
不同林龄胡杨活立木枯枝生物量和化学计量特征
2017-04-10 12:31:03史军辉刘茂秀王新英马学喜
生态学报 2017年3期
史军辉,刘茂秀,王新英,马学喜
新疆林科院造林治沙研究所, 乌鲁木齐 830046
不同林龄胡杨活立木枯枝生物量和化学计量特征
史军辉*,刘茂秀,王新英,马学喜
新疆林科院造林治沙研究所, 乌鲁木齐 830046
基于不同林龄胡杨天然林样地活立木枯枝(DBST)生物量调查,探讨不同发育阶段胡杨活立木枯枝化学计量和碳储量的时空变化规律。研究结果表明:(1)不同林龄胡杨林活立木枯枝生物量随着林龄增加呈先增加后减小趋势,其中成熟林活立木枯枝生物量最大(10.93 t/hm2),占地上生物量的17.79%。对于单株胡杨而言,随林龄增加活立木枯枝生物量增大,年平均活立木枯枝生物量为0.22 kg/株。同时胡杨枝径级越大越易形成枯枝,生物量也随枯枝径级的增加而增加。(2)胡杨活立木枯枝C、N、P、K平均含量分别为491.01、4.13、2.75、1.83 g/kg,活立木枯枝C、N、P元素含量高于活体枝,而活体枝K元素含量高于活立木枯枝。胡杨群落活立木枯枝平均C∶N、C∶P、N∶P比分别为:128.58、232.79、1.95,其活立木枯枝化学计量比均高于活体枝。林龄对化学元素含量和化学计量比影响均差 异不显著(P>0.05),说明了胡杨活立木枯枝的元素含量随龄级变化不大,具有稳定的特点。(3)活立木枯枝碳密度随林龄的增加先增加后降低,其中成熟林碳密度最大;胡杨群落平均碳密度为2.63 t/hm2,占地上活立木碳密度的20.74%。研究认为胡杨群落枯枝生物量和碳密度随林龄增加的变化规律明显,胡杨活立木枯枝在荒漠森林生态系统中是不可忽略的C库。
胡杨;活立木枯枝;生物量;化学计量
森林木质残体是森林生态系统中重要的结构性和功能性组成要素,是组成森林生态系统食物网结构、空间结构的重要单元,也是联系森林生态系统养分循环、碳库贮存、群落更新以及为其它有机体提供生境等主要功能的载体和纽带[1- 6]。木质物残体在保持森林生态系统的完整性方面发挥着重要的生态功能,影响着系统内外相关的生物和非生物过程,不仅为微生物、节肢动物、鸟类及哺乳动物提供栖息环境,也是生态系统中重要的碳库和养分库[7]。木质物残体包括粗木质残体(CWD,枝径≥10 cm)和细木质残体(FWD,1 cm≤枝径≤10 cm)[8],其中粗木质残体主要由倒木、枯立木枯桩和枯落大枝组成,是森林木质物残体的主体;细木质残体主要指小枝,其数量和生态功能相对次要。
随着全球变暖的日趋严重,碳循环研究也越来越向区域尺度发展,准确量化区域森林碳储量及其动态十分重要[9]。目前,对森林碳库的估算主要以活立木植被与土壤的碳储量为主,然而,森林枯立木及枯枝的碳储量的作用也引起学者的重视。李凌浩等[10- 11]对秦岭巴山冷杉林和武夷山甜槠林、杨礼攀[12]对哀牢山山地湿性常绿阔叶林、闫恩荣等[8]对常绿阔叶林的CWD的贮量、动态和功能进行了研究,蔡慧颖等[13]研究了小兴安岭谷地云冷杉林粗木质残体碳密度特征,唐旭利等[14]相继开展了鼎湖山季风常绿阔叶林CWD碳贮量研究,均表明CWD碳储量较高,对森林生态系统碳平衡具有很重要的意义。因此,枯枝碳储量在森林碳库中具有重要地位。
荒漠森林生物量是荒漠生态系统的基本数量特征,是研究荒漠生态系统物质循环、能量流动及荒漠生态系统固碳能力的基本要素[15-16]。新疆塔里木河流域胡杨林(Populuseuphratica)是世界分布面积最大,群落类型最多,生境条件复杂的胡杨森林生态系统,构成塔里木河流域、和田河流域、叶尔羌河等大河流域绿色屏障的主体,生态地位十分重要,对其的深入研究既可以评价荒漠生态系统生产潜力,也可为荒漠生态系统经营管理提供必需的基础数据。目前,新疆胡杨生物量方面的研究主要集中在胡杨生物量及其变化规律、生物量预测方法、模型以及与其他相关环境因子之间关系方面[17- 19],而有关胡杨木质残体研究较少。胡杨活立木宿存的枯枝是胡杨森林群落明显的特征之一,是对干旱高温等环境胁迫的重要的适应策略。活立木枯枝广泛分布于胡杨不同群落,是胡杨森林生态系统的物质与能量循环的重要组成。从野外调查分析,胡杨活立木枯枝径级有不同尺寸,既可以将其划入CWD,也能划入FWD,便于研究,把活立木上宿存的不同径级枯枝统称为活立木枯枝(DBST),它是木质残体的一种特殊表现形式。开展该领域的研究,对丰富胡杨林木质残体在物质和能量循环中的功能与作用具有重要的科学意义。
本文以塔河流域胡杨群落活立木的枯枝为研究对象,研究不同林龄胡杨活立木枯枝生物量、化学计量的特征及碳储量的变化,旨在为揭示胡杨生态系统物质循环过程规律提供科学数据,为进一步了解和研究胡杨活立木枯枝形成机制及胡杨群落演替,同时也为制定胡杨林合理经营、永续利用提供科学依据。
1 研究区概况
研究区域位于新疆巴音郭楞蒙古自治州轮台县轮南镇塔里木河胡杨林自然保护区(84°20′E,41°10′N,海拔912 m)。该区东西长109.7 km,南北宽47.1 km,是全世界原始胡杨林面积最大,分布最集中,保存最完整的地区。该区属典型温带荒漠气候,年平均气温10.9 ℃,夏季平均气温26.8 ℃;年积温(≥10℃)3900—4300 ℃,无霜期180—224 d;多年平均降水量为65.5 mm,降水主要集中在5—8月,占全年降水总量的94.4%;年平均蒸发量2024 mm,年平均相对湿度为35%—55%,平均风速1.8 m/s。该地区草本植物主要有芦苇(Phragmitescommunis)、胀果甘草(Glycyrrhizainflata)、骆驼刺(Alhagisparsifolia)等,灌木主要以多枝柽柳(Tamarixramosissima)、铃铛刺(Halimodendronhalodendron)为主,胡杨为该地区乔木层主要建群种,它们共同构成了塔里木河中游荒漠河岸群落,研究区土壤类型主要为风沙土。
2 材料和方法
2.1 样品的采集
2013年9月初在塔里木河流域胡杨林自然保护区进行野外调查,选取群落特征相近、立地条件基本一致的不同林龄的天然胡杨林为研究对象。胡杨龄级的划分参照王世绩等[20]对胡杨的研究结果,分为幼龄林、中龄林、近熟林、成熟林和过熟林。每个龄级设置3个标准样地,标准样地规格为100 m×100 m的正方形,四角以管径为3 cm的PVC管定点,四边拉线。采用每木检尺的方法对胸径>1.0 cm的胡杨进行调查,测定指标为:胸径、树高、冠幅、群落郁闭度等林分特征(表1)。每个样地按龄级,以平均高度与胸径,选择3株发育完整的胡杨,采集枯枝和活体枝鲜样,枝按径级<1 cm、1—2 cm、2—4 cm和>4 cm采集适量,每个标准株按不同枝径级将样品装袋,放置便携式移动冰箱,带回实验室分析化学元素含量。每个龄级以平均高度与胸径,选择3株标准株,将标准株从其根颈处锯伐,锯伐后收集枯枝的生物量,枯枝生物量按径级<1 cm、1—2 cm、2—4 cm和 >4 cm采集。同时将树干在树高1.3 m处分段,其后按2 m长度分段锯断称重并取圆盘,分别测定其干、枝、叶、皮的新鲜生物量。将鲜样带回实验室杀青后,后置于80℃烘箱中烘至质量恒定用以计算胡杨地上生物量。
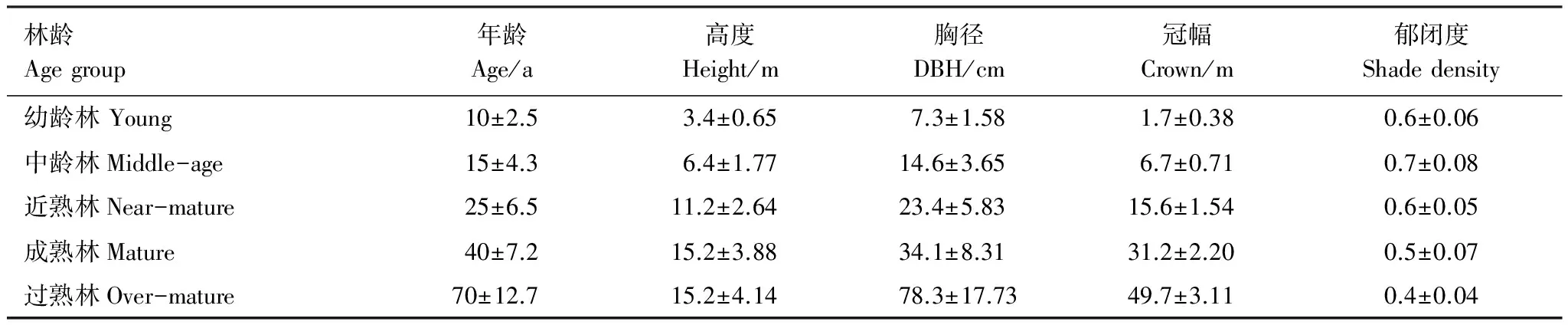
表1 林分基本概况
2.2 样品的处理及测定
采集的样品带回实验室,鲜样杀青后,与枯枝置于80℃烘箱中烘至质量恒定,磨碎、过筛后装瓶待用。有机碳和全氮测定分别采用重铬酸钾容量法-外加热法(GB 9834- 88)和半微量开氏法(GB 7173- 87),全磷采用酸溶法(GB 9837- 88),全钾采用火焰光度法。
2.3 数据分析
运用Excel 2010将数据进行处理,利用SPSS 16.0 one-way ANOVA进行单因素方差分析,采用Duncan新复极差法在显著性水平为α=0.05下比较各指标不同林龄的差异性。
3 结果与分析
3.1 单株胡杨活立木枯枝生物量变化
3.1.1 不同枝径枯枝生物量特征
不同枝径枯枝生物量均随林龄的增加而增加(图1)。就枯枝径级而言,不同林龄<1 cm的枯枝差异不显著,其中成熟林枯枝的生物量最大(7.15 kg),幼龄林枯枝最小(0.35 kg),差值达6.80 kg。1—2 cm径级枯枝中,过熟林枯枝生物量为12.80 kg,是幼龄林的28.44倍。方差分析,过熟林枯枝生物量与幼龄林、中熟林、近熟林差异显著(P<0.05)。不同林龄的2—4 cm枯枝生物量差异显著,过熟林生物量高达20.60 kg,是幼龄林枯枝生物量的41.20倍。方差分析,幼龄林、成熟林和过熟林枯枝生物量差异显著(P<0.05)。在幼龄林和中熟林中,没有>4 cm的枯枝;近熟林到过熟林出现>4 cm的枯枝,不同林龄的枯枝生物量差异不显著,依然是过熟林枯枝生物量最大,是近熟林枯枝生物量的2.45倍。
相同林龄单株胡杨,枯枝生物量随径级的增加而增加。幼龄林枯枝变化不明显,最大的径级(2—4 cm)生物量为0.50 kg,与最小径级(<1 cm)生物量相差0.15 kg。中龄林枯枝变化也不明显,最大的径级(2—4 cm)生物量为1.30 kg,与最小径级(<1 cm)生物量差值较小(0.50 kg)。近熟林和过熟林>4 cm枯枝生物量分别是<1 cm枯枝生物量的5.17倍和4.36倍。成熟林不同径级枯枝生物量从大到小的顺序依次为:>4 cm、2—4 cm、<1 cm、1—2 cm。结果表明:近熟林、成熟林和过熟林容易形成>4 cm径级的枯枝,而幼龄林和中龄林容易形成2—4cm径级的枯枝。
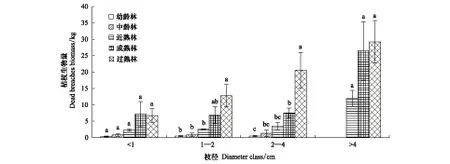
图1 不同径级枯枝生物量的变化Fig.1 Change of dead branches biomass in different diameter class (mean±S.E.)不同小写字母间表示差异显著
3.1.2 不同林龄枯枝生物量特征
枯枝生物量和地上生物量均随林龄的增加而增加(表2),林龄对单株枯枝生物量影响差异显著(P<0.05)。统计分析,幼龄林和中龄林单株枯枝生物量差异不显著,差值为1.65 kg,但均与其他林龄枯枝生物量差异显著;其中过熟林与幼龄林枯枝生物量差异较大(68.00 kg)。过熟林枯枝生物量是中龄林、近熟林、成熟林枯枝生物量的23.49、3.42、1.46倍。不同林龄地上生物量差异显著(P<0.05),过熟林地上生物量分别是幼龄林、中龄林、近熟林、成熟林生物量的62.70、21.70、5.54、1.51倍。分析发现,枯枝生物量占地上生物量的比例在13.72%—21.79%之间,平均为17.10%,其中近熟林的枯枝比例最大(21.79%),中龄林比例最小(13.72%)。因此,胡杨活立木枯枝占据了地上生物量较大的比例,其生物量是胡杨生态系统中重要的碳库组成。

表2 不同林龄枯枝生物量变化及比例
不同小写字母代表不同林龄间的差异显著
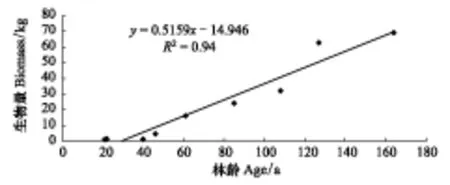
图2 枯枝生物量与林龄的关系 Fig.2 The relationship between dead branches biomass and forest age
通过对解析木的年轮进行推算,得出枯枝生物量年累计率随着林龄增加(图2)。枯枝生物量与林龄呈正相关(y=0.5159x-14.946,R2=0.94)。通过计算得知单株胡杨年平均枯枝生物量为0.22 kg/株。
3.2 胡杨群落活立木枯枝生物量特征
不同林龄胡杨群落枯枝生物量和地上生物量变化规律相同(表3)。随着胡杨群落林龄的增加,林分的密度逐渐减小,而枯枝和地上活体群落生物量均随林龄的增加先增加后减小,在成熟林达到了最大值(10.93 t/hm2和61.61 t/hm2)。成熟林群落枯枝生物量分别是幼龄林、中龄林、近熟林和过熟林群落枯枝生物量的11.04、8.67、2.02、1.33倍。过熟林群落枯枝生物量次之(8.20 t/hm2),是近熟林的1.51倍。从群落枯枝生物量占地上活体生物量的比例来看,近熟林枯枝生物量比例最大(27.85%),最小的是中龄林(15.90%),差值较大(11.95%),过熟林和成熟林枯枝生物量比例趋于稳定。因此,胡杨群落枯枝生物量占地上活体生物量比例较高(平均19.79%),是胡杨生态系统中重要的碳库与养分库。
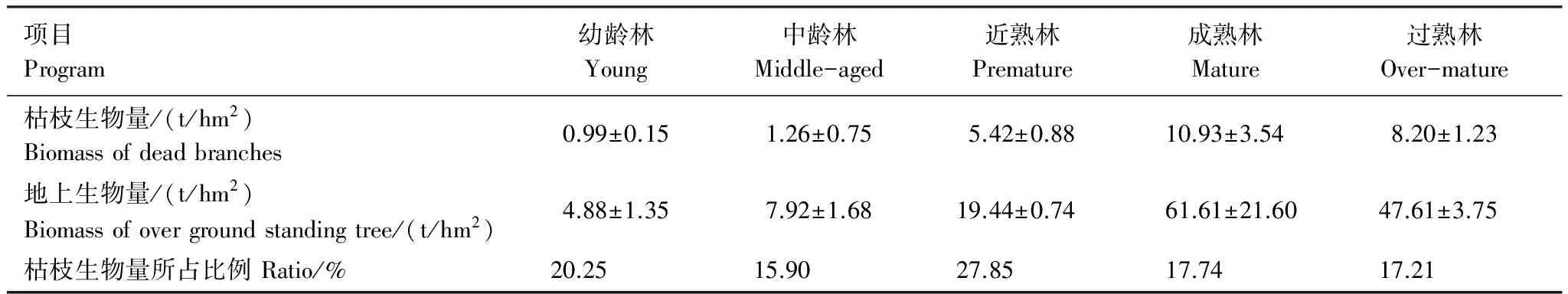
表3 不同林龄胡杨群落枯枝生物量变化
3.3 不同林龄活立木枯枝元素含量及化学计量变化
3.3.1 不同林龄枯枝元素含量变化
枯枝有机碳的含量在476.42—497.90 g/kg(表4),统计分析,各龄级间差异不显著,其中:中龄林枯枝C含量最高,成熟林最低,差值为21.48 g/kg。枯枝N含量在3.64—5.00 g/kg,总体而言,各龄级间差异不显著,仅中龄林和近熟林N含量差异显著;近熟林N含量最大,中龄林N含量最小,差值为1.36 g/kg。枯枝P含量在1.74—3.74 g/kg,统计分析,各龄级间差异不显著,但枯枝P含量随着林龄的增加而持续降低。枯枝K含量在0.87—2.59 g/kg,方差分析,林龄对活立木枯枝元素含量影响均差异不显著(P>0.05),但枯枝K含量具有随着林龄增加而增加的趋势。
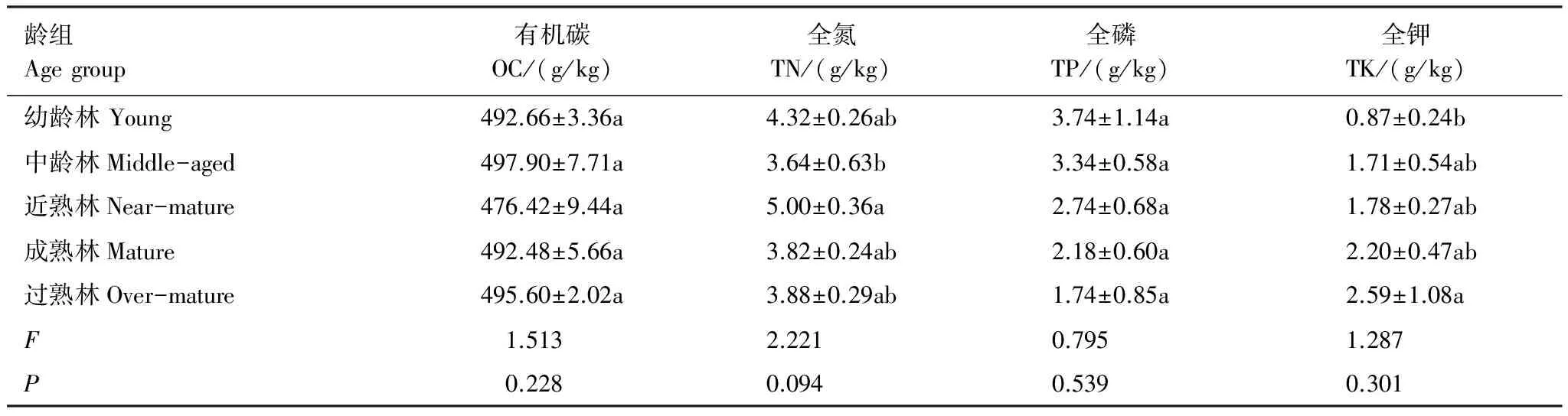
表4 不同林龄枯枝C、N、P、K含量变化
胡杨群落活立木枯枝C、N、P、K平均含量分别为:491.01、4.13、2.75、1.83 g/kg。不同林龄枯枝K含量变异系数最大(0.35),枯枝P含量变异系数次之(0.30),而枯枝C含量变异系数最小(0.02),表明不同林龄活立木枯枝C含量比较稳定,P和K含量波动性变化。枯枝与活体枝相比,枯枝的有机质、全氮、全磷含量都明显高于活枝的含量(表5),但差值不大,枯枝的有机质、全氮、全磷含量是活枝的1.06、1.03、1.18倍;而全钾则表现为活体枝高于枯枝。

表5 枯枝和活枝的C、N、P、K含量对比
3.3.2 不同林龄枯枝化学计量的变化
从表6可以看出,不同林龄枯枝C∶N比在99.19—170.34,平均C∶N比为128.58(表6),变异系数为0.20;枯枝C∶N比随林龄的增加先增加后减小,其中近熟林C∶N比最小,中龄林最大。不同林龄枯枝C∶P比在164.73—304.12,平均C∶P比为232.79,变异系数为0.26;C∶P比随林龄增加呈先减小后增加又降低的趋势,其中中龄林C∶P比最小,成熟林最大,中龄林和成熟林C∶P比差异显著(P<0.05)。不同林龄枯枝N∶P比在1.25—2.35,不同林龄平均N∶P比为1.95,变异系数为0.25,N∶P比随林龄呈波动趋势。方差分析,林龄对活立木枯枝化学计量比影响均不显著(P>0.05)。
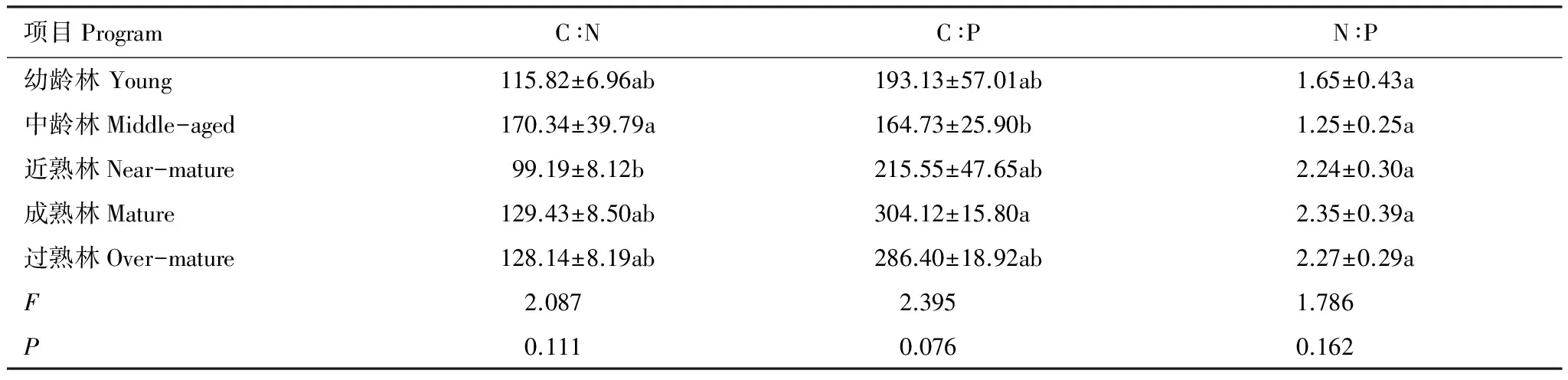
表6 不同林龄枯枝化学计量变化
由表7可知,不同林龄枯枝平均C∶N、C∶P、N∶P分别为:128.58、232.79、1.95,化学计量比变异系数基本接近,分别为:0.20、0.26、0.25。枯枝C∶N、C∶P、N∶P比均高于活枝的计量比,但差异不显著。

表7 枯枝与活枝化学计量对比
3.4 不同林龄枯枝碳密度特征
将不同林龄的胡杨枯枝及地上活体生物量与其碳素含量的相乘,求出不同年龄阶段胡杨群落枯枝和地上活体的碳密度。从表8看出,胡杨群落枯枝碳密度随林龄增加先增加后降低,在成熟林达到了最大值(5.38 t/hm2),是过熟林枯枝碳密度的1.33倍,而幼龄林最小(0.49 t/hm2),分别是成熟林和过熟林碳密度的0.09和0.12倍。胡杨群落枯枝碳密度平均值为2.63 t/hm2。地上活体碳密度与枯枝碳密度具有相同规律,成熟林碳密度最大(30.26 t/hm2)。从枯枝碳密度占地上活体碳密度比例来看,近熟林最大(27.90%),不同林龄其所占的比例依次为:近熟林> 幼龄林> 过熟林> 成熟林> 中龄林,其平均比例为20.74%。通过对新疆南部平原区胡杨不同林龄面积进行统计,计算出不同林龄活立木枯枝碳储量(表8),成熟林碳储量最大(155520.05 t),幼龄林最小(29023.63 t),近熟林碳储量是幼龄林的5.36倍,胡杨林活立木枯枝碳储量总计为435125.83 t,并且年平均碳储量为31370.63 t。表明胡杨活立木枯枝贮碳量在胡杨森林生态系统碳素流动中具有重要的地位。
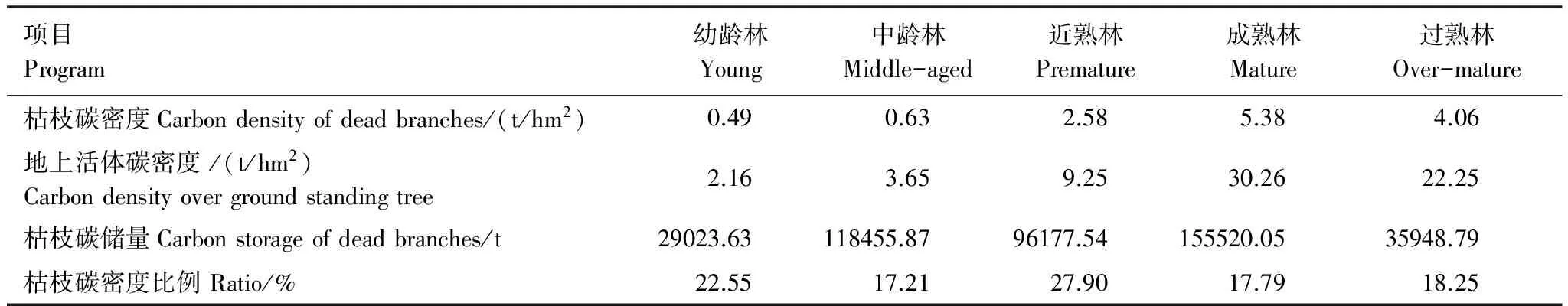
表8 不同林龄胡杨群落枯枝碳储量变化
4 讨论
4.1 胡杨活立木枯枝特征
生物量累积是森林有机碳累积的主要方式[21-22],以枯立木、倒木、枯落大枝和死根等形式存在的木质物残体是森林生态系统中重要的碳库和养分库。胡杨林是新疆干旱荒漠区域自然成林的乔木森林类型,乔木层仅为胡杨或灰胡杨(Populuspruinosa),其广泛分布于塔里木河流域、叶尔羌河流域和和田河流域。胡杨森林群落除了具有枯立木、地上枯枝等粗质残体类型外,活立木上还存在相当数量的枯枝,这一特殊现象使胡杨生态系统在物质循环过程丰富了木质物残体类型。
生态系统不同组分生物量大小可以表征其在生态系统中的重要性,据估计森林生态系统中CWD约占地上生物量的2%—10%[23]。据估计,世界上天然针叶林中粗木质残体的储量可达30—200 t/hm2,美国西北部奥林匹克国家公园的黄杉(Pseudotsugasinensis) -铁杉(Tsugachinensis) 林中粗木质残体最高可达537 t/hm2,阔叶林(含针阔混交林) 为8—50 t/hm2[24]。研究表明,我国不同森林类型中也含有大量的粗木质残体,西南哀牢山中山湿性常绿阔叶林的粗木质残体为98. 46 t/hm2[12],暖温带与亚热带过渡带的秦岭巴山冷杉(Abiesfargesii) 林中粗木质残体储量为15. 85 t/hm2[10],南亚热带鼎湖山季风常绿阔叶林CWD的生物量为42.09 t/hm2[25]。本文研究表明,胡杨成熟林群落活立木枯枝生物量为10.93 t/hm2,与其他森林粗木质残体生物量比较,低于秦岭巴山冷杉林、鼎湖山季风常绿阔叶和哀牢山中山湿性常绿阔叶林,与大兴安岭森林CWD储量11.63 t/hm2相近[26];但与其枝木质残体生物量比,远高于哀牢山中山湿性常绿阔叶林6.00 t/hm2[12],鼎湖山季风常绿阔叶林4.65 t/hm2[14]。胡杨群落枯枝生物量平均值为5.36 t/hm2,其枯枝生物量占地上活体生物量的19.79%。因此,胡杨活立木枯枝生物量在极端干旱环境森林生态系统中物质与能量循环过程具有重要的意义。
从寒温带到热带森林,森林中CWD的来源主要有3个方面,1)森林中林木生长竞争排斥和老龄林自然死亡;2)自然干扰(主要是风、雨、雪、火灾及病虫害等)导致的林木倒伏;3)人为干扰(伐木、拾樵等)[27- 28],因此,竞争与干扰是寒温带森林到热带森林产生CWD普遍原因。而对于生存在极端干旱环境下的荒漠河岸林胡杨群落,由于周期性旱湿交替过程及地下水位季节性动态变化,在其生长季面临高温干燥的大气与干旱的土壤条件,水分亏缺迫使胡杨活立木形成枯枝以减少个体水分蒸腾和水分需求,是胡杨群落在极端环境条件下的一种生存策略,因此,胡杨活立木枯枝形成机制及成因具有主动性和特殊性,其应有复杂的生理生化过程的响应。从林分发育阶段分析,从寒温带到热带森林,由于密度效应,森林中较小的树木受到竞争、压抑,其直径、高度都生长缓慢,容易形成CWD,其储量较高;过熟林树木的中空和自然死亡是大径级林木形成CWD的主要途径;成熟林中CWD的数量最低。因此,有学者推论出林分在整个生长发育过程CWD数量累计呈“U”形[29]。作为CWD一种特殊类型,本研究表明,随林龄增加,胡杨活力木枯枝碳密度呈现增加趋势,幼龄林最小(0.49 t/hm2),在成熟林达到了最大值(5.38 t/hm2),过熟林有所降低(4.06 t/hm2),从林分发育过程分析,胡杨活立木枯枝形成机制与前人研究有所不同。因此,胡杨活立木枯枝作为一种特殊的木质残体类型,其形成机制及成因需要深入研究。
4.2 胡杨活立木枯枝碳汇的潜力
化石燃料燃烧产生的二氧化碳,部分存储于大气(导致大气二氧化碳浓度升高),部分进入海洋,但还有一部分不知去向,即所谓的“迷失碳汇”。森林是陆地最大的贮碳库,全球陆地总碳库为1800 Pg,其中森林的碳储量约占陆地生物圈地上碳储量的80%和地下碳储量的40%[30]。我国森林年均吸收的二氧化碳占生物固碳总量的80%,在维护全球气候系统、调节全球碳平衡、减缓大气温室气体上升等方面具有不可替代的作用[31-32]。李彦等[33]推测荒漠区很可能是一个很大的碳汇,初步估计地下碳库总量(全球)高达1000亿t,是陆地上植物、土壤之外的第三个活动碳库,为寻找“迷失碳汇”开创了一个全新的方向。粗木质残体作为森林生态系统中非常重要的营养库和碳库,占地上有机物质储量的1%—45%[34]。胡杨生活在极端干旱区,活枝形成枯枝时,枯枝固定了部分碳形成树上碳库。由于胡杨处于特殊的生存环境和胡杨木质的特殊性,其枯枝不易腐烂分解,从而形成不活动碳库,这与胡杨林中凋落的枯枝形成的土壤碳库在分解速率等方面存在明显的差异,值得探讨。据估算,胡杨林枯枝碳储量高达435125.83 t,具有较大储碳能力,是区域生态系统服务价值的评价中不可缺失的指标[35]。胡杨活立木枯枝碳储量比较高,但在干旱区碳汇计量中往往被忽略,枯枝可能占据了部分“迷失碳汇”。
4.3 胡杨枯枝化学计量特征
生态化学计量学是研究生态系统中能量与化学元素间平衡的科学,主要强调的是有机体的主要组成元素(特别是C、N和P)的关系,化学计量值可以反映生物进化的结果,主要包括影响生物生长速率、健康状况、新陈代谢、组织结构和生态演替等的因素[36]。因此,化学计量主要研究活体叶片,有关枯死的生物化学计量未见报道。在本研究中,通过研究枯枝化学元素含量和化学计量,与活体枝进行了比较。研究表明,除钾元素活体枝高于枯枝外,其他元素含量和化学计量均高于活体枝的,但两者差值不大。枯枝和活体枝C、N、P和K差值为26.31、0.13、0.21 g/kg和0.76 g/kg,枯枝和活体枝C∶N、C∶P、N∶P差值分别为:10.72、5.64、0.05。枯枝的元素含量和化学计量变化与植物活体表现出一致的规律,这与枯枝秉承了植物活体枝的特性密切相关[37]。胡杨龄级对活立木枯枝元素含量和化学计量影响不显著,这说明胡杨枯枝的C、N、P、K含量具有稳定的特点。
5 结论
胡杨单株枯枝生物量随林龄增加而增大,年平均枯枝累计生物量为0.22 kg/株。随着枯枝径级的增加,生物量也随之增加。不同林龄群落枯枝生物量随着林龄增加,先增加后减小,在成熟林枯枝生物量最大,枯枝生物量占地上生物量17.79%。除钾元素外,其他元素活立木枯枝均高于活体枝,活立木枯枝的化学计量均高于活体枝。林龄对化学元素含量及化学计量比影响不显著,说明活立木枯枝化学元素含量具有稳定性特点。胡杨群落枯枝碳密度平均值为2.63 t/hm2。枯枝碳密度随林龄增大先增加后降低,成熟林碳密度最大(5.38 t/hm2)。枯枝碳密度占地上活体碳密度平均比例为20.74%。
总体来说,胡杨群落枯枝生物量和碳储量随林龄增加具有明显变化规律,碳汇潜力巨大。因此,活立木枯枝是胡杨森林生态系统中重要的结构性和功能性组成要素,是联系森林生态系统养分循环、碳库储存及为其他有机体提供生境等主要功能的载体,对森林生态系统物质循环以及在维护森林生态系统的完整性和稳定性方面具有重要作用。胡杨活立木上的枯枝碳储量研究对未来我国荒漠区森林的可持续发展和碳汇潜力的增加都具有重要的意义,其活立木枯枝形成机制需进一步深入研究。
[1] Harmon M E, Franklin J F, Swanson F J, Sollins P, Gregory S V, Lattin J D, Anderson N H, Cline S P, Aumen N G, Sedell J R, Lienkaemper G W, Cromack K Jr, Cummins K W. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research, 1986, 15: 133- 302.
[2] Sturtevant B R, Bissonette J A, Long J N, Roberts D W. Coarse woody debris as a function of age, stand structure, and disturbance in boreal Newfoundland. Ecological Applications, 1997, 7(2): 702- 712.
[3] Woodall C W, Walters B F, Oswalt S N, Domke G M, Toney C, Gray A N. Biomass and carbon attributes of downed woody materials in forests of the United States. Forest Ecology and Management, 2013, 305: 48- 59.
[4] Sullivan T P, Sullivan D S. Voles, trees, and woody debris structures as habitat: Balancing forest crop protection and biodiversity. Crop protection, 2014, 60: 70- 77.
[5] Iwashita D K, Litton C M, Giardina C P. Coarse woody debris carbon storage across a mean annual temperature gradient in tropical montane wet forest. Forest Ecology and Management, 2013, 291: 336- 343
[6] 郝占庆, 吕航. 木质物残体在森林生态系统中的功能评述. 生态学进展, 1989, 6(3): 179- 183.
[7] 杨礼攀. 哀牢山山地湿性常绿阔叶林木质物残体的贮量、组成和生态学功能研究[D]. 昆明: 西双版纳热带植物园, 2007.
[8] 闫恩荣, 王希华, 黄建军. 森林粗死木质残体的概念及其分类. 生态学报, 2005, 25(1): 158- 167.
[9] 张修玉, 管东生, 张海东. 广州三种森林粗死木质残体(CWD)的储量与分解特征. 生态学报, 2009, 29(10): 5227- 5236.
[10] 李凌浩, 党高弟, 汪铁军, 赵雷刚. 秦岭巴山冷杉林粗死木质残体研究. 植物生态学报, 1998, 22(5): 434- 440.
[11] 李凌浩, 王其兵, 邢雪荣. 森林生态系统研究中几个重要方面的进展. 植物学通报, 1998, 15(1):17- 26.
[12] 杨礼攀, 刘文耀, 杨国平, 马文章, 李达文. 哀牢山湿性常绿阔叶林和次生林木质物残体的组成与碳贮量. 应用生态学报, 2007, 18(10): 2153- 2159.
[13] 蔡慧颖, 邸雪颖, 金光泽. 小兴安岭谷地云冷杉林粗木质残体碳密度特征. 生态学报, 2015, 35(24): 8194- 8201.
[14] 唐旭利, 周国逸, 周霞, 温达志, 张倩媚, 尹光彩. 鼎湖山季风常绿阔叶林粗死木质残体的研究. 植物生态学报, 2003, 27(4): 484- 489.
[15] 杨昊天, 李新荣, 刘立超, 贾荣亮, 王增如, 李小军, 李刚. 荒漠草地4种灌木生物量分配特征. 中国沙漠, 2013, 33(5): 1340- 1348.
[16] Cai T J, Ju C Y, Yang X H. Comparison of ridge regression and partial least squares regression for estimating above-ground biomass with landsat images and terrain data in Muus sandy land, China. Arid Land Research and Management, 2009, 23(3): 248- 261.
[17] 曹德昌, 李景文. 胡杨种群生殖生态学研究进展. 科学技术与工程, 2009, 9(5): 1202- 1209.
[18] 张昊, 李景文, 李俊清, 张玉波, 孙立. 额济纳绿洲胡杨地上部分建构模式的研究. 西北林学院学报, 2009, 24(5): 46- 49.
[19] 崔德宝, 于瑞德, 于洋, 王思奇. 新疆艾比湖自然保护区阿奇克苏河沿岸胡杨年龄结构和生长量状况研究. 干旱区资源与环境, 2012, 26(1): 194- 200.
[20] 王世绩, 陈炳浩, 李护群. 胡杨林. 北京: 中国环境科学出版社, 1995.
[21] 唐守正, 张会儒, 胥辉. 相容性生物量模型的建立及其估计方法研究. 林业科学, 2000, 36(S1): 19- 27.
[22] Richter D D, Markewitz D, Trumbore S E, Wells C G. Rapid accumulation and turnover of soil carbon in a re-establishing forest. Nature, 1999, 400(6739): 56- 58.
[23] Delaney M, Brown S, Lugo A E, Torres-Lezama A, Quintero N B. The quantity and turnover of dead wood in permanent forest plots in six life zones of Venezuela. Biotropica, 1998, 30(1): 2- 11.
[24] 王洪岩, 王文杰, 邱岭, 苏冬雪, 安静, 郑广宇, 祖元刚. 兴安落叶松林生物量、地表枯落物量及土壤有机碳储量随林分生长的变化差异. 生态学报, 2012, 32(3): 833- 843.
[25] 杨方方, 李跃林. 鼎湖山粗死木质残体生物量特征. 应用与环境生物学报, 2011, 17(5): 750- 752.
[26] 玉宝, 张秋良, 王立明. 兴安落叶松过伐林枯立木分布格局特征分析. 林业科学研究, 2015, 28(1): 81- 87.
[27] Carmona M R, Armesto J J, Aravena J C, Pérez C A. Coarse woody debris biomass in successional and primary temperate forests in Chiloé Island, Chile. Forest Ecology and Management, 2002, 164(1/3): 265- 275.
[28] Eaton J M, Lawrence D. Woody debris stocks and fluxes during succession in a dry tropical forest. Forest Ecology and Management, 2006, 232(1/3): 46- 55.
[29] KaPeluck P R. Coarse woody debris in an age sequence of southern Appalachian forests [D]. Clemson: Clemson University, 2002.
[30] 王春权, 孟宪民, 张晓光, 夏丹丹. 陆地生态系统碳收支/碳平衡研究进展. 资源开发与市场, 2009, 25(2): 165- 171.
[31] 刘茂秀, 史军辉, 王新英, 彭秋梅. 林分郁闭度和龄级对防护林碳汇功能及固碳价值的影响. 东北林业大学学报, 2011, 39(6): 30- 32.
[32] 魏亚伟, 周旺明, 于大炮, 周莉, 方向民, 赵伟, 包也, 孟莹莹, 代力民. 我国东北天然林保护工程区森林植被的碳储量. 生态学报, 2014, 34(20): 5696- 5705.
[33] Li Y, Wang Y G, Houghton R A, Tang L S. Hidden carbon sink beneath desert. Geophysical Research Letters, 2015, 42(14): 5880- 5887.
[34] Moseley K R, Castleberry S B, Mark Ford W. Coarse woody debris and pine litter manipulation effects on movement and microhabitat use ofAmbystomatalpoideumin aPinustaedastand. Forest Ecology and Management, 2004, 191(1/3): 387- 396.
[35] 杨丽雯, 何秉宇, 黄培祐, 努尔巴依. 和田河流域天然胡杨林的生态服务价值评估. 生态学报, 2006, 26(3): 681- 689.
[36] 王维奇, 徐玲琳, 曾从盛, 仝川, 张林海. 河口湿地植物活体-枯落物-土壤的碳氮磷生态化学计量特征. 生态学报, 2011, 31(23): 7119- 7124.
[37] 程滨, 赵永军, 张文广, 安树青. 生态化学计量学研究进展. 生态学报, 2010, 30(6): 1628- 1637.
Biomass and stoichiometry of dead branches of standing trees inPopuluseuphraticaforests with different ages
SHI Junhui*, LIU Maoxiu, WANG Xinying, MA Xuexi
ResearchInstituteofAfforestationandSandControl,XinjiangAcademyofForestryScience,Urumqi830046,China
The temporal and spatial variations of stoichiometry and carbon (C) storage of the dead branches were revealed in the natural forest trees after having investigated their biomass. The results indicated that: (1) the biomass of the dead branches from a single tree, increased with the forest age and the average annual cumulative biomass of the dead branches, was 0.22 kg per tree; The biomass of the dead branches increased with the enhancing diameters, and the larger diameter grades would induce the easier dead branches. Biomass of the dead branches originally increased and then decreased with increasing forest age, and the highest biomass value was 10.93 t/hm2contributing 17.79% of the total ground living biomass in the mature forests; (2) the C, N, P, K contents of the dead branches were 491.01, 4.13, 2.75 g/kg and 1.83 g/kg, respectively, and were higher than those on the living branches, whereas the K content showed the opposite trend. The ratios of C∶N, C∶P, and N∶P in dead branches of different forests were 128.58, 232.79, and 1.95, respectively, and were higher than those in the living branches. Forest age had little effect on element contents and stoichiometry, which could indicate that element content of dead branches of the standing trees varied slightly; and (3) the C density of the dead branches first increased and then decreased with forest age, and was highest in the mature forests. The average carbon density of different aged forests was 2.63 t/hm2, which was 20.74% of that of the living forests. It was considered the variation law that the biomass and carbon density of the dead branches increased with forest age, resulting in a potentially huge carbon sink. This is a fundamental factor to consider for environmental and forestry management.
Populuseuphratica; dead branches of standing trees; biomass; stoichiometry
中央财政林业科技推广项目(xjlk[2013]010)
2015- 09- 17;
日期:2016- 06- 14
10.5846/stxb201509171916
*通讯作者Corresponding author.E-mail: junhui_shi@sohu.com
史军辉,刘茂秀,王新英,马学喜.不同林龄胡杨活立木枯枝生物量和化学计量特征.生态学报,2017,37(3):887- 895.
Shi J H, Liu M X, Wang X Y, Ma X X.Biomass and stoichiometry of dead branches of standing trees inPopuluseuphraticaforests with different ages.Acta Ecologica Sinica,2017,37(3):887- 895.
猜你喜欢
中国肿瘤外科杂志(2023年5期)2023-11-04 05:50:14
江西农业大学学报(2022年1期)2022-02-04 11:51:42
土壤学报(2021年4期)2021-09-10 03:45:40
江西农业大学学报(2021年2期)2021-05-16 02:18:24
农业机械学报(2020年2期)2020-03-09 08:03:32
森林工程(2019年3期)2019-06-07 07:22:54
森林工程(2019年3期)2019-06-07 07:22:54
防护林科技(2016年5期)2016-09-05 01:19:50
林业与生态(2016年2期)2016-02-27 14:23:42
西藏大学学报(自然科学版)(2015年1期)2015-12-25 11:43:44
