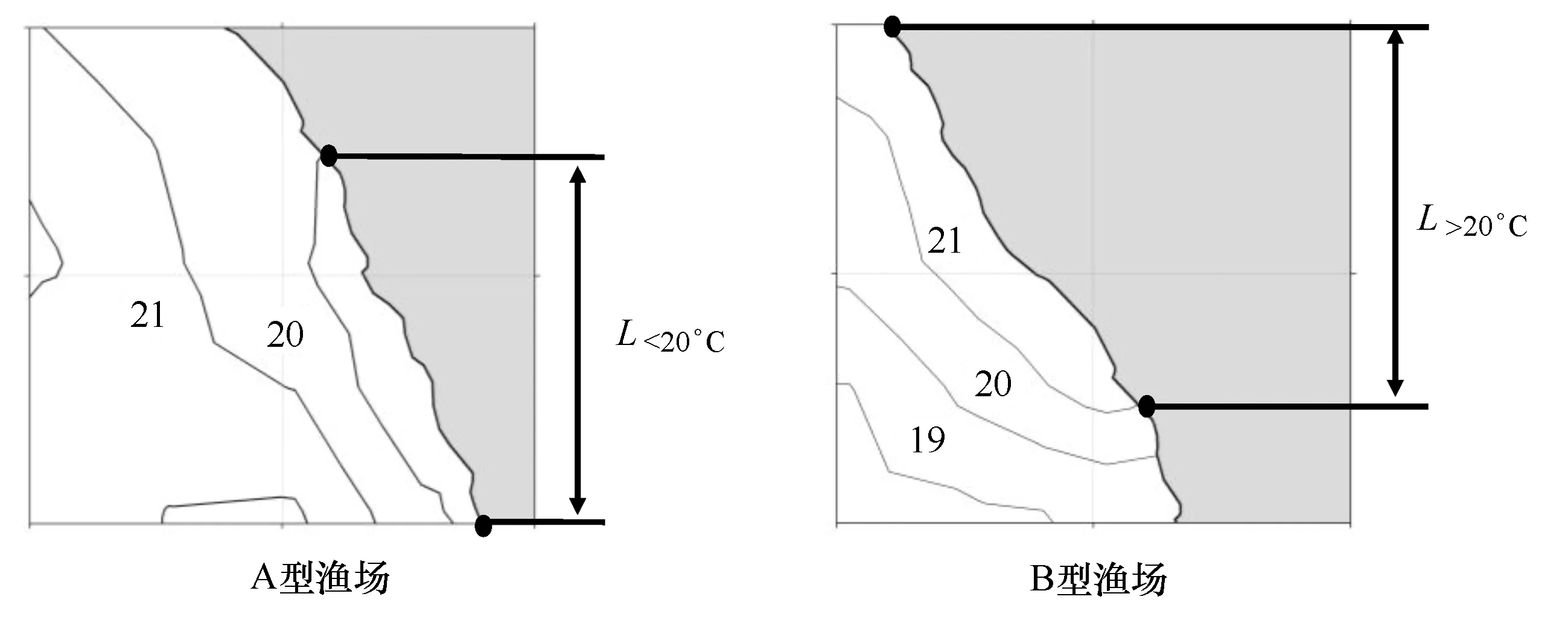
图4 基于表层水温(℃)结构的秘鲁鳀渔场类型划分Fig.4 Classification to the fishing ground of anchoveta (Engraulis ringens) based on the surface temperature (℃) structure
3.3 不同渔场类型与渔场指数的关系
在渔汛前期,中部和南部区域完全为A型渔场(100%),北部区域中绝大多数渔场类型也为A型渔场(91.7%);而在渔汛中期,3个区域内的A型渔场的比例都出现了下降;到了渔汛末期,除了南部区域A型渔场的比例略有升高以外,其他两个区域A型渔场比例依然呈现下降的趋势(图5)。

图5 不同渔汛阶段和不同区域间A型渔场所占比例Fig.5 Percentage of the “A” type fishing ground in different fishing periods and fishing areas
由于渔汛前期北部渔场中B型渔场仅出现过两次,数量过小不具有代表性,因此只对中期和末期各区域的渔场指数进行分析:t检验表明,在渔汛中期,3个区域的A型渔场的渔场指数均要显著地大于B型渔场(P<0.05);在渔汛末期,3个区域的A型渔场的渔场指数与B型渔场均不存在显著的差异(P>0.05)。
3.4 A型渔场水温分布与渔场指数的关系
对A型渔场的水温分布做进一步分析(图6):判断沿岸水温情况,同样以沿岸小于某个温度(20℃、19℃、18℃、17℃和小于17℃)区域的南北跨度大于1°纬度为标准,取最小温度,认为该区域受到这个等温线的控制。统计不同等温线控制下的各捕捞区域和渔讯阶段的中心渔场(渔场指数大于0.6)占所有A型渔场比例,结果如下(图7):北部区域,在渔汛前期、中期和末期中心渔场占所有A型渔场的比例分别为72.7%、69.2%和21.4%,其中,前期受到19℃和20℃等温线控制的区域比例较高,占36.4%和27%,中期受到18℃和19℃等温线控制的区域比例最高,占23%和23%;中部区域,在渔汛前期、中期和末期中心渔场占所有A型渔场的比例分别为100%、81.3%和40.0%,其中,前期受到19℃和20℃等温线控制的区域比例较高,占37.5%和45.8%,中期受到18℃等温线控制的区域比例最高,占37.4%;南部区域,在渔汛前期、中期和末期中心渔场占所有A型渔场的比例分别为91.7%、84.4%和46.7%,其中前期受到19℃等温线控制的区域比例最高,占66.7%,中期受到18℃等温线控制的区域比例最高,占40.6%。
可见,在渔汛前期和中期阶段,A型渔场的出现有利于渔场的形成,其中渔汛前期沿岸19℃或20℃等温线的出现以及渔汛中期沿岸18℃或19℃等温线的出现可以作为中心渔场形成的指标。

图6 不同水温类型A型渔场区分方法举例Fig.6 Example of the classification to different temperature types in “A” type fishing ground

图7 不同等温线控制情况下的中心渔场(渔场指数大于0.6)占所有A型渔场比例Fig.7 Percentage of the productive fishing ground (fishing ground index is larger than 0.6) accounting for all “A” type fishing ground under different controlling conditions of temperature isotherms
4 讨论与分析
水温作为海洋遥感中最容易获得的环境因子,对侦查鱼群和确定渔场形成有着决定性的作用[15]。在以往的渔场和水温的关系分析中,通常将水温数据栅格化处理利用经验统计的方法确定渔场的适宜温度范围[21—23],而在水温结构与渔场的关系上,往往只是定性的描述[24—25],缺乏定量的分析。在本研究中,由于秘鲁鳀一般生活在近表层的区域(50 m以上)[8,26],因此研究假设表层水温的分布情况与秘鲁鳀渔场存在关系,尝试利用表层水温结构确定渔场类型,以定量地分析不同沿岸水温分布情况与渔场的关系。
研究发现,以表层水温大于20℃的海水是否入侵到近岸为标志,可以将秘鲁沿岸渔场分成两种类型(图4)。在渔汛前期(图5),除了北部区域沿岸出现过大于20℃的暖水(B型渔场)情况以外,中部和南部都是小于20℃的冷水,而方差分析多重比较表明(表1),在渔汛前期的渔场指数与中期后期相比是显著地要高的;中期开始,出现温度大于20℃的暖水情况增多,而t检验表明,在中期3个区域出现A型渔场时的渔场指数要显著地高于B型渔场。可见沿岸出现温度大面积小于20℃的暖水的情况(A型渔场)将有利于秘鲁鳀渔场的生成。研究中选用20℃作为冷暖水的标志与前人对秘鲁鳀适宜水温的调查是一致的:秘鲁鳀的栖息水温范围在13~23℃之间[27],但是适宜水温主要在15~20℃[28]。同时研究表明,秘鲁鳀栖息的东南太平洋沿岸海域内存在着强劲的秘鲁上升流[29],上升流区域使得海域有着冷水[1]、低氧[10]及存在丰富的饵料[30]等特征正好适合了的秘鲁鳀的生物学特性,有利其生存。而在秘鲁外海,通常存在由赤道逆流带来的温度较高的赤道表层水(surface equatorial water SEW),SEW向南部和近岸延生,形成亚热带表层水(subtropical surface water,SSW)[30],SSW到达近岸与沿岸上升流水(upwelled cold coastal water,CCW)形成沿岸和亚热带表层水的混合区域(mixed coastal-subtropical water,MCS)[31],Swartzman等[31]通过调查发现秘鲁鳀一般出现在CCW和MCS内,SEW和SSW内秘鲁鳀的资源丰度很低,这表明暖水入侵近岸会形成不利于秘鲁鳀的栖息环境,因此不利于秘鲁鳀渔场的生成。将这些研究结果对比本研究可以认为,20℃等温线的存在是海域中冷暖水的标志,即大于20℃的海域一般存在着SEW和SSW,而小于20℃的海域一般存在着有利于秘鲁鳀渔场生成的CCW和MCS。
国内外学者对秘鲁鳀资源变动与大尺度的气候变化(如厄尔尼诺南方涛动现象)的关系也从生物因素上解释了冷水的A型渔场导致秘鲁鳀渔场指数增加的原因。iquen和Bouchon[9]也发现,在暖水年份,秘鲁鳀整体的分布呈现向南部海域和更深的水层迁移的趋势。Alheit和Niguen[29]归纳了近年来对ENSO现象发生的暖水年份海域内生物调查研究结果,包括:ENSO发生时秘鲁鳀会往近岸迁移,导致鱼群生活空间狭窄,食物竞争加剧,长时间来看必然导致秘鲁鳀资源的减少;同时秘鲁鳀的天敌如竹筴鱼类(Trachurus)得益于暖水的扩张,能够与秘鲁鳀的栖息地存在重叠因此能够对其进行大量的捕食,当时Alheit等便指出表层SST的变化即能作为发生这种现象的指标;此外,暖水的扩张还会导致秘鲁鳀的饵料生物——桡足类(Copepods)动物资源量的减少。可见水温结构的变动能够导致秘鲁鳀的迁徙以及通过生物捕食被捕食的关系改变资源的状态,因此决定了海域秘鲁鳀渔场情况的好坏。
到了渔汛后期,通过t检验发现A型渔场的渔场指数与B型渔场不存在显著的关系,同时通过方差分析和多重比较可以发现(表1),渔汛末期的时候渔场指数要显著地低于前面两个渔汛阶段。这可以通过上升流的季节性变换来解释:秘鲁上升流主要是由海表面强劲的信风(trade wind)和地球自转共同导致的,其中风是主要的因素[1],渔汛末期(6月末至7月)渔场已从夏秋季转入为冬季,而冬季海域上方常常存在着风场转变的情况,即吹向赤道的信风可能会减弱甚至改风向吹向极地,这将对海域的上升流有着减弱的作用[32]。而上升流的减弱,造成海域虽然没有暖水入侵但是渔场也不是很好的情况,这必然对秘鲁鳀的栖息环境带来不利影响。
进一步分析表明(图7),沿岸19℃(或20℃)等温线和18℃(或19℃)等温线分别可以作为渔汛前期和中期中心渔场形成的指标。渔场判断过程中也出现过沿岸海域存在大面积低温水(小于17℃)的情况(图7),但是这种情况形成中心渔场的频率没有出现18~20℃等温线的情况高。研究发现[33],强的上升流同时也伴随着强的离岸输送,导致了海域营养盐的不易聚集,这也会影响到秘鲁鳀对饵料生物利用进而导致秘鲁鳀资源量的变化。因此,若将低温水的出现假设为是由上升流过强导致的,那么就可以解释为什么出现低温水的时候出现中心渔场的频率较低的情况。
研究探究了秘鲁鳀渔场与表层水温结构的关系,得到了中心渔场的表征指标。但是仅仅只用表层水温进行预报确实会造成误差,例如渔汛末期使用这些表征指标就难以说明中心渔场的存在。因此建议在今后的分析中,也可以将秘鲁沿岸的垂直水温资料(Argo数据)和风场情况考虑进去,利用海水中温跃层的数据或建立算法将实时的风场资料反演成海水中上升流的情况。此外也应该考虑到海域其他动力学因素(如涡旋的存在)和长时间的气候变化对渔场的影响,为更好地预报渔场服务。
[1] 陈芃, 汪金涛, 陈新军. 秘鲁鳀资源变动及与海洋环境要素的关系研究进展[J]. 海洋渔业, 2016, 38(2): 206-216.
Chen Peng, Wang Jintao, Chen Xinjun. Review on relationship between oceanic environment factors and population dynamics of anchoveta (Engraulisringens)[J]. Marine Fisheries, 2016, 38(2): 206-216.
[2] Fréon P, Bouchon M, Mullon C, et al. Interdecadal variability of anchoveta abundance and overcapacity of the fishery in Peru[J]. Progress in Oceanography, 2008, 79(2/4): 401-412.
[3] 联合国粮农组织. 联合国粮农组织渔业统计数据——1950-2013年全球捕捞产量[DB/OL]. http://www.fao.org/fishery/statistics/global-capture-production/query/zh.
Food and Agriculture Organization of the United Nations. Food and Agriculture Organization of the United Nations’ fisheries database-Global capture production from 1950 to 2013[DB/OL]. http://www.fao.org/fishery/statistics/global-capture-production/query/zh.
[4] Aranda M. Developments on fisheries management in Peru: the new individual vessel quota system for the anchoveta fishery[J]. Fisheries Research, 2009, 96(2/3): 308-312.
[5] 彭淇, 王斐, 吴彬, 等. 2种罗非鱼加工下脚料产物替代秘鲁鱼粉养殖奥尼罗非鱼(Oreochromisniloticus×O.aureus)稚鱼效果评价[J]. 海洋与湖沼, 2014, 45(3): 602-607.
Peng Qi, Wang Fei, Wu Bin, et al. Evaluation of two produces from byproduct of Tilapia as replacements of Peru fish meal in practical diets of Juvenile Tilapia (Oreochromisniloticus×O.aureus)[J]. Oceanologia et Limnologia Sinca, 2014, 45(3): 602-607.
[6] 李励年, 缪圣赐, 熊敏思. 国际渔业动态[J]. 渔业信息与战略, 2015(2): 156-160.
Li Linian, Miao Shengci, Xiong Minsi. International fishery information[J]. Fishery Information and Strategy, 2015(2): 156-160.
[7] 韦震. 鱼粉: 上半年秘鲁鱼粉供应不足, 饲料厂寻求鱼粉替代品[J]. 当代水产, 2015(2): 68-69.
Wei Zhen. Fish meat: during the first half of the year the Peru fish meal was in short supply, so feed factories search the replacements of it[J]. Current Fisheries, 2015(2): 68-69.
[10] Levin L A. Oxygen minimum zone benthos: adaptation and community response to hypoxia[J]. Oceanography and Marine Biology: An Annual Review, 2003, 41: 1-45.
[11] 官文江, 陈新军, 潘德炉. 遥感在海洋渔业中的应用与研究进展[J]. 大连水产学院学报, 2007, 22(1): 62-66.
Guan Wenjiang, Chen Xinjun, Pan Delu. A review: application and research of remote sensing in marine fisheries[J]. Journal of Dalian Ocean University, 2007, 22(1): 62-66.
[12] 余为, 陈新军. 西北太平洋柔鱼栖息地环境因子分析及其对资源丰度的影响[J]. 生态学报, 2015, 35(15): 5032-5039.
Yu Wei, Chen Xinjun. Analysis of environmental conditions and their influence on the abundance of neon flying squid in the Northwest Pacific Ocean[J]. Acta Ecologica Sinica, 2015, 35(15): 5032-5039.
[13] 陈芃, 陈新军. 基于最大熵模型分析西南大西洋阿根廷滑柔鱼栖息地分布[J]. 水产学报, 2016, 40(6): 893-902.
Chen Peng, Chen Xinjun. Analysis to the habitat distribution of Argentine shortfin squid (Illexargentinus) in the Southwest Atlantic Ocean using the maximum entropy model[J]. Journal of Fisheries of China, 2016, 40(6): 893-902.
[14] Arellano C E, Swartzman G. The Peruvian artisanal fishery: changes in patterns and distribution over time[J]. Fisheries Research, 2010, 101(3): 133-145.
[15] 陈新军. 渔业资源与渔场学[M]. 北京: 海洋出版社, 2004: 116-141.
Chen Xinjun. Fisheries Biology and Oceanography[M]. Beijing: China Ocean Press, 2004: 116-141.
[16] Chen Xinjun, Tian Siquan, Chen Yong, et al. A modeling approach to identify optimal habitat and suitable fishing grounds for neon flying squid (Ommastrephesbartramii) in the Northwest Pacific Ocean[J]. Fishery Bulletin, 2010, 108(1): 1-14.
[17] Tian Siquan, Chen Xinjun, Chen Yong, et al. Evaluating habitat suitability indices derived from CPUE and fishing effort data forOmmatrephesbratramiiin the northwestern Pacific Ocean[J]. Fisheries Research, 2009, 95(2/3): 181-188.
[18] 李春喜, 邵云, 姜丽娜. 生物统计学[M]. 4版. 北京: 科学出版社, 2008: 85-110.
Li Chunxi, Shao Yun, Jiang Lina. Biological Statistics[M]. 4th ed. Beijing: Science Press, 2008: 85-110.
[19] Brown M B, Forsythe A B. Robust tests for the equality of variances[J]. Journal of the American Statistical Association, 1974, 69(346): 364-367.
[20] 金岳, 陈新军. 利用栖息地指数模型预测秘鲁外海茎柔鱼热点区[J]. 渔业科学进展, 2014, 35(3): 19-26.
Jin Yue, Chen Xinjun. Forecasting hotspots ofDosidicusgigasin the offshore waters of Peru using habitat suitability model[J]. Progress in Fishery Sciences, 2014, 35(3): 19-26.
[21] 陈峰, 陈新军, 刘必林, 等. 西北太平洋柔鱼渔场与水温垂直结构关系[J]. 上海海洋大学学报, 2010, 19(4): 495-504.
Chen Feng, Chen Xinjun, Liu Bilin, et al. Relationship between fishing ground ofOmmastrephesbartramiiand vertical temperature structure in the northwestern Pacific Ocean[J]. Journal of Shanghai Ocean University, 2010, 19(4): 495-504.
[22] 陈新军, 许柳雄. 北太平洋150°E~165°E海域柔鱼渔场与表温及水温垂直结构的关系[J]. 海洋湖沼通报, 2004(2): 36-44.
Chen Xinjun, Xu Liuxiong. Analysis of relationship between fishing ground ofOmmastrephebartramiand surface water temperature and its vertical distribution from 150°E to 160°E in the Northwestern Pacific[J]. Transactions of Oceanology and Limnology, 2004(2): 36-44.
[23] 杨胜龙, 张忭忭, 靳少非, 等. 中西太平洋延绳钓黄鳍金枪鱼渔场时空分布与温跃层关系[J]. 海洋学报, 2015, 37(6): 78-87.
Yang Shenglong, Zhang Bianbian, Jin Shaofei, et al. Relationship between the temporal-spatial distribution of longline fishing grounds of yellowfin tuna (Thunnusalbacares) and the thermocline characteristics in the Western and Central Pacific Ocean[J]. Haiyang Xuebao, 2015, 37(6): 78-87.
[24] 邵全琴, 戎恺, 马巍巍, 等. 西北太平洋柔鱼中心渔场分布模式[J]. 地理研究, 2004, 23(1): 1-9.
Shao Quanqin, Rong Kai, Ma Weiwei, et al. Study on spatial patterns ofOmmastrephesbartramiifishing ground in the Northwest Pacific Ocean[J]. Geographical Research, 2004, 23(1): 1-9.
[25] 陈新军. 关于西北太平洋的柔鱼渔场形成的海洋环境因子的分析[J]. 上海水产大学学报, 1997, 6(4): 263-267.
Chen Xinjun. An analysis on Marine environment factors of fishing ground ofOmmastrephesbartramiiin Northwestern Pacipic[J]. Journal of Shanghai Fisheries University, 1997, 6(4): 263-267.
[27] Gutiérrez M, Swartzman G, Bertrand A, et al. Anchovy (Engraulisringens) and sardine (Sardinopssagax) spatial dynamics and aggregation patterns in the Humboldt Current ecosystem, Peru, from 1983-2003[J]. Fisheries Oceanography, 2007, 16(2): 155-168.
[28] Muck P, Sanchez G. The importance of mackerel and horse mackerel predation for the Peruvian anchoveta stock (a population and feeding model)[C]//Pauly D, Tsukayama I. The Peruvian Anchoveta and Its Upwelling Ecosystem: Three Decades of Change. Manila: ICLARM, 1987: 276-293.
[29] Alheit J, Niquen M. Regime shifts in the Humboldt Current ecosystem[J]. Progress in Oceanography, 2004, 60(2/4): 201-222.
[30] Montecino V, Lange C B. The Humboldt current system: ecosystem components and processes, fisheries, and sediment studies[J]. Progress in Oceanography, 2009, 83(1/4): 65-79.
[31] Swartzman G, Bertrand A, Gutiérrez M, et al. The relationship of anchovy and sardine to water masses in the Peruvian Humboldt Current System from 1983 to 2005[J]. Progress in Oceanography, 2008, 79(2/4): 228-237.
[32] Castro L R, Claramunt G, Krautz M C, et al. Egg trait variation in anchovetaEngraulisringens: a maternal response to changing environmental conditions in contrasting spawning habitats[J]. Marine Ecology Progress Series, 2009, 381: 237-248.
[33] Bakun A, Weeks S J. The marine ecosystem off Peru: what are the secrets of its fishery productivity and what might its future hold[J]. Progress in Oceanography, 2008, 79(2/4): 290-299.
Evaluating the effects of sea surface temperature variations on fishing ground of anchoveta (Engraulisringens) in the southeast Pacific Ocean
Chen Peng1,4,Chen Xinjun1,2,3,4
(1.CollegeofMarineSciences,ShanghaiOceanUniversity,Shanghai201306,China;2.TheKeyLaboratoryofSustainableExploitationofOceanicFisheriesResources,MinistryofEducation,ShanghaiOceanUniversity,Shanghai201306,China; 3.NationalEngineeringResearchCenterforOceanicFisheries,ShanghaiOceanUniversity,Shanghai201306,China; 4.CollaborativeInnovationCenterforDistant-waterFisheries,Shanghai201306,China)
Anchoveta (Engraulisringens) is one of the important small pelagic species living in the southeast Pacific Ocean. Exploring the process that fishing grounds vary with oceanic environmental factors could help us to understand the variations in fish stocks, which is beneficial for fishing industry. In this study, based on the Peruvian fishery logbooks of anchoveta from 2005 to 2014 as well as sea surface temperature (SST) data, we developed the fishing ground index (FGI) by using the catch per unit effort (CPUE) and fishing effort to understand the variation of fishing ground. The fishing ground were also divided into different types and correlated with the SST isotherms in order to evaluate their forming mechanism. The results indicated that significant differences (P<0.01) were found for FGI with the fishing periods (earlier stage, mid stage and later stage) and fishing areas (north area, middle area and south area) on the basis of analysis of variance. According to the distribution of SST, we defined the fishing ground of anchoveta as “A” type fishing ground and “B” type fishing ground. When the warm sea surface waters (>20℃) had not intruded into the near shore area, the “A” type fishing ground occurred. Otherwise, the “B” type fishing ground occurred. Our findings suggested that the surface temperature structure with “A” type is favorable for the formation of fishing ground in the earlier or mid stage of fishing periods. Moreover, the 19℃ or 20℃ SST isotherm in earlier stage and the 18℃ or 19℃ SST isotherm in mid stage could be used as an indicator to search for the productive fishing ground.
Anchoveta (Engraulisringens); southeast Pacific Ocean; surface temperature structure; fishing ground
10.3969/j.issn.0253-4193.2017.04.008
2016-07-28;
2016-12-18。
上海市科技创新行动计划(15DZ1202200);海洋局公益性行业专项(20155014)。
陈芃(1992—),男,福建省建瓯市人,研究方向为渔业资源与渔场学。E-mail:pengchen@yeah.com
*通信作者:陈新军(1967—),男,教授,博士生导师,主要研究领域为渔业资源学。E-mail:xjchen@shou.edu.cn
S931
A
0253-4193(2017)04-0079-10
陈芃,陈新军. 表层水温结构变化对东南太平洋秘鲁鳀渔场的影响[J].海洋学报,2017,39(4):79—88,
Chen Peng,Chen Xinjun. Evaluating the effects of sea surface temperature variations on fishing ground of anchoveta (Engraulisringens) in the southeast Pacific Ocean[J]. Haiyang Xuebao,2017,39(4):79—88, doi:10.3969/j.issn.0253-4193.2017.04.008
猜你喜欢
中国水产(2021年12期)2021-12-06 11:14:00
今日农业(2021年2期)2021-11-27 19:19:53
中国特种设备安全(2021年4期)2021-10-13 06:42:14
大自然探索(2021年12期)2021-02-07 01:37:04
当代水产(2019年4期)2019-05-16 03:04:20
华人时刊(2018年15期)2018-11-18 16:31:42
知识经济·中国直销(2017年8期)2017-09-05 09:52:46
作品(2016年10期)2016-12-06 10:41:54
渔业致富指南(2016年12期)2016-11-11 07:07:24
中学政史地·教学指导版(2014年10期)2015-02-02 08:59:38
