
美洲龟甲轮虫的分类特征及与我国热带地区常见龟甲轮虫的比较
2017-04-12 09:49:52喻婧怡温展明韩博平林秋奇
水生生物学报 2017年2期
喻婧怡温展明韩博平,林秋奇,
(1. 暨南大学水生生物研究中心, 广州 510632; 2. 广东省水库蓝藻水华防治中心, 广州 510632)
美洲龟甲轮虫的分类特征及与我国热带地区常见龟甲轮虫的比较
喻婧怡1温展明1韩博平1,2林秋奇1,2
(1. 暨南大学水生生物研究中心, 广州 510632; 2. 广东省水库蓝藻水华防治中心, 广州 510632)
美洲龟甲轮虫Keratella americana Carlin, 1943年被认为主要分布在美洲地区, 但陆续报告在其他地区有出现。2015年在对华南地区浮游动物调查时发现, 星云湖、抚仙湖、百色水库及飞来峡水库中均出现该种类。检测了广西百色水库中龟甲轮虫的COⅠ序列, 与已知美洲龟甲轮虫序列相似度高达98%。中国该龟甲轮虫种群与墨西哥美洲龟甲轮虫种群间差异度为2%, 进一步确定为美洲龟甲轮虫。文章描述了美洲龟甲轮虫形态及生境, 使用扫描电镜法比较了美洲龟甲轮虫、螺形龟甲轮虫Keratella cochlearis (Gosse, 1851)、热带龟甲轮虫Keratella tropica (Apstein, 1907)及无棘龟甲轮虫Keratella tecta (Gosse, 1851)的咀嚼器。美洲龟甲轮虫主要特点为: 背甲长度远大于宽度, 侧中棘刺短于侧棘刺, 背甲中央最后一个板块未封闭, 向后棘刺延伸。4种龟甲轮虫咀嚼器主要区别在于砧基的形状, 美洲龟甲轮虫砧基比其他3种轮虫发达, 螺形龟甲轮虫砧基似“T”形, 热带龟甲轮虫砧基为扇形, 无棘龟甲轮虫平直。种间的砧基存在明显差异, 可作为热带地区龟甲轮属的分类依据。
美洲龟甲轮虫; 形态描述; 咀嚼器; 热带地区
龟甲轮属Keratella Bory de St. Vincent隶属于单巢目臂尾轮科, 目前全世界记录龟甲轮虫53种,在淡水及咸水中均有出现, 我国目前报道的龟甲轮虫有17种。龟甲轮属中的螺形龟甲轮虫和无棘龟甲轮虫等为广布种, 缘锯龟甲轮虫等为暖水种[1]。龟甲轮虫的个体小, 背甲长大约为80—120 μm, 龟甲轮虫常作为富营养化水体的指示种, Gopko等[2]认为龟甲轮虫指数(KIN)可作为评判营养状况的指数, 指数越高营养水平越高, 但也有报道指出在寡营养型水库中螺形龟甲轮虫可以成为绝对优势种[3]。诸葛燕[4]、王全喜等[5]曾对龟甲轮虫新种进行了报道。美洲龟甲轮虫被认为是美洲地区特有种类, 但陆续报告在其他地区有出现[6], 认为该种类正通过人类的某种方式在全球扩散。美洲龟甲轮虫在形态上与螺形龟甲轮虫相似, 在常规生态调查中容易出错漏。目前龟甲轮属的定种主要依据背甲上板块的排列, 一些形态相似或接近的种类需要对咀嚼器作进一步的对比, 咀嚼器对分类及系统发育有指示作用[7], 我国对龟甲轮属的咀嚼器分析也比较少。2015年我们开展华南地区浮游动物多样性调查时, 在云南、广西及广东三省均发现美洲龟甲轮虫且有较高的丰度。本文利用分子标记, 对所获样品进行了确认, 并对该种形态及分布进行详细描述,与我国热带地区3种常见的龟甲轮虫进行了比较,为轮虫今后的定种提供参考。
1 材料与方法
1.1 样品采集
于2015年分别对广西百色水库(6月)、广东飞来峡水库(11月)及云南星云湖、抚仙湖(12月)进行调查。用YSI85型水质仪现场测定水体酸碱度(pH)、水温(T)等环境参数。定性样品用64 μm孔径的浮游动物网在水平垂直拖网2—3次, 加入96%乙醇, 于4℃保存。定量样品用采水器从表层0.5 m处到底层, 每隔1 m取水5 L, 用64 μm网过滤后, 加入4%甲醛现场固定(云南未采定量样品)。用于分子鉴定的样品用95%的酒精保存。轮虫定种依据参考Koste[8]、Ahlstrom[9], 定量样品在显微镜下观察计数。
1.2 轮虫COⅠ扩增及数据分析
将广西百色水库中所采集的轮虫从酒精样品中挑出后, 将单个龟甲轮虫材料放入离心管中, 使用Hotshot方法提取DNA[10], 用COⅠ引物[11](ZPF1: TGTAAAACGACGGCCAGTTCTASWAATCATA ARGATATTGG; ZPR1: CAGGAAACAGCTATGA CTTCAGGRTGRCCRAARAATCA)。
对标本龟甲轮虫rDNA进行PCR扩增。PCR反应程序为: 98℃预变性1min→98℃变性40s→55℃(54℃、53℃)退火40s→68℃延伸1min, 各一循环→98℃变性40s→62℃退火40s→68℃延伸1min, 循环32次→68℃延伸5min→4℃保存。将广西龟甲轮虫COⅠ序列与GenBank上已有序列进行Blast, 寻找与其亲缘关系最近的种。测序结果经Bioedit软件处理后, 使用MEGA6.0软件分析墨西哥美洲龟甲轮虫种群与采集的龟甲轮虫种群间差异度。以萼花臂尾轮虫作为龟甲轮属的外类群, 将百色水库龟甲轮虫的COⅠ序列与GenBank中已有的4种龟甲轮虫COⅠ序列构建邻接(NJ)分子系统树。
1.3 形态学描述
利用显微镜(OlumpusCX41)进行外部形态学观察, 与我国热带地区常见龟甲轮虫(螺形龟甲轮虫、热带龟甲轮虫、无棘龟甲轮虫)进行形态学比较, 其中螺形龟甲轮虫、无棘龟甲轮虫采集于广州流溪河水库、热带龟甲轮虫采集于珠海南屏水库。测量4种龟甲轮虫背甲长、背甲宽及美洲龟甲轮虫棘刺长度。制作轮虫咀嚼器方法参考De Smet[12]。在载玻片上滴上甘油, 将圆形盖玻片(直径1 cm)放上固定。将3—5只轮虫置于圆形盖玻片上, 加入3%的次氯酸钠溶液, 在解剖镜下观察其溶解过程。用毛细玻璃管吸取次氯酸钠溶液, 同时用胶头滴管加入纯净水。反复冲洗8—10次, 直至圆形盖玻片上无结晶。风干24h后, 喷金后在扫描电子显微镜(Hitachi TM-3000)下观察拍照。用绘图仪(Nikon H550L)对4种轮虫外部形态进行绘图。
2 结果
2.1 COⅠ序列分析
将百色水库龟甲轮虫的COⅠ序列与GenBank中已知序列进行比对, 发现该龟甲轮虫与美洲龟甲轮虫的COⅠ序列相似度最高, 有98%的相似度。该龟甲轮虫(N1-N3)与从GenBank中下载的美洲龟甲轮虫序列(AG0386-AG0388)以100%置信度聚为一枝(图 1)。通过K2P双参数模型计算, 该龟甲轮虫种群与墨西哥美洲龟甲轮虫种群间差异度为2%, 这低于轮虫的种间差异度3%[13], 可以确定该龟甲轮虫为Keratella americana Carlin, 1943。

图 1 基于龟甲轮虫COⅠ序列构建的邻接(NJ)系统树Fig. 1 The Neighbor-Joining tree based on COⅠ sequences of four Keratella speciesN1-N3为广西百色水库龟甲轮虫的COⅠ序列N1-N3 represent the COⅠsequences of Keratella species from Baise Reservoir
2.2 美洲龟甲轮虫雌性个体形态描述
背甲前端有三对棘刺, 前中棘刺最长, 侧棘刺长于亚中棘刺, 三对均向腹面弯曲, 背甲后端有一根较长的棘刺, 背甲长宽比为1.8—3.7, 比龟甲轮属中的其他种类长宽比都大。从侧面看, 外侧棘刺向腹面弯曲, 末端或多或少向内弯曲, 腹面前端呈U型凹陷, 通常背甲中部有一排板块, 最前端板块呈一个未闭合的六边形, 前中板块及中间板块为两个完整的六边形板块, 后中板块类似梯形, 并向末端延伸。在其两侧各有两个封闭的板块[9](图 2a、图 3ab)。咀嚼器: 左右几乎对称, 砧基呈火腿形, 基部细长, 逐步向末端扩大, 末端不具托; 砧枝侧缘1/3处各有一翼状突起, 砧枝有7对齿状突出, 砧枝背部中央各有一背基孔(Basifenestras); 槌钩具8对齿, 前端第一对齿最大, 向后逐渐变小, 第7、8对齿退化, 槌钩齿间的间距形状呈长条形。槌柄背面边缘及末端基部各具一小孔。测量: n=30, 背甲长=(104.6± 6.1) μm, 背甲宽=(48.1±5.6) μm, 前棘刺长度=(27.3± 7.7) μm, 后棘刺长度=(90.6±10.2) μm, 咀嚼器长= 18.5 μm。
2.3 中国热带地区常见龟甲轮属种类检索表
1. 没有后棘刺…………………………………2
有一根或两根后棘刺………………………3
2. 背甲中央有一条脊状凸起……………无棘龟甲轮虫K. tecta
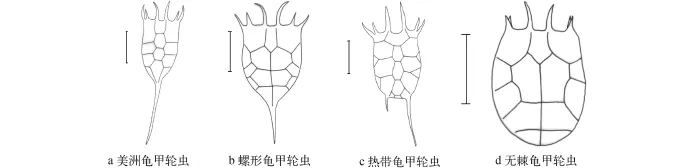
图 2 四种龟甲轮虫背面观Fig. 2 Dorsal view of four Keratella speciesa. 美洲龟甲轮虫; b. 螺形龟甲轮虫; c. 热带龟甲轮虫; d. 无棘龟甲轮虫. 比例尺为0.5 mma. Keratella americana; b. K. cochlearis; c. K. tropica; d. K. tecta. Scale bar=0.5 mm
背甲中央无脊状凸起…………缘锯龟甲轮虫K. lenzi
3. 有一根后棘刺………………………………4
有两根后棘刺………………………………5
4. 背甲中央有一条脊状凸起……………螺形龟甲轮虫K. cochlearis
背甲中央无脊状凸起……………美洲龟甲轮虫K. americana
5. 背甲末端有一长方形板块…………热带龟甲轮虫K. tropica
螺形龟甲轮虫Keratella cochlearis (Gosse, 1851)(图 2b、图 3c-d)背甲前端有三对棘刺,中间棘刺最长, 侧中棘刺比侧棘刺短。背甲后端有一根棘刺, 长度不均一。背甲宽度为长度的2/3。背甲中间有一条脊, 从前中区域延伸至后棘刺。中部有一对封闭的六边形板块, 两侧面各有一对闭合的多边形板块。咀嚼器: 砧基呈棒状, 似倒"T"形;砧枝呈三角形, 前段有齿状突出, 背面各有一个背基孔, 孔的面积为砧枝面积的1/4; 左右槌钩根部各有8个齿, 所有的齿由大逐渐变小, 第7, 8对齿退化,槌钩齿间的间距形状呈倒三角形; 槌钩与槌柄相连处膨大, 槌柄基部背面各有一个孔。测量: n=30, 背甲长=(85.5±8.3) μm, 背甲宽=(47.5±4.4) μm, 咀嚼器长=15 μm。分布: 广布种, 非洲区、南极区、澳新区、新北区、新热带区、东洋区、古北区。
热带龟甲轮虫Keratella tropica (Apstein, 1907) (图 2c、图 3e-f)背甲后端有两根棘刺, 左右两根棘刺不等长, 长度随环境变化。背甲无脊状突起,中间有一排板块, 后三个板块为闭合六边形, 背甲中央末端具一方形小板块, 是区分其与曲腿龟甲轮虫的主要依据。咀嚼器: 砧基细长, 末端具托, 呈扇形; 砧枝呈钝三角形, 有齿状突出, 正面左右砧枝基部边缘各有一个小孔, 背面各有一个孔, 孔的面积为砧枝面积的1/9; 左右槌钩根部各有八个齿, 所有的齿由大逐渐变小, 第7、8对齿退化, 槌钩齿间距两头较大, 中间间距小。测量: n=30, 背甲长=(96.4± 7.3) μm, 背甲宽=(56.8±7.8) μm, 咀嚼器长=23.7 μm。分布: 广布种, 为暖水种, 非洲区、澳新区、新北区、新热带区、东洋区、古北区。
无棘龟甲轮虫Keratella tecta (Gosse, 1851)(图2d、图 3g-h)背甲后端无棘刺, 个体较小, 背甲中间有一条脊状突起, 中间有两个不规则的六边形,该种经常被误认为螺形龟甲轮虫亚种的变异。咀嚼器: 砧基细长平直; 砧枝呈三角形, 侧缘1/3处各有一附属片状结构, 前端有齿状突出; 左右槌钩各有8个齿, 第7、8对齿退化, 所有的齿长度由长逐渐变短, 槌钩齿间距两头较大, 中间间距紧凑。测量: n=30, 背甲长=(86.7±1.9) μm, 背甲宽=(52.5±7.5) μm,咀嚼器长=14.2 μm。分布: 广布种, 非洲区、澳新区、新北区、新热带区、东洋区、古北区。
2.4 分布及生态
百色水库位于广西省百色市(23°57′30.1″N 106°22′57.1″E), pH为8.4, 夏季表层水温30.5℃。轮虫主要优势种为螺形龟甲轮虫、剪形臂尾轮虫、热带龟甲轮虫。飞来峡水库位于广东省清远市(23°42′49.2″N 113°11′0.7″E), pH为7.7, 冬季表层水温为20℃。螺形龟甲轮虫、无棘龟甲轮虫及热带龟甲轮虫为轮虫优势种。百色水库美洲龟甲轮虫所占龟甲轮属丰度比例为30.8%, 而飞来峡水库仅为0.4%(图 4)。星云湖位于云南昆明市(24°22′36″N 102°46′47″E), pH为7.7, 冬季表层水温15.9℃, 与其共存的轮虫种类为螺形龟甲轮虫、热带龟甲轮虫、方形臂尾轮虫。抚仙湖位于云南省昆明市(24°37′58″N 102°54′04″E), pH为8.1, 冬季表层水温为14℃, 无棘龟甲轮虫、螺形龟甲轮虫、广布多肢轮虫也出现在该湖泊中。

图 3 4种龟甲轮虫咀嚼器Fig. 3 Trophi of four Keratella speciesa-b. 美洲龟甲轮虫(a. 腹面观; b. 背面观); c-d. 螺形龟甲轮虫(c. 腹面观; d. 背面观); e-f. 热带龟甲轮虫(e. 腹面观; f. 背面观); g-h. 无棘龟甲轮虫(g. 腹面观; h. 背面观)a-b. Keratella americana (a. Ventral view; b. Dorsal view); c-d. K. cochlearis (c. Ventral view; b. Dorsal view); e-f. K. tropica (e. Ventral view; f. Dorsal view); g-h. K. tecta (g. Ventral view; h. Dorsal view)
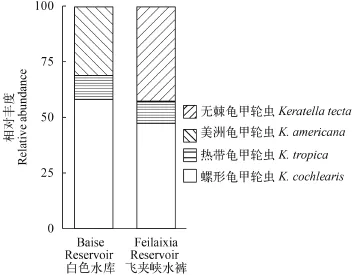
图 4 百色水库和飞来峡水库中美洲龟甲轮虫在龟甲轮属中相对丰度Fig. 4 The relative abundance of Keratella americana in Baise Reservoir and Feilaixia Reservoir
3 讨论
全世界龟甲轮属有53种, 其中8种为广布种[1]。美洲龟甲轮虫被认为主要分布在美洲地区, 但陆续报告在冰岛等其他地区出现[14]。我们将所采集美洲龟甲轮虫标本与我国热带地区常见龟甲轮虫进行了详细比较, 美洲龟甲轮虫与螺形龟甲轮虫形态较为相似。螺形龟甲轮虫与美洲龟甲轮虫均有一根后棘刺, 但美洲龟甲轮虫身体狭长, 后棘刺长度比身体长度略长, 且其长宽比例要比螺形龟甲轮虫大, 美洲龟甲轮虫背甲中央无脊状凸起, 为一排板块, 而螺形龟甲轮虫背甲中央具一脊状凸起将板块分为两列。广西水体中所测美洲龟甲轮虫背甲长为(108.5±6.9) μm, 后棘刺长为(89±9.7) μm, 广东美洲龟甲轮虫背甲长为(104.8±5.1) μm, 后棘刺长为(90±8.3) μm, 云南美洲龟甲轮虫背甲长为(101.1± 6) μm, 后棘刺长为(92.9±11.1) μm, 其中广西与云南美洲龟甲轮虫种群的背甲长有显著差异(P<0.05)。该测量结果与Ahlstrom[9]所测数据接近。美洲龟甲轮虫体长同环境因素有关, 食物的获得性可能是其形态变化的原因[15], 但水体中捕食者的数量对美洲龟甲轮虫后棘刺长度无显著影响[16]。
轮虫咀嚼器大小及形态在其生长过程中都不会发生改变, 对咀嚼器进行详细研究, 有助于区分相似种类和隐种[17]。我们所比较的四种轮虫在咀嚼器上的主要区别在于砧基的形状。螺形龟甲轮虫砧枝第一对齿最长, 热带龟甲轮虫砧枝第一对齿最短。在砧基形状上, 相比于其他3种龟甲轮虫, 美洲龟甲轮虫的砧基更加发达, 无棘龟甲轮虫砧基最窄, 螺形龟甲轮虫及热带龟甲轮虫末端均有托。砧基形状存在明显差异, 可作为热带地区龟甲轮属的分类依据。此外通过形态特征可将龟甲轮虫分为两种模式, 一类为矩形龟甲轮虫模式, 背甲中部有一列板块。第二类为螺形龟甲轮虫模式, 背甲有一个脊状凸起将其分为两侧板块。Ahlstrom[9]指出美洲龟甲轮虫属于矩形龟甲轮虫模式。将4种龟甲轮虫咀嚼器与已报道的龟甲轮虫咀嚼器进行对比后,发现槌钩齿的对数及砧基的形状是区分该属的主要特征。美洲龟甲轮虫咀嚼器与中华龟甲轮虫(Keratella sinensis Segers & Wang, 1997)[5]及诸氏龟甲轮虫(Keratella zhugeae Segers & Rong, 1998)[18]的咀嚼器相似, 中华龟甲轮虫与美洲龟甲轮虫砧基都很发达, 均似火腿形。诸氏龟甲轮虫及美洲龟甲轮虫的槌钩齿数均为8对, 而中华龟甲轮虫槌钩齿为6对。轮虫咀嚼器形状上的不同可能是对生境的适应, 咀嚼器形态与轮虫摄食习惯相关, 同时歧化选择对咀嚼器的形态变化有影响[19,20]。
美洲龟甲轮虫主要分布在南极区、新北区、新热带区, 目前已扩散至古北区及非洲区[1], 在河流、湖泊、酸性铀坑湖中均有发现[21,22], 其常与螺形龟甲轮虫共存[23,24]。近十几年在越南、中国、阿尔及利亚[25—27]相继报道了该种的出现。飞来峡水库自建库(1997)以来, 轮虫主要优势种为螺形龟甲轮虫, 未报道过美洲龟甲轮虫[28], 同样, 在百色水库及星云湖早期有调查中也未报道过该种的存在。我们在我国华南地区3个省的湖泊或水库中,均检出了美洲龟甲轮虫, 由于对这类小型生物通常缺少历史记录, 对扩散的时间和来源有待通过进一步遗传数据进行分析。
[1]Segers H, De Smet W H. Diversity and endemism in Rotifera: a review, and Keratella Bory de St Vincent [J]. Biodiversity and Conservation, 2008, 17(2): 303—316
[2]Gopko M, Telesh I V. Estuarine trophic state assessment: New plankton index based on morphology of Keratella rotifers [J]. Estuarine, Coastal and Shelf Science, 2013, 130(3): 222—230
[3]Lin Q Q, Zhao S Y, Han B P. Rotifer distribution in tropical reservoirs, Guangdong Province, China [J]. Acta Ecologica Sinica, 2005, 25(5): 1123—1131 [林秋奇, 赵帅营, 韩博平. 广东省水库轮虫分布特征. 生态学报, 2005, 25(5): 1123—1131]
[4]Yan Z, Huang X. On a new species of Keratella (Rotifera: Monogononta: Brachionidae) [J]. Hydrobiologia, 1997, 387—388(1—3): 35—37
[5]Segers H, Wang Q. On a new species of Keratella (Rotifera: Monogononta: Brachionidae) [J]. Hydrobiologia, 1997, 344(1—3): 163—167
[6]Hansson L A, Hylander S, Dartnall H J G, et al. High zooplankton diversity in the extreme environments of the McMurdo Dry Valley lakes, Antarctica [J]. Antarctic Science, 2012, 24(2): 131—138
[7]Guerrero-Jiménez G, Silva-Briano M, Adabache-Ortiz A, et al. Ultra Structural Comparison of Trophi in Species of the Genus Brachionus Pallas, 1766 in Aguascalientes State, Mexico [J]. Natural Science, 2015, 7(9): 425—437.
[8]Koste, W. Rotatoria. Die Rädertiere Mitteleuropas [M]. GebrÜder Borntraeger. Berlin. Stuttgart. 1978, 22
[9]Ahlstrom E H. A Revision of the Rotatorian Genus Keratella with Descriptions of Three New Species and Five New Varieties [M]. American Museum of Natural History. 1943, 435
[10]Montero-Pau J, Gómez A, Muñoz J. Application of an inexpensive and high-throughput genomic DNA extraction method for the molecular ecology of zooplanktonic diapausing eggs [J]. Limnology and Oceanography: Methods, 2008, 6(6): 218—222
[11]Prosser S, Martínez-Arce A, Elías-Gutiérrez M. A new set of primers for COI amplification from freshwater microcrustaceans [J]. Molecular Ecology Resources, 2013, 13(6): 1151—1155
[12]De Smet W H. Preparation of rotifer trophi for light and scanning electron microscopy [J]. Hydrobiologia, 1998, 387/388: 117—121
[13]García-Morales A E, Elías-Gutiérrez M. DNA barcoding of freshwater Rotifera in Mexico: Evidence of cryptic speciation in common rotifers [J]. Molecular Ecology Resources, 2013, 13(6): 1097—1107
[14]Evtimova V, Pandourski I. Rotifers and lower crustaceans from South-western Iceland [J]. Biodiversity Data Journal, 2016, 4(4): e7522
[15]Vásquez E, Koste W, Medina M L. Variaciones morfologicas del rotifero Keratella americana (Carlin, 1943) de una laguna de inundacion del Rio Orinoco, Venezuela [J]. Revue d'hydrobiologie tropicale, 1991, 24(2): 83—90
[16]Garza-Mouriño G, Silva-Briano M, Nandini S, et al. Morphological and morphometrical variations of selected rotifer species in response to predation: a seasonal study of selected brachionid species from Lake Xochimilco (Mexico) [J]. Hydrobiologia, 2005, 181(546): 169—179
[17]Fontaneto D, Giordani I, Melone G, et al. Disentangling the morphological stasis in two rotifer species of the Brachionus plicatilis species complex [J]. Hydrobiologia, 2007, 583(1): 297—307
[18]Segers H, Rong S. Two new species of Keratella (Rotifera: Monogononta: Brachionidae) from Inner Mongolia, China [J]. Hydrobiologia, 1998, 382(1—3): 175—181
[19]Salt G W, Sabbadini G F, Commins M L. Trophi morphology relative to food habits in six species of rotifers (Asplanchnidae) [J]. Transactions of the American Microscopical Society, 1978, 97(4): 469—485
[20]Hochberg R, Wallace R L, Walsh E J. Soft bodies, hard jaws: an introduction to the symposium, with rotifers as models of jaw diversity [J]. Integrative and Comparative Biology, 2015, 37(1—2): 180—188
[21]De Paggi S B J, Devercelli M. Land use and basin characteristics determine the composition and abundance of themicrozooplankton [J]. Water, Air, & Soil Pollution, 2011, 218(1—4): 93—108
[22]Ferrari C R, Azevedo H D, Wisniewski M J S, et al. An overview of an acidic uranium mine pit lake (Caldas, Brazil): composition of the zooplankton community and limnochemical aspects [J]. Mine Water & the Environment, 2015, 34(3): 343—351
[23]Nova C C, Lopes V G, Souza L C, et al. The effect of seasonality on the structure of rotifers in a black-water shallow lake in Central Amazonia. [J]. Anais Da Academia Brasileira De Ciências, 2014, 86(3):1359—72
[24]Brito S L, Maia-Barbosa P M, Pinto-Coelho R M. Zooplankton as an indicator of trophic conditions in two large reservoirs in Brazil [J]. Lakes & Reservoirs Research & Management, 2011, 16(4): 253—264
[25]Segers H. Zoogeography of the Southeast Asian Rotifera [J]. Hydrobiologia, 2001, 446—447(1): 233—246
[26]Yan Q Y, Yu Y H, Feng W S, et al. Genetic diversity of plankton community as depicted by PCR-DGGE fingerprinting and its relation to morphological composition and environmental factors in Lake Donghu [J]. Microbial Ecology, 2007, 54(2): 290—297
[27]Hamaidi-Chergui F, Hamaidi M S, Errahmani M B, et al. Studies on biodiversity of rotifera in five artificial lakes in Algeria: Systematical and zoogeographical remarks [J]. Kragujevac Journal of Science, 2013, 35: 115—138
[28]Lin Q Q, Zhao S Y, Han B P. Dynamics of rotifer and crustacean zooplankton in a newly constructed reservoir [J]. Chinese Journal of Ecology, 2006, 25(3): 270—276 [林秋奇, 赵帅营, 韩博平. 新建水库轮虫和甲壳类浮游动物动态特征. 生态学杂志, 2006, 25(3): 270—276]
MORPHOLOGICAL COMPARISON OF KERATELLA AMERICANA CARLIN, 1943 AND KERATELLA COMMON SPECIES IN TROPICAL CHINA
YU Jing-Yi1, WEN Zhan-Ming1, HAN Bo-Ping1,2and LIN Qiu-Qi1,2
(1. Institute of Hydrobiology, Jinan University, Guangzhou 510632, China; 2. Guangdong Center for Control and Prevention of Reservoir Cyanobacterial Blooms, Guangzhou 510632, China)
Keratella americana Carlin, 1943 mainly distributes in the America, and it had also been found in other continents. When we investigated zooplankton in South China in 2015, the species was found in Xingyun Lake, Fuxian Lake, Baise Reservoir and Feilaixia Reservoir. Its mitochondrial cytochrome oxidase subunitⅠ(COⅠ) gene was sequenced and showed a similarity of 98% with the sequences from GenBank, and this confirmed the species as K. americana. In the present study, a description was given of K. americana. Its trophi were compared with these of three common species: Keratella cochlearis (Gosse, 1851), Keratella tropica (Apstein, 1907) and Keratella tecta (Gosse, 1851), using a scanning electron microscope. K. americana is characterized by the following features: width of lorica narrower than its length; intermediate spines shorter than lateral spines; last facet in dorsal plate polygonal, open. The trophi of the four species differ in the shape of the fulcrum: The fulcrum of K. americana is stronger than the other three species. K. cochlearis has a T-shaped fulcrum and that of K. tropica is fan-shaped. The fulcrum of K. tecta is straight. The fulcrum therefore appears to significantly differ in shape and can be used for taxonomy and identification of different Keratella.
Keratella americana; Morphological description; Trophi; Tropics
Q959.181
A
1000-3207(2017)02-0448-06
10.7541/2017.56
2016-04-18;
2016-07-20
广东省科技计划项目(2013B080500022); 广东省水资源节保专项资助 [Supported by Science and Technology Planning Project of Guangdong Province (2013B080500022); Water Resource Conservation and Protection Project of Guangdong Province]
喻婧怡(1992—), 女, 汉族, 湖南长沙人; 硕士研究生; 主要从事水生生物学研究。E-mail: 270116321@qq.com
林秋奇, E-mail: tlinqq@jnu.edu.cn
猜你喜欢
科学养鱼(2020年10期)2020-11-23 09:04:10
家教世界·创新阅读(2020年6期)2020-07-09 03:14:57
———在龟甲、兽骨上刻字的朝代
家教世界(2020年16期)2020-07-08 09:08:56
课外语文·中(2020年2期)2020-03-24 22:18:09
课外语文(2020年5期)2020-03-14 08:16:40
奇闻怪事(2019年8期)2019-08-30 04:57:39
知识窗(2019年4期)2019-04-26 03:16:02
科学之友(2019年1期)2019-02-24 08:22:24
中成药(2018年10期)2018-10-26 03:41:34
作文评点报·中考版(2017年1期)2017-03-06 21:37:28
