
长江中下游湖泊群落水平下沉水植物碳、氮、磷化学计量特征及其影响因素*
2017-04-12 01:18苏豪杰夏午来
湖泊科学 2017年2期
苏豪杰,吴 耀,夏午来,谢 平,曹 特
(1:中国科学院水生生物研究所东湖湖泊生态系统试验站,武汉 430072)(2:中国科学院大学,北京 100049)
长江中下游湖泊群落水平下沉水植物碳、氮、磷化学计量特征及其影响因素*
苏豪杰1,2,吴 耀1,2,夏午来1,2,谢 平1**,曹 特1
(1:中国科学院水生生物研究所东湖湖泊生态系统试验站,武汉 430072)(2:中国科学院大学,北京 100049)
以长江中下游14个湖泊沉水植物为研究对象,分析群落水平下沉水植物碳(C)、氮(N)、磷(P)的化学计量特征及其影响因素. 结果显示,沉水植物群落C含量平均为386.93±25.80 mg/g,范围为315.98~441.97 mg/g,N和P含量的平均值分别为26.10±4.84和2.64±0.99 mg/g,范围分别为13.75~40.89和1.01~5.92 mg/g. 与物种水平研究相比,群落水平下沉水植物C、N、P化学计量特征变异系数较小,内稳定性更强. 群落C含量与P含量具有显著相关性,而与N含量不相关,这说明C元素与N和P之间的耦合关系并不一致;N∶P比与P含量之间具有极显著的负相关关系,但与N含量不相关,说明沉水植物群落P含量的变异对N∶P比起主导作用. 冗余分析表明,环境因子解释了沉水植物群落化学计量特征总变异的30.2%,在P<0.01水平下对沉水植物群落化学计量特征具有显著影响的环境因子为底泥总磷、底泥N∶P比、水深和消光系数. 变差分解结果表明,底泥和上覆水分别独自解释了化学计量特征总变异的5.21%和10.19%,其交互作用解释了总变异的5.25%. 通过该研究结果推测,在恢复湖泊水生植被的实践过程中,相对于底泥,对上覆水光照条件进行控制可能更加迫切.
沉水植物群落;生态化学计量学;长江中下游湖泊;底泥;上覆水
生态化学计量学综合了生物学、化学和物理学的基本原理,利用生态过程中多重化学元素的平衡关系,为研究C、N、P等元素在生态系统过程中的耦合关系提供了一种综合方法[1-2]. 目前,化学计量学作为一种新的生态学研究工具已经被应用于分子、细胞、个体、群落以及生态系统等各个层次[3-4].N和P是生物体的重要组成成分,是生命活动过程所必需的大量营养元素,也是生态系统中限制生物生长的重要限制因子[5]. 氮磷比(N∶P比)不仅是决定群落结构和功能的关键指标,还可以作为判断环境因子,特别是土壤对植物生长的养分供应状况的指标[5-6];碳氮比(C∶N)和碳磷比(C∶P)则表述了生物量与养分之间的比值关系,即养分利用效率[7]. 因此,生物体内的任一元素的稀缺或过量都会导致内部元素比例失衡,从而影响到整个生态学过程,如生态系统的初级生产力以及种群动态等[8-10].
沉水植物是湖泊中重要的初级生产者,在维持清水状态方面发挥着独特的作用[11]. 与陆生植物不同,沉水植物的生长不仅受到养分条件的限制,还会受到低光照条件的胁迫[12-13]. 而且,沉水植物吸收N、P等营养元素既可以通过根部从沉积物中获取,也可以通过枝叶从水中获取[14-15]. 因此,沉水植物C、N、P的化学计量特征会受到多方面因素的影响. 虽然大型沉水植物的生长不易受沉积物营养条件的制约,但是研究表明沉积物条件却可以决定沉水植物的生活型[16]. 如,在沉积物贫瘠时,生物量较低的底栖型植被占据优势;而在沉积物肥沃时,生物量较高的冠层型植被占据优势地位. 此外,沉积物类型(沙质或黏质)会影响沉水植物生物量的分配关系[17],从而导致沉水植被化学计量特征的变化. 另一方面,虽然根生大型沉水植物主要是从沉积物中而不是上覆水中吸收营养,但上覆水中的光照条件会影响植物光合作用的强弱从而改变化学计量特征[18-19]. 有研究表明,低光照条件或者水位的增加会迫使沉水植物分配更多的生物量到茎,使其可以伸展到水面以减轻低光照条件的限制[20],导致沉水植物茎中C∶N、C∶P和N∶P比的升高[21]. 因此,底泥与上覆水的物理化学特性可能是影响沉水植物化学计量特征的两个重要变异来源.
目前国内对水生态系统的生态计量学研究主要是针对植物物种水平的调查以及个体水平的研究[22-26],基于群落水平的研究较少. 那么群落水平下沉水植物的C、N、P的化学计量特征是怎样的?底泥与上覆水是如何对沉水植物群落化学计量特征产生影响的?其相对重要性如何?基于以上问题,本文通过对长江中下游14个湖泊沉水植被进行调查,分析群落水平沉水植物C、N、P的化学计量特征,并研究了其与环境因子的关系,以期为我国区域群落沉水植物元素计量学研究及沉水植被的恢复提供数据支持.
1 材料与方法
1.1 样品采集与处理
于2014年6-8月对长江中下游的太湖、高邮湖、骆马湖、龙感湖、赤湖、花园湖、武昌湖、野猪湖、武湖、斧头湖、梁子湖、洪湖、崇湖、西凉湖14个湖泊进行沉水植物的调查与采集(图1). 采集植物之前,对植物所在区域的水深、透明度以及水下的光合有效辐射进行测定,并采集底泥与水体样品. 透明度和水深分别用直径为30 cm的透明度盘和测深锤测定. 水下光合有效辐射用照度计(Li-COR UWQ7839, Li-Cor, Lincoln,NE)测定. 底泥用柱状采泥器采集表层0~10 cm获得,水样用采水器在水下0.5 m处获取. 水生植物使用水下镰刀(0.2 m2)进行采集,每个采样点随机采集至少3个样方,每个样方采集到的沉水植物洗去泥沙和杂物,沥干浮水,分拣出种类并分别称取鲜重. 每个物种取2~3株地上部分样品(包括茎和叶)装进封口袋并贴上防水标签. 水样、底泥以及植物样品一并放入便携式冰箱带回实验室进一步处理.

图1 长江中下游采样湖泊分布
水生植物样品与底泥在80℃烘箱里烘干至恒重,用研钵磨碎后过100目筛,用元素分析仪(Flash EA 1112 series, CE Instruments, Italy)测定C、N含量,用硫酸-过氧化氢消解植物样品,硫酸-高氯酸消解消解底泥样品,钼锑抗分光光度法分别测定P含量[27]. 底泥含水率用质量法测定,即先将底泥于105±1℃条件下烘干至恒重,以前、后质量的差值计算含水率.参照文献[28]测定水样中的总氮(TN)、总磷(TP)和叶绿素a(Chl.a)浓度.
1.2 数据分析
消光系数(light attenuation coefficient,K)是由纯水、腐殖质浓度、颗粒物浓度和植物的光合作用产生的消光率组成,其计算方法为[29]:
K=(1/Z)ln(I0/IZ)
(1)
式中,I0为表层光合有效辐射,IZ为水深Z处的光合有效辐射.
沉水植物群落的C、N、P含量是群落内物种C、N、P含量的加权平均数(以相对生物量为依据). 因此,群落水平下沉水植物的C、N、P含量为:
群落C/N/P=(相对生物量1×C/N/P1)+(相对生物量2×C/N/P2)+… +(相对生物量n×C/N/Pn)
(2)
当群落内物种的C/N/P数据缺失时,则用该物种C/N/P含量的平均值代替.
采用PASWStatistics18(SPSSInc.,Chicago,USA)对沉水植物群落C、N、P、C∶N比、C∶P比和N∶P比之间进行Pearson相关性分析;采用CANOCO(canocoforwindows5.0)中的冗余分析(redundancyanalysis,RDA)揭示底泥与水体中环境因子对群落化学计量特征的影响,使用MonteCarlo置换检验(置换次数=999)来筛选出显著性的环境变量(P<0.01)[30];采用R语言[31]环境中的vegan数据包中的偏冗余分析(partialredundancyanalysis,pRDA)进行方差分解,以阐释底泥与上覆水对沉水植物群落化学计量特征的单独影响与交互作用.
2 结果与分析
2.1 环境因子的统计特征
对所调查的14个湖泊共计126个水生植物群落所在区域的环境进行分析,结果表明,底泥C含量平均为20.26±14.78mg/g,底泥N含量平均为2.03±1.06mg/g,底泥P含量平均为0.56±0.16mg/g,含水率为49%±13%. 底泥N含量均与底泥C含量(r=0.882,P<0.001)和含水率(r=0.793, P<0.001)之间具有较高的相关关系,而与底泥P(r=0.279,P=0.002)含量之间的相关关系较弱(图2). 14个湖泊水体TN浓度平均值为0.71±0.19mg/L,TP浓度平均值为0.04±0.03mg/L,消光系数为2.47±1.09.

图2 底泥N含量分别与C含量、含水率和P含量之间的相关关系
2.2 沉水植物群落C、N、P生态化学计量学特征及其与环境因子之间的关系
沉水植物群落C含量平均为386.93±25.80 mg/g,范围为315.98~441.97 mg/g,N和P含量的平均值分别为26.10±4.84和2.64±0.99 mg/g,范围分别为13.75~40.89和1.01~5.92 mg/g. 3种元素含量的变异系数由大到小表现为P>N>C. 由于P含量变化较大,导致C∶P比和N∶P比变幅较大(变异系数分别为44.30%和34.10%)(图3).

图3 沉水植物群落C、N、P含量和C∶N, C∶P, N∶P比频率直方图(n=126)
Pearson相关分析结果表明(表1),沉水植物群落C含量与N含量没有相关性(r=0.066,P=0.462),而与P含量之间具有显著的负相关关系(r=-0.254,P=0.004). 这说明C作为植物体内干物质的最主要元素,与N和P元素之间的耦合关系并不一致. N和P含量之间具有极显著的相关关系(r=0.578,P<0.001),说明N和P作为蛋白质和RNA的重要元素,在共同执行生命活动中具有较强的耦合关系. 但同时发现,N∶P 比与P含量之间具有极显著的相关关系(r=-0.799,P<0.001)而与N无显著相关关系(r=-0.139,P=0.121),由此可见沉水植物群落P含量的变异对N∶P比的动态变化起主导作用.

表1 沉水植物群落C、N、P化学计量特征之间的相关关系(n=126)
*代表P<0.05,显著相关;**代表P<0.01,极显著相关.
冗余分析(RDA)解释了总变异的30.2%(df=15,F=3.17,P=0.001)(图4). 在P<0.01水平下对沉水植物群落化学计量特征具有显著影响的环境因子为底泥P(r2=0.1367,P=0.002)、底泥N∶P比(r2=0.0989,P=0.002)、水深(r2=0.1504,P=0.002)和消光系数(r2=0.1301,P=0.001)(表2). 偏冗余分析结果表明,底泥和水体分别独自解释了总变异的5.21%和10.19%,其交互作用解释了化学计量特征总变异的5.25%.

图4 环境因子与沉水植物群落化学计量特征之间的RDA分析
表2 底泥、上覆水与沉水植物群落化学计量特征之间的RDA分析结果*
Tab.2 Results of RDA analysis for sediment,water column and stoichiometric signatures of submerged macrophyte community

指标底泥r2P指标上覆水r2P底泥N0.02040.295水深0.15040.002底泥C0.02640.191透明度0.01160.479含水率0.01200.492TN0.06740.012底泥P0.13670.002TP0.07290.012底泥C∶N比0.01830.324Chl.a0.04620.035底泥C∶P比0.05470.039消光系数0.13010.001底泥N∶P比0.09890.002透明度水深比0.05630.022水体N∶P比0.05850.030
*字体加粗表示P<0.01.
3 讨论
作为群落水平上大尺度的C、N、P化学计量研究,通过对比发现,本研究中C含量高于Xia等[22]在中国东部区域的研究结果,N含量与中国东部区域水平基本一致,而P含量低于中国东部区域,这也导致了本研究中C∶P比较高;与长江中下游区域物种水平的研究相比[25],本研究中C、N含量较高,而P含量较低(表3). 研究还发现,在群落水平下化学计量特征的变异系数,无论是C、N、P含量还是C∶N、C∶P、N∶P比,都低于中国东部区域[22]和长江中下游区域[25]物种水平的研究结果,这说明群落水平下沉水植物C、N、P化学计量特征具有更高的内稳定性. 本研究作为大尺度的空间调查,很难做到取样时间一致. 此外,研究表明沉水植物物种间的化学计量特征的变异大于物种内部的变异[26]. 因此,群落水平下沉水植物化学计量特征主要取决于群落结构,而采样时间对其影响相对较小.
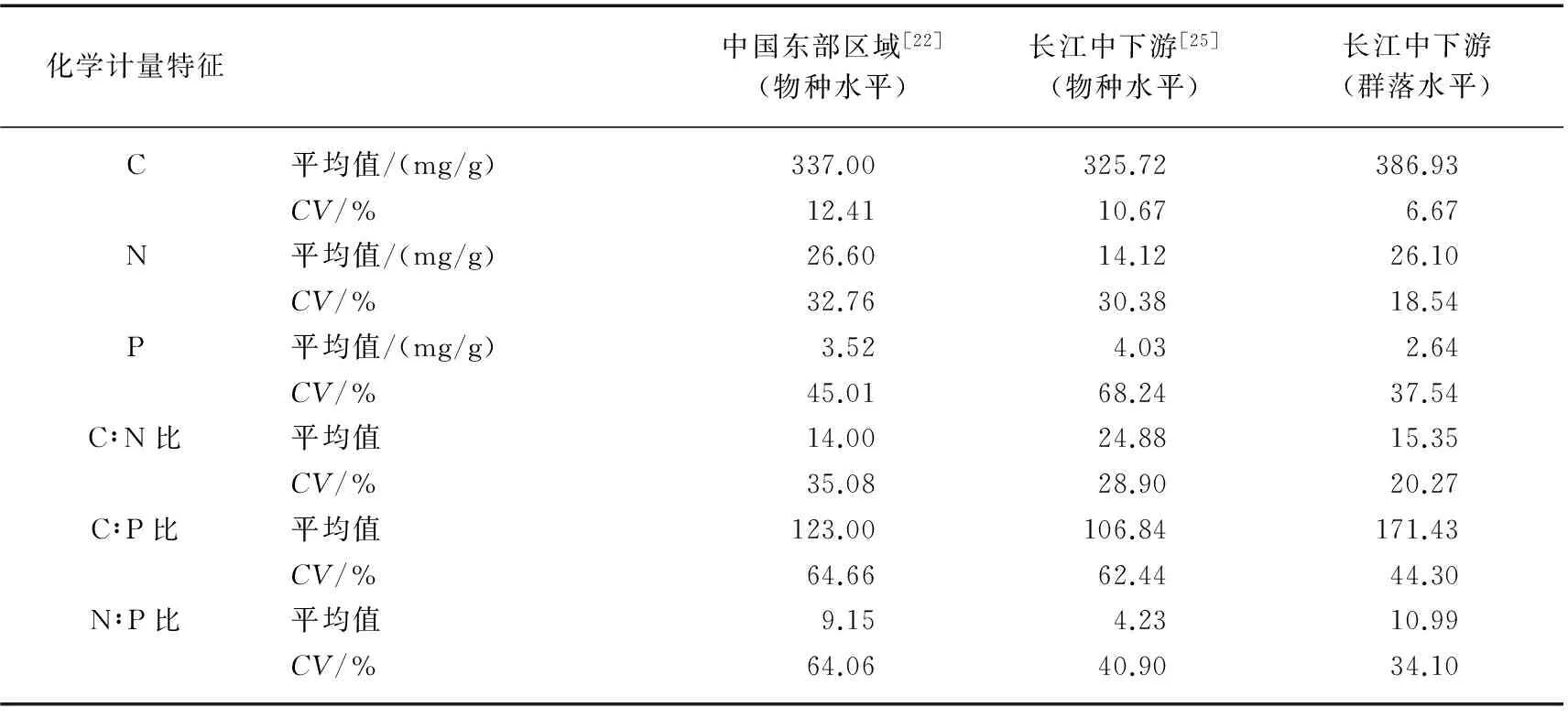
表3 长江中下游及中国东部区域沉水植物在物种和群落水平下的化学计量特征
通常情况下,我们认为大尺度上的化学计量特征是对环境条件长期适应的结果. 然而,这并不意味着外界环境条件不会对C、N、P化学计量特征产生影响. 其中,外界营养条件是影响沉生植物C、N、P化学计量特征的重要影响因素,而底泥是根生沉水植物生长所需营养物质的重要来源. 本研究中,底泥TP和N∶P比均会对沉水植物群落化学计量特征产生显著影响. 本次调查底泥TN、TP含量平均值分别为2.03和0.56 mg/g,与云南高原湖泊洱海相比(TN=4.2 mg/g,TP=0.90 mg/g)较低[32-33]. 这可能是长江中下游湖泊大多是浅水湖泊,底泥受风浪的影响较大,营养物质频繁的再悬浮所致. 此外,本次调查无论是底泥TN、TP还是含水率都显著低于无沉水植物分布的武汉东湖(TN=4.55 mg/g,TP=0.73 mg/g;含水率76%,苏豪杰,未发表数据). 这说明,长江中下游湖泊底质的营养条件并不会限制沉水植物的发展,相反,调查中发现,沉水植物更喜于分布在营养含量较低、底质较硬的区域中,使得根生沉水植物可以牢固地定植于水体中,抗风浪等外界干扰能力更强. 本研究还发现,底泥N与C含量之间有较强的相关性,但与P之间的耦合关系较弱(图2). 底泥N、P的这种非同步关系可能会导致沉水植物“供应-需求”上的错配矛盾,从而导致水生植物化学计量特征的变化.通过Monte Carlo置换检验筛选显著的环境因子发现,底泥的N∶P比会对沉水植物群落化学计量特征产生显著的影响(P<0.01),恰好验证了这一观点. 此外,还可以根据植物组织中N∶P比来判断环境中的养分状况,当植物N∶P比<14时,表现为受N的限制;当N∶P比>16时,表现为受P的限制[3]. 本研究中沉水植物群落N∶P比平均值为10.99±3.75,表明长江中下游湖泊沉水植物群落P过量而N相对不足.
光照是影响沉生植物C、N、P化学计量特征的另一个重要影响因素. 李威等[24]在洱海中研究发现,苦草叶片C含量、C∶N比和C∶P比均随水深增加而降低,N和P含量则随水深增加而升高. 然而在本研究中却有着截然不同的结果,即C∶N、C∶P和N∶P比随着水深增加而增加,而N和P含量则随水深的增加而减小, 这说明沉水植物在群落和物种水平下对水深的响应是不同的. 这可能是由于不同的沉水植物对水深的响应具有不同的生长和代谢策略[34]所导致的. 随水深的增加,沉水植物群落C∶N比、C∶P比的增加可能有以下两方面的原因. 一是,马来眼子菜(Potamogetoncrispus)和狐尾藻(Myriophyllumverticillatum)等冠层型植物为了减弱低光照的胁迫,伸长茎以获得光照,从而导致C∶N比和C∶P比升高;二是,随着水深的增加,C∶N比和C∶P 比较高的沉水植物(如马来眼子菜、狐尾藻等)较其他沉水植物更具有竞争优势,导致这些植物在群落结构中的比例升高,从而导致C∶N比和C∶P比升高. 根据调查,在浊度较高的浅水湖泊中,狐尾藻和马来眼子菜能够通过伸长茎将叶“漂浮”于水面. 在水深小于株高的情况下,水深和透明度都已不再是其生长的限制因素,因而在竞争中能够占据优势. 因此可以推测,狐尾藻和马来眼子菜等冠层型植物更能够适应浊度较高的浅水湖泊环境.
变差分解结果表明,底泥和水体均会对沉水植物群落化学计量特征产生显著影响,说明沉水植物N、P组织营养浓度与湖泊的富营养化状态有关[35]. 水体比底泥的影响作用更为明显,可能是因为长江中下游湖泊多数为富营养化湖泊,营养条件已不再是限制水生植物生长的制约因素,而由富营养化所引起的低透明度、高消光系数等水体环境条件则严重影响了沉水植物的光合作用,进而导致内部化学元素失衡及沉水植物的消亡. 这与Xing等[25]在长江中下游湖泊研究所得出的结论是一致的. 因此,根据结果可以推测,在恢复湖泊水生植被的实践过程中,对于上覆水光照条件(如水位、透明度、消光系数等)的控制相对于底泥可能更加迫切.
致谢:感谢中国科学院水生生物研究所陈隽研究员和过龙根副研究员的指导,感谢杨磊和卢巍在野外采样中的帮助.
[1] Elser JJ, Sterner RW, Gorokhova Eetal. Biological stoichiometry from genes to ecosystems.EcologyLetters, 2000, 3: 540-550.
[2] He Jinsheng, Han Xingguo. Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems.ChineseJournalofPlantEcology, 2010, 34: 2-6. [贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论. 植物生态学报, 2010, 34: 2-6.]
[3] Elser JJ, Dobberfuhl DR, MacKay NAetal. Organism size, life history, and N: P stoichiometry toward a unified view of cellular and ecosystem processes.BioScience, 1996, 46: 674-684.
[4] Cheng Bin, Zhao Yongjun, Zhang Wenguangetal. The research advances and prospect of ecological stoichiometry.ActaEcologicaSinica, 2010, 30(6): 1628-1637. [程滨, 赵永军, 张文广等. 生态化学计量学研究进展. 生态学报, 2010, 30(6): 1628-1637.]
[5] Güsewell S. N∶P ratios in terrestrial plants: Variation and functional significance.NewPhytologist, 2004, 164: 243-266.
[6] Aerts R, Chapin FS III. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns.AdvancesinEcologicalResearch, 2000, 30: 1-67.
[7] Vitousek P. Nutrient cycling and nutrient use efficiency.AmericanNaturalist, 1982, 119: 553-572.
[8] Elser JJ, Bracken ME, Cleland EEetal. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems.EcologyLetters, 2007, 10: 1135-1142.
[9] LeBauer DS, Treseder KK. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.Ecology, 2008, 89(2): 371-379.
[10] Andersen T, Elser JJ, Hessen DO. Stoichiometry and population dynamics.EcologyLetters, 2004, 7: 884-900.
[11] Scheffer M, Redelijkheid MRD, Noppert F. Distribution and dynamics of submerged vegetation in a chain of shallow eutrophic lakes.AquaticBotany, 1992, 42: 199-216.
[12] Cronin G, Lodge DM. Effects of light and nutrient availability on the growth, allocation, carbon/nitrogen balance, phenolic chemistry, and resistance to herbivory of two freshwater macrophytes.Oecologia, 2003, 137: 32-41.
[13] Cao T, Ni LY, Xie Petal. Effects of moderate ammonium enrichment on three submersed macrophytes under contrasting light availability.FreshwaterBiology, 2011, 56: 1620-1629.
[14] Barko JW, Smart RM. Sediment-related mechanisms of growth limitation in submersed macrophytes.Ecology, 1986, 67: 1328-1340.
[15] Rattray M, Howard-Williams C, Brown J. Sediment and water as sources of nitrogen and phosphorus for submerged rooted aquatic macrophytes.AquaticBotany, 1991, 40: 225-237.
[16] Chambers PA. Light and nutrients in the control of aquatic plant community structure. II. In situ observations.JournalofEcology, 1987, 75: 621-628.
[17] Xie YH, An SQ, Wu BF. Resource allocation in the submerged plantVallisnerianatansrelated to sediment type, rather than water-column nutrients.FreshwaterBiology, 2005, 50: 391-402.
[18] Sterner RW, Elser JJ, Fee EJetal. The light: nutrient relation in lakes: The balance of energy and materials affects ecosystem structure and process.AmericanNaturalist, 1997, 150: 663-684.
[19] Sterner RW, Elser JJ. Ecological stoichiometry: The biology of elements from molecules to the biosphere. Princeton:Princeton University Press, 2002.
[20] Fu H, Yuan GX, Cao Tetal. An alternative mechanism for shade adaptation: Implication of allometric responses of three submersed macrophytes to water depth.EcologicalResearch, 2012, 27: 1087-1094.
[21] Li W, Cao T, Ni LYetal. Size-dependent C, N and P stoichiometry of three submersed macrophytes along water depth gradients.EnvironmentalEarthSciences, 2015, 74: 3733-3738.
[22] Xia CX, Yu D, Wang Zetal. Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China.EcologicalEngineering, 2014, 70: 406-413.
[23] Hao Beibei, Wu Haoping, Shi Qiaoetal. Stoichiometry characteristics of submerged macrophytes in ten lakes of Yunnan plateau.JLakeSci, 2013, 25(4): 539-544. DOI: 10.18307/2013.0411. [郝贝贝, 吴昊平, 史俏等. 云南高原10个湖泊沉水植物的碳、氮、磷化学计量学特征. 湖泊科学, 2013, 25(4): 539-544.]
[24] Li Wei, He Liang, Zhu Tianshunetal. Distribution and leaf C, N, P stoichiometry ofVallisnerianatansin response to various water depths in a large mesotrophic lake, Lake Erhai, China.JLakeSci, 2014, 26(4): 585-592. DOI: 10.18307/2014.0413. [李威, 何亮, 朱天顺等. 洱海苦草(Vallisnerianatans)水深分布和叶片C、N、P化学计量学对不同水深的响应. 湖泊科学, 2014, 26(4): 585-592.]
[25] Xing W, Wu HP, Hao BBetal. Stoichiometric characteristics and responses of submerged macrophytes to eutrophication in lakes along the middle and lower reaches of the Yangtze River.EcologicalEngineering, 2013, 54: 16-21. DOI: 10.1016/j.ecoleng.2013.01.026.
[26] Li W, Cao T, Ni LYetal. Effects of water depth on carbon, nitrogen and phosphorus stoichiometry of five submersed macrophytes in an in situ experiment.EcologicalEngineering, 2013, 61: 358-365.
[27] Bao Shidan ed. Soil agro-chemistrical analysis (3rd edition). Beijing: China Agriculture Press, 2000. [鲍士旦. 土壤农化分析(第三版). 北京:中国农业出版社, 2000.]
[28] Huang Xiangfei, Chen Weimin, Cai Qiming eds. Survey, observation and analysis of lake ecology:Standard methods for observation and analysis in Chinese Ecosystem Research Network Series V. Beijing: Standards Press of China, 1999. [黄祥飞, 陈伟民, 蔡启铭. 湖泊生态调查观测与分析: 中国生态系统研究网络观测与分析标准方法. 北京:中国标准出版社, 1999.]
[29] Krause-Jensen D, Sand-Jensen K. Light attenuation and photosynthesis of aquatic plant communities.LimnologyandOceanography, 1998, 43: 396-407.
[30] Manly BFJ ed. Randomization, bootstrap and Monte Carlo methods in biology. Boca Raton: CRC Press, 2006.
[31] R Core Team (2014). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http: //www. R-project. org/.
[32] Zhao Haichao, Wang Shengrui, Jiao Lixinetal. Characteristics of temporal and spatial distribution of the nitrogen forms in the sediments of Erhai Lake.ResearchofEnvironmentalSciences, 2013, 26(3): 235-242. [赵海超, 王圣瑞, 焦立新等. 洱海沉积物中不同形态氮的时空分布特征. 环境科学研究, 2013, 26(3): 235-242.]
[33] Zhao Haichao, Wang Shengrui, Jiao Lixinetal. Characteristics of temporal and spatial distribution of different forms of phosphorus in the sediments of Erhai Lake.ResearchofEnvironmentalSciences, 2013, 26(3): 227-234. [赵海超, 王圣瑞, 焦立新等. 洱海沉积物中不同形态磷的时空分布特征. 环境科学研究, 2013, 26(3): 227-234.]
[34] Yuan GX, Fu H, Zhong JYetal. Growth and C/N metabolism of three submersed macrophytes in response to water depths.EnvironmentalandExperimentalBotany, 2016, 122: 94-99.
[35] Wu Aiping, Wu Shikai, Ni Leyi. Study of macrophytes nitrogen and phosphorus contents of the shallow lakes in the middle reaches of Changjiang River.ActaHydrobiologicaSinica, 2005, 29: 406-412. [吴爱平, 吴世凯, 倪乐意. 长江中游浅水湖泊水生植物氮磷含量与水柱营养的关系. 水生生物学报, 2005, 29: 406-412.
Community level stoichiometric characteristics of submerged macrophytes and their influencing factors in the mid-lower Yangtze lakes
SU Haojie1,2, WU Yao1,2, XIA Wulai1,2, XIE Ping1**& CAO Te1
(1:DonghuExperimentalStationofLakeEcosystems,StateKeyLaboratoryofFreshwaterEcologyandBiotechnology,InstituteofHydrobiology,ChineseAcademyofSciences,Wuhan430072,P.R.China)(2: University of Chinese Academy of Sciences, Beijing 100049, P.R.China)
Toresearchcommunitylevelstoichiometriccharacteristicsofsubmergedmacrophytesandtheirinfluencingfactors,14lakesalongthemid-lowerreachesofYangtzeRiverwereinvestigatedfromJunetoAugustin2014.Resultsshowedthattheaveragetotalcarbon(C)contentsofthecommunitieswere386.93±25.80mg/gandtheconcentrationsoftotalCrangedfrom315.98to441.97mg/g.Theaveragetotalnitrogen(N)andtotalphosphorus(P)contentswere26.10±4.84mg/gand2.64±0.99mg/g,respectively,andtheconcentrationsoftotalNandPrangedfrom13.75to40.89mg/gand1.01to5.92mg/g,respectively.Comparedwiththeresearchesatspecieslevel,variationcoefficientsofstoichiometriccharacteristicsatcommunitylevelinourstudyweremuchlower,indicatingthatcommunitylevelstoichiometrichomeostasiswasstrongerthanthatofspecieslevel.CommunityCcontentsweresignificantcorrelatedwithPcontents,whilehadnorelationshipwithN,indicatingthatcouplingrelationsbetweenelementCandnutrientelementswereinconsistent.CommunityN∶PweresignificantlycorrelatedwithPcontents,whilehadnorelationshipwithN,indicatingthatN∶PwerecontrolledbyPcontentsratherthanbyN.Redundancyanalysis(RDA)resultshowedthatenvironmentalfactorsexplained30.2%oftotalvariationofstoichiometricsignatures.SedimentP,sedimentN∶P,waterdepthandlightattenuationcoefficienthadsignificantimpactsoncommunitystoichiometricsignatures.Resultsofvariancepartitioningshowedthatsedimentandwatercolumnexplained5.21%and10.19%oftotalvariance,respectively.Theinteractioneffectsofsedimentandwatercolumnexplained5.25%oftotalvariance.Ourresultssuggestthatthecontroloflightconditionsofwatercolumnismoreurgentthanthatofsedimentinlakerecoveryofsubmergedvegetation.
Submergedmacrophytecommunity;ecologicalstoichiometry;themid-lowerYangtzelakes;sediment;watercolumn
*淡水生态与生物技术国家重点实验室项目(2014FBZ02)资助. 2016-05-08收稿;2016-06-07收修改稿.苏豪杰(1989~),男,博士研究生;E-mail: hdsk1129@126.com.
J.LakeSci.(湖泊科学), 2017, 29(2): 430-438
DOI 10.18307/2017.0219
©2017 byJournalofLakeSciences
**通信作者;E-mail: xieping@ihb.ac.cn.
猜你喜欢
河北水利(2022年10期)2022-12-29
海洋通报(2022年4期)2022-10-10
海洋信息技术与应用(2022年1期)2022-06-05
湖泊科学(2021年4期)2021-07-07
园林(2020年8期)2020-09-18
皮革制作与环保科技(2020年14期)2020-03-17
花卉(2020年12期)2020-01-09
——以武汉紫阳湖为例
绿色科技(2019年2期)2019-05-21
浙江大学学报(工学版)(2016年9期)2016-06-05
铜业工程(2015年4期)2015-12-29
