
黑潮营养盐输入对东海陆架浮游生态系统影响的模型研究*
2017-03-31 06:17:45王玉衡沈家葳
海洋与湖沼 2017年6期
王玉衡 赵 亮 沈家葳
(1. 天津科技大学海洋与环境学院 天津 300457; 2. 天津市海洋资源与化学重点实验室 天津 300457)
东海是西北太平洋典型的边缘海, 也是我国重要的渔业产地(宁修仁等, 1995)。黑潮作为东海重要的外源营养盐来源一直受到研究者的关注。对黑潮输入东海陆架营养盐总量的估算, 现有的研究大部分是基于水量平衡的箱式模型, Chen等(1999)最早给出黑潮上陆架营养盐通量的估算, 并指出黑潮和台湾暖流是东海陆架主要的外源营养盐来源。这一结论得到了后续研究的支持(Fang, 2004; Zhanget al, 2007)。此外也有部分研究采用实测营养盐与文献中的流量数据进行估算, 如董书航等(2016)给出了春季黑潮上陆架的营养盐通量; 或者采用数值模式的模拟结果进行计算, 如 Zhao等(2011)基于耦合生态模型对黑潮沿200m等深线跨陆架通量的模拟。虽然不同研究的计算结果有差异, 但现有研究普遍显示黑潮至少贡献了东海陆架一半的外源营养盐输入。
除了对总量的估算, 富含营养盐的黑潮次表层水入侵陆架的路径是另一个研究的焦点, Kondo(1985)认为台湾东北上陆架的黑潮分支可以越过100m等深线抵达 30°N左右的长江浅滩外缘, 这一结论得到了Yang等(2011)数值模拟结果的支持。Su等(1987, 1994)通过对水文数据和锚系站流速数据的分析, 认为黑潮次表层水入侵后, 可以北上至 28°—29°N, 但主要沿 100—200m 等深线移动, 基本不会进入中陆架和内陆架。Qiu等(1990)利用数值模型对东海黑潮进行了模拟, 结果显示台湾东北入侵的黑潮次表层水北上至 27°N附近就返回了黑潮主干, 同样基本不会越过 100m等深线。Ichikawa等(2002)认为黑潮进入东海陆架的分支可以越过100m等深线, 但会在28°N附近回到黑潮主干或者汇入九州以西的对马暖流源地。黑潮次表层水入侵陆架的路径目前仍然存在一定争议。
箱式模型解决了黑潮对东海陆架营养盐输入总量的问题, 但是无法给出输入的营养盐在陆架上是如何分布的, 黑潮次表层水入侵路径的研究在一定程度上有助于了解黑潮输入营养盐的分布范围, 但无法很好定量黑潮输入营养盐在各海区的分配。本文通过数值模式的敏感性实验, 对黑潮在不同季节输入到不同海区和水层的营养盐进行了定量化的对比。浮游植物的情况则更为复杂, 除营养盐以外, 温度、光照以及海水稳定性等都会对浮游植物的生长产生影响(Sverdrup, 1953; Eppley, 1972; Mitchellet al,1991)。不同季节, 输入到不同海区和水层的营养盐对浮游植物生长的贡献是不一样的, 输入限制性营养盐和非限制性营养盐的效果也有差别。黑潮对东海陆架不同海区浮游植物或者初级生产力的贡献是现有研究较为缺乏的, 也是我们试图回答的另一个问题。
1 模型配置与验证
本文采用 Zhao等(2011)建立的生态动力学耦合模型进行敏感性实验, 包括水动力和生态动力学两个模块, 以在线耦合的方式运行。水动力部分模块——普林斯顿大学海洋模型 (Princeton Ocean Model,POM, Blumberget al, 1987), 包含3层嵌套, 内层水平分辨率1/18°×1/18°, 垂向采用σ坐标, 分为21层。生态动力学模块(Aksneset al, 1995; Skogenet al,1995; Zhaoet al, 2011)包括氮(DIN)、磷(DIP)、硅(DSi)三种营养盐, 硅藻和甲藻两种浮游植物类群, 以及有机碎屑和生源硅两种有机颗粒物。模型运行3年以达到稳定状态, 取第4年的结果进行分析, 分析区域如图1所示。

图1 研究区域与地形Fig.1 Study area and the topography
Zhao等(2011)就模式PN断面的模拟结果与日本气象厅的调查数据进行了对比验证, 发现模式模拟的叶绿素和 DIN断面分布都很好的抓住了观测中的主要特征。为了分析黑潮对东海陆架的影响, 本文选取 Gong等(2003)在东海陆架的表层叶绿素观测数据对模型进行进一步的验证(图 2)。观测数据显示了冬季到初春东海陆架表层叶绿素浓度普遍较低, 夏季在长江口和杭州湾以东出现高值区, 秋季整体回落的规律, 在模式结果中都得到了较好的模拟。虽然个别高值中心位置的模拟略有偏差, 但考虑到 Gong等(2003)的观测结果是1997—1998年间4个航次的数据,而模型强迫是多年平均的结果, 这一差异是可以理解的。这充分说明了模型在对黑潮模拟方面的可靠性。

图2 表层叶绿素分布的观测与模拟结果Fig.2 Simulation and observation on distribution of the chlorophyll in surface layer
为了研究黑潮对东海陆架生态系统的影响, 设定一组敏感性实验改变开边界黑潮输入营养盐浓度,并通过与受控实验的结果进行对比, 来定量化分析黑潮营养盐输入对东海陆架不同区域不同层次浮游生态系统的贡献。考虑到浮游生态系统是个复杂的非线性系统, 为了保证敏感性实验的合理性, 敏感性实验的条件设置参考了Guo等(2012)黑潮营养盐通量年际变化的研究结果。Guo等(2012)通过分析 1987—2009年间的现场观测数据计算了PN断面黑潮硝氮通量的年际变化, 得到的多年平均硝氮通量为 170.8kmol/s,年际变化的趋势是每年增加 0.88kmol/s。据此估算,过去20年黑潮的硝氮通量大约增加了10%。该研究同时指出硝氮和 DIP浓度具有良好的线性关系(R=0.99), 可以认为 DIP的通量变化规律与硝氮类似。DSi通量的年际变化缺乏足够的观测数据。但Kodama等(2014)利用黑潮134°E断面2002—2013年现场观测数据的分析显示, 在中上层DSi与硝氮和亚硝氮浓度之和也存在较好的线性关系。考虑到除表层水外, 黑潮 DIN主要以硝氮的形式存在(在次表层约占88%, 中层约占98%, Zhanget al, 2007), 在敏感性实验的配置中, DIP和DSi通量都采用了与DIN相同的设定, 即对黑潮区开边界的DIN、DIP和DSi浓度改变 10%, 来模拟黑潮输入营养盐年际变化对东中国海浮游生态系统的影响。
2 结果
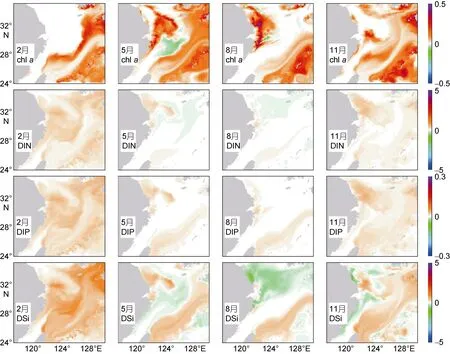
图3 黑潮营养盐增加对东海表层叶绿素及营养盐贡献的分布Fig.3 Contribution of nutrient increase in Kuroshio to the surface chlorophyll and nutrients in the East China Sea填色代表敏感性实验与受控实验结果之差, 从上至下依次为叶绿素(chl a, mg/m3)、溶解态无机氮(DIN, μmol/L)、溶解态无机氮(DIP,μmol/L)和溶解态硅酸盐(DSi, μmol/L)
图 3显示了黑潮营养盐增加 10%后给东海表层叶绿素及营养盐带来的增量, 黑潮的营养盐输入以高磷著称(Zhanget al, 2007), 反映在实验结果中, DIP各个季节都表现为增加, 但秋冬季增加的区域连成一片, 春夏季则分割为黑潮区和长江口-苏北两个孤立的区域。
DIN和DSi分布有一定的相似性。冬季都是普遍增加; 春季黑潮区和苏北浅滩增加, 东海内陆架和中陆架减少; 夏季黑潮区增加, 长江冲淡水范围内减少;秋季DIN是整体增加的, 但DSi在浙闽沿岸、苏北和苏岩礁附近是减少的。
叶绿素相对复杂。冬季是黑潮主轴两侧增加明显;春季黑潮区和长江口以东增加, 东海陆架减少; 夏季跟春季类似, 但没有了陆架上减少的区域; 秋季则与冬季类似, 但在杭州湾以东多出一个孤立的增加区域。
从大面分布中可以明显的看到, 黑潮对东海表层营养盐和叶绿素的影响非常广泛, 几乎遍布整个陆架, 但是在不同区域体现出不同的特征。单纯从表层浮游植物的增量来看, 呈现出内陆架和外陆架增量较高, 中陆架则基本不受影响的特点, 尤以春夏两季最为明显。从营养盐的分布, 也可以看到高营养盐的黑潮水对陆架的入侵, 从南到北并不均匀。为详细了解黑潮对东海陆架不同区域生态系统的影响, 本文选择了横跨陆架的 PN断面(图 4)以及沿50m等深线的ES1断面(图5)进行分析, 断面位置见图1。

图4 黑潮营养盐增加对PN断面叶绿素及营养盐贡献的分布Fig. 4 Contribution of nutrient increase in Kuroshio to the chlorophyll and nutrients in the PN section
PN断面3种营养盐变化量的分布规律较为相似,冬季垂向混合均匀, 从外陆架到内陆架浓度逐渐降低, 底层增量大于表层; 春夏两季虽然表层只能在内陆架看到营养盐增加的海区, 但是中层和底层的黑潮水入侵仍然清晰可见; 秋季随着垂向混合的增强,黑潮水的影响逐渐从中底层向表层扩展。
叶绿素的变化, 冬季主要在外陆架, 表层最为明显, 但在 50—100m 的水深上, 也仍然可以看到叶绿素浓度的增加; 春季主要在表层, 内陆架和外陆架升高, 中陆架则有所降低, 这可能与早春水华对表层营养盐的消耗有关; 夏季内陆架表现在表层, 中陆架和外陆架主要体现在次表层, 这与夏季真光层加深有关; 秋季则在整个表层海区普遍出现上升的态势, 随着跃层的减弱, 中底层营养盐向上补充有利于表层叶绿素的增加。值得注意的是除了冬季以外, 其他季节叶绿素的高值区往往对应着营养盐的低值, 这种反向分布在一定程度上反映了东海陆架浮游生态系统在夏半年有下行控制的特点。冬季的叶绿素局限于外陆架体现了光限制, 夏秋季中陆架表层的降低则主要是营养盐限制的结果。

图5 黑潮营养盐增加对ES1断面叶绿素及营养盐贡献的分布Fig.5 Contribution of nutrient increase in Kuroshio to the chlorophyll and nutrients in the ES1 section注: 图中以倒三角标注与S断面和PN断面的交点
对ES1断面, 根据其与PN、S两断面的交点, 分成南中北3段。黑潮水对ES1断面的营养盐输入明显集中于中段, 而且自南向北影响从底层逐渐向表层扩展; 南段接收的营养盐较少, 叶绿素也几乎没有变化; 北段从营养盐浓度来看, 变化不如中段明显, 甚至夏季还出现了DIN和DSi的降低, 叶绿素的增加却是最为显著的。考虑到ES1断面的北段位于长江浅滩的外缘, 属于初级生产非常旺盛的海区, 比较合理的解释是北段浮游植物的生长消耗了大量营养盐, 从而呈现出叶绿素增加而营养盐几乎不变的分布特征,同时长江口外区域以磷限制为主(林军等, 2011; 王奎等, 2013), 黑潮大量补充 DIP, 在刺激叶绿素增加的同时, 也造成了该区域DIN和DSi的减少。
两个断面的叶绿素和营养盐分布除了反映黑潮贡献在沿等深线和垂直等深线两个方向的差异, 也反映了不同水层受黑潮影响的不同。比如PN断面中段夏秋季的次表层叶绿素峰值和 ES1断面中段营养盐自南向北逐渐向表层入侵等特征, 都需要通过对叶绿素和营养盐的变化进行分层统计来加以体现。因此, 本文分别统计了敏感性实验中不同海区不同水层中叶绿素和营养盐变化的积分值, 以便定量化分析黑潮对东海陆架浮游生态系统的贡献。
3 讨论
从图4和图5的断面分布可以看出, 黑潮对东海陆架生态系统的贡献既有沿黑潮主轴方向的变化,又有从外陆架到内陆架的差异。因此, 以PN和S两个断面将东海陆架分为南、中、北三部分, 以ES1(50m等深线)和 ES2(100m等深线)两个断面划分内陆架、中陆架和外陆架。这样, 整个东海陆架被分成9个海区, 具体海区划分如表1所示。
对不同区域的叶绿素和营养盐变化进行积分,年平均以后的统计结果见表1。三种营养盐的分布较为相似, 都呈现出自外陆架向内陆架递减, 从南向北递增的趋势。具体到9个区域, 内陆架南部几乎没有黑潮营养盐的输入, 内陆架的中北部和中陆架的南部得到的营养盐输入也较少, 营养盐的输入主要还是集中在中陆架的中北部和外陆架。这一分布趋势与Ichikawa等(2002)根据盐度调查给出的台湾以北黑潮分支的路径较为相似, Yang等(2011)利用ROMS模型进行的质点追踪实验也得到了类似的结果, 即台湾东北部黑潮次表层水上陆架后, 大致沿122°E向北扩散, 在26°N附近越过100m等深线, 在29°N附近达到60m等深线, 之后沿60m等深线抵达长江口和杭州湾以东海区, 其流经海区与上述敏感性实验中营养盐增加较为明显的海区基本是一致的。

表1 年平均的区域积分叶绿素及营养盐变化Tab.1 The annual mean of region-integrated increment of chlorophyll and nutrients
叶绿素的分布则与营养盐存在一定差异, 表现在内陆架北部和外陆架中部两个海区的叶绿素增量在整个东海陆架中的占比明显高于营养盐的比例(两个海区的叶绿素增量分别占整个陆架的 7.4%和27.0%, 营养盐以DIP为例则只占3.8%和18%); 与之对应的, 外陆架南部则是营养盐输入较多但叶绿素增量偏低的典型(叶绿素和 DIP占比分别为 7.1%和12.1%)。为了分析这一差距的来源, 本文对敏感性实验的结果分冬季(图6)和夏季(图7)进行了分层积分。
3种营养盐的垂向分布和季节变化表现有一致特征。就垂向分布而言, 整体上以50m以深的中下层占据优势, 夏季尤为明显, 以DIP为例, 2月和8月中下层的增量分别占到整个海区的 48%和 68%; 分海区来看, 外陆架的 3个海区中下层的优势都非常显著,而中陆架 20—50m的次表层和中下层所占的比例较为接近, 内陆架营养盐增量比较小, 由于水深的原因,没有 50m以深的水层, 营养盐集中在表层和次表层,冬季表层占优, 夏季二者相当。就季节变化而言, 营养盐输入的总量和区域分布格局, 2月和8月的统计结果都没有明显差异, 仍以DIP为例, 2月整个东海陆架的增量为294×109mol, 8月份略低为261×109mol,但在层次分配上, 有一定的区别, 8月营养盐输入更加集中于底层。
叶绿素的分布则与营养盐体现出截然不同的特征。在垂向上, 绝大部分海区的叶绿素增量集中在表层和次表层。在季节变化上, 虽然叶绿素增加的总量变化不大(2月为282×106g, 8月为239×106g), 但是在区域和层次上存在明显的差异。2月的叶绿素增量主要集中在外陆架的中北部和中陆架的中部, 表层和次表层几乎各半。而8月份则主要在外陆架的中部、中陆架的中北部和内陆架的北部, 除了内陆架北部以外都以次表层为主。从前面的大面和断面分布可以看到夏季的叶绿素和营养盐高值是反向分布的, 所以如果一个海区的叶绿素增量集中在夏季, 对应的营养盐增量会显得较低。从图6和图7可以看出内陆架北部和外陆架中部夏季叶绿素增量明显高于冬季,这在一定程度上解释了为什么这两个海区的叶绿素增量占比明显高于营养盐。

图7 8月份各海区分层叶绿素(a)及营养盐(b: DIN; c: DIP;d: DSi)变化Fig.7 Integrated increment of chlorophyll (a) and nutrients(b:DIN; c: DIP; d: DSi) in August.
目前利用观测对陆架区营养盐收支的估算主要有Chen等(1999)和Zhang等(2007)对三种营养盐在陆架区整体收支分季节的估算, 以及Fang(2004)对磷酸盐的估算, 其中 Chen等(1999)的结果还提供了黑潮分层的收支。此外 Zhao等(2011)采用与本文相同的三维水动力模型, 估算了黑潮对东海陆架的营养盐输入。
整体来讲, 对黑潮输入的营养盐, 多数文献的估算结果差别不大, 仍以 DIP为例, Chen等(1999),Zhang等(2007), Zhao 等(2011)和Fang(2004)四组研究估算的年均通量分别为0.34、0.652、0.7和0.82kmol/s,只有Chen的结果较其他研究偏低。对比Chen等(1999)和 Zhang等(2007)冬夏两季的收支, 可以看到冬季Chen等(1999)的估算值(0.37kmol/s)明显低于 Zhang等(2007)的结果(0.92kmol/s), 而夏季则差别不大(二者分别为 0.32和 0.384kmol/s)。这主要是由于 Chen等(1999)对黑潮的输入采用分层估算, 而 Zhang等(2007)采用整层估算, 二者选用的黑潮入侵陆架的水量数据冬季都高于夏季, 但 Chen等(1999)的数据中冬夏季黑潮差别主要在表层水入侵, 次表层和中层变化不大, 而表层水营养盐浓度相对较低, 所以冬季黑潮入侵流量虽大, 营养盐输入通量的变化却没有那么明显。
对黑潮的估算, Chen等(1999)的方法显然更加合理, 本文敏感性实验中的模拟结果显示黑潮入侵带来的营养盐并不存在明显的季节变化, 以 DIP为例,黑潮输入营养盐浓度增加 10%的情况下, 陆架区增加的 DIP在 8月份为 261×109mol, 2月份为 294×109mol, 这一结果与 Chen等(1999)的估算中黑潮输入营养盐的季节变化规律是一致的。但是根据 Zhao等(2011)的估算, 模式中黑潮输入陆架的营养盐总量却与Zhang等(2007)的结果更接近。这一差别显示了目前对黑潮营养盐贡献的估算仍然有待改进, 黑潮对陆架的入侵并不是单向的从黑潮主干进入东海陆架, 而是在不同的区域, 既有黑潮水入侵陆架, 又有入侵后的黑潮水返回黑潮主干, 根据 Zhao等(2011)对模型结果的计算, 黑潮在台湾东北大约有5.8Sv入侵陆架, 在九州西南有 0.5Sv入侵, 而其他海区则有约4.7Sv的入侵水返回了黑潮主干。在以往箱式模型的估算中(如Chenet al, 1999; Zhanget al, 2007), 黑潮的营养盐通量是用黑潮水入侵陆架的净通量乘以黑潮营养盐浓度来进行估算的, 这其中隐含的假设是黑潮水入侵陆架时和从陆架返回黑潮主干时, 营养盐浓度是相等的, 这一假设显然有待商榷。Zhao等(2011)的计算方法则是用黑潮入侵陆架的水量乘以黑潮营养盐浓度再减去返回黑潮主干的水量乘以陆架水的营养盐浓度, 考虑到黑潮水整体营养盐浓度是高于陆架水的, 这一算法得到的黑潮营养盐通量要高于箱式模型。
4 结论
本文利用三维物理-生化耦合模型, 通过设计敏感性实验, 分析了黑潮营养盐输入增加后, 东海营养盐和叶绿素的响应。并分区域、季节和水层进行定量统计, 得到的主要结论如下:
黑潮输入营养盐增加 10%的情况下, 东海陆架区年平均的营养盐增量呈现出自外陆架向内陆架递减, 从南向北递增的趋势。其对区域积分叶绿素的贡献, 以外陆架中部(63.68×106g)、外陆架北部(52.90×106g)和中陆架中部(41.14×106g)最为显著。
冬季, 营养盐的输入主要集中在中陆架和外陆架, 外陆架整体上以50m以深的中下层占据优势, 而中陆架 20—50m的次表层和中下层所占的比例较为接近。叶绿素增量主要分布在外陆架的中北部和中陆架的中部, 表层(0—20m)和次表层几乎各半。
夏季, 营养盐增量的分布整体与冬季相似, 但在垂向上, 中下层的优势更明显。叶绿素则主要在外陆架的中部、中陆架的中北部和内陆架的北部, 除了内陆架北部以外都以次表层为主。
王 奎, 陈建芳, 金海燕等, 2013. 长江口及邻近海区营养盐结构与限制. 海洋学报, 35(3): 128—136
宁修仁, 刘子琳, 史君贤, 1995. 渤、黄、东海初级生产力和潜在渔业生产量的评估. 海洋学报, 17(3): 72—84
林 军, 朱建荣, 张 经等, 2011. 长江口外海区浮游植物生物量分布及其与环境因子的关系. 水产学报, 35(1): 74—87
董书航, 刘素美, 任景玲等, 2016. 春季东海营养盐跨陆架输运通量的初步估算. 海洋环境科学, 35(3): 385—391
Aksnes D L, Ulvestad K B, Baliño B Met al, 1995. Ecological modelling in coastal waters: Towards predictive physical-chemical-biological simulation models. Ophelia,41(1): 5—36
Blumberg A F, Mellor G L, 1987. A description of a three dimensional coastal ocean circulation model. In: Heaps N S ed. Three-Dimensional Coastal Ocean Models. Washington,DC, USA: American Geophysical Union, 1—16
Chen C T A, Wang S L, 1999. Carbon, alkalinity and nutrient budgets on the East China Sea continental shelf. Journal of Geophysical Research: Oceans, 104(C9): 20675—20686
Eppley R W, 1972. Temperature and phytoplankton growth in the sea. Fishery Bulletin, 70(4): 1063—1085
Fang T H, 2004. Phosphorus speciation and budget of the East China Sea. Continental Shelf Research, 24(12): 1285—1299
Gong G C, Wen Y H, Wang B Wet al, 2003. Seasonal variation of chlorophyll a concentration, primary production and environmental conditions in the subtropical East China Sea.Deep Sea Research Part II: Topical Studies in Oceanography,50(6—7): 1219—1236
Guo X Y, Zhu X H, Wu Q Set al, 2012. The Kuroshio nutrient stream and its temporal variation in the East China Sea.Journal of Geophysical Research: Oceans, 117(C1): C01026 Ichikawa H, Beardsley R C, 2002. The current system in the Yellow and East China Seas. Journal of Oceanography, 58(1):77—92
Kodama T, Shimizu Y, Ichikawa Tet al, 2014. Seasonal and spatial contrast in the surface layer nutrient content around the Kuroshio along 138E, observed between 2002 and 2013.Journal of Oceanography, 70(6): 489—503
Kondo M, 1985. Oceanographic investigations of fishing grounds in the East China Sea and the Yellow Sea, I: characteristics of the mean temperature and salinity distributions measured at 50 m and near the bottom. Bulletin of the Seikai Regional Fisheries Research Laboratory, 62: 19—66
Mitchell B G, Brody E A, Holm-Hansen Oet al, 1991. Light limitation of phytoplankton biomass and macronutrient utilization in the Southern Ocean. Limnology and Oceanography, 36(8): 1662—1677
Qiu B, Imasato N, 1990. A numerical study on the formation of the Kuroshio counter current and the kuroshio branch current in the East China Sea. Continental Shelf Research,10(2): 165—184
Skogen M D, Svendsen E, Berntsen Jet al, 1995. Modelling the primary production in the North Sea using a coupled three-dimensional physical-chemical-biological ocean model.Estuarine, Coastal and Shelf Science, 41(5): 545—565
Su J L, Pan Y Q, 1987. On the shelf circulation north of taiwan.Acta Oceanologica Sinica, 6(S1): 1—20
Su J L, Pan Y Q, Liang X S, 1994. Kuroshio intrusion and Taiwan Warm Current. In: Zhou D, Liang Y B, Zeng C K eds.Oceanology of China Seas. Dordrecht, Holland: Kluwer Academic Publishers, 59—70
Sverdrup H U, 1953. On conditions for the vernal blooming of phytoplankton. ICES Journal Marine Science, 18(3): 287—295 Yang D Z, Yin B S, Liu Z Let al, 2011. Numerical study of the ocean circulation on the East China Sea shelf and a Kuroshio bottom branch northeast of Taiwan in summer. Journal of Geophysical Research: Oceans, 116(C5): C05015
Zhang J, Liu S M, Ren J Let al, 2007. Nutrient gradients from the eutrophic Changjiang (Yangtze River) Estuary to the oligotrophic Kuroshio waters and re-evaluation of budgets for the East China Sea shelf. Progress in Oceanography,74(4): 449—478
Zhao L, Guo X, 2011. Influence of cross-shelf water transport on nutrients and phytoplankton in the East China Sea: A model study. Ocean Science, 7(1): 27—43
猜你喜欢
沉积学报(2023年1期)2023-04-29 18:57:49
海洋通报(2022年6期)2023-01-07 02:48:56
海洋通报(2022年2期)2022-06-30 06:07:04
海洋信息技术与应用(2021年3期)2022-01-17 02:37:26
建筑热能通风空调(2018年5期)2018-07-09 03:16:44
海洋世界(2017年1期)2017-02-13 08:31:44
中国科技博览(2016年25期)2016-12-20 18:28:41
海洋科学(2016年1期)2016-07-06 09:42:34
海洋渔业(2016年6期)2016-04-16 03:00:23
中国工程咨询(2016年9期)2016-02-13 03:12:40
