
甘蔗农业生产系统模拟模型模块化设计与应用研究进展
2017-03-29 12:12:23InmanBamber刘家勇Jackson范源洪
中国糖料 2017年1期
毛 钧,N.G.Inman-Bamber,杨 昆,陆 鑫,刘家勇,P.A.Jackson,范源洪*
(1.云南省农业科学院甘蔗研究所,开远661699;2.云南省甘蔗遗传改良重点实验室,开远661699;3.詹姆斯库克大学科技与工程学院,汤斯维尔;4.澳大利亚联邦科学与工业研究组织热带农业科学与创新研究中心,汤斯维尔)
甘蔗农业生产系统模拟模型模块化设计与应用研究进展
毛 钧1,2,N.G.Inman-Bamber3,杨 昆1,2,陆 鑫1,2,刘家勇1,2,P.A.Jackson4,范源洪1,2*
(1.云南省农业科学院甘蔗研究所,开远661699;2.云南省甘蔗遗传改良重点实验室,开远661699;3.詹姆斯库克大学科技与工程学院,汤斯维尔;4.澳大利亚联邦科学与工业研究组织热带农业科学与创新研究中心,汤斯维尔)
以APSIM甘蔗模型(APSIM-Sugar)为例,简要介绍了由澳大利亚农业生产系统研究协作组(APSRU)开发的农业生产系统模拟模型(Agricultural Production System siMulator,APSIM)中作物、土壤等核心模块的基本过程原理、参数确定、模型验证等模块化设计与应用研究进展,为学习了解和使用APSIM模型开展农田土壤-作物系统模拟研究提供参考。
甘蔗;农业生产系统模拟模型;生理过程;模块;参数
农业生产系统模拟模型(Agricultural Production System siMulator,APSIM)是由澳大利亚农业生产系统研究协作组(APSRU)自1991年开始研发的一套农田土壤-作物系统模拟软件框架和一系列作物生长模型的总称,目前已经开发到APSIM v7.8版本。作为一个基于农田作物生长发育机理过程模拟的模型系统,APSIM创新地采用了模块化的“插拔(plug-in/plug-out)”系统设计,用户可以根据研究需要将各种相关模块进行自由组合集成,从而对不同类型的农田生产系统进行有效模拟。APSIM模型的特色是采用了基于生物和物理过程的模块化设计理念,其核心构成包括:模拟作物品种生长发育过程的作物模块;模拟土壤水氮动态的土壤模块;模拟农田耕作和种植收获过程的管理模块;模拟环境光、温、水、气变化的气候模块;生物量、产量等模拟结果输出模块;驱动模拟过程和控制不同模块协同运作的中心引擎。其中作物模块和土壤模块是最为关键的核心模块,气候模块和管理模块是主要的数据输入模块。APSIM模型中模拟不同作物的生长发育及其对气候因子和土壤水氮变化的响应都是通过中心引擎控制各模块协同运作来实现的,作物模块中所呈现的各种作物生理过程变量和胁迫因子,能够对气候模块中的太阳辐射、温度、降水等气候变化以及管理模块中的耕作措施做出响应,特别是对土壤模块中的水分和氮素供应较为敏感[1-2]。
甘蔗模型(APSIM-Sugar)是APSIM系列模型中开发较早、较为完善、应用较广的代表性作物模型之一。在APSIM通用模块(土壤、管理、气候等)的基本构架上,插入甘蔗作物模块(Sugar module)并通过甘蔗品种配置文件(sugar.xml)定义甘蔗品种特定的遗传参数,就完成了APSIM-Sugar甘蔗模型的组装。APSIM-Sugar甘蔗模型基于日值气象数据,可用于模拟土壤条件均一的蔗田生产,预测甘蔗地上部生物量、蔗茎、糖分产量、含糖率、水分利用、氮素摄取和分配等[3-4]。本文以APSIM-Sugar甘蔗模型为例,重点介绍APSIM模型中作物生长发育、产量形成、土壤水氮动态模拟相关的过程原理、模块参数设置与应用研究进展,旨在为学习了解和使用APSIM模型开展农业生产系统模拟研究提供参考。
1 APSIM-Sugar模型模块设计与核心模拟过程
APSIM模型的机理性模拟原理在1986年前后开始逐渐形成,Ritchie在1991年进行了综合[5-8]。AUSCANE是澳大利亚第一个被广泛使用的甘蔗模型,其模拟原理基于EPIC模型,但该模型在甘蔗生物学模拟和程序设计方面存在着不足[9]。此后,APSRU研发小组在APSIM模型中开发了一个甘蔗模块(Sugar module),其基本原理综合了CERES-Maize和AUSCANE等早期模型,并充分利用了APSRU在土壤水和氮素模拟方面的研究成果。APSIM-Sugar模型的开发指导思想是构建一个相对简单的甘蔗生理学过程模型,对每个甘蔗生理过程都采用简约的方法以避免过于复杂化[3,10]。甘蔗模块简化了大量的过于复杂而难以测定的变量参数,但又能确保在各种生长环境条件下,甘蔗生长发育模拟过程必需的描述性参数需求。构建APSIM甘蔗模型所需要的基础数据库是科学选取了澳大利亚、夏威夷、南非、新西兰和斯威士兰等不同纬度跨度国家,经过大量试验调查数据所建立的,涵盖了不同的种植时间、种植品种、土壤水分和氮素供给等真实环境条件,模型参数由这些数据集综合分析得到。由于模型框架结构和模块参数设计合理,并有大田数据支持,APSIM模型和甘蔗模块在包括中国在内的世界各国得到了广泛应用[2-3,10-13]。
APSIM-Sugar模型的核心模拟过程反映了在设定的太阳辐射、气温、土壤水分和氮素供应环境下,甘蔗植株的生长发育、冠层扩展、生物量形成与分配等相关变量对气候因子、土壤水分和氮素的响应和利用情况(表 1)。
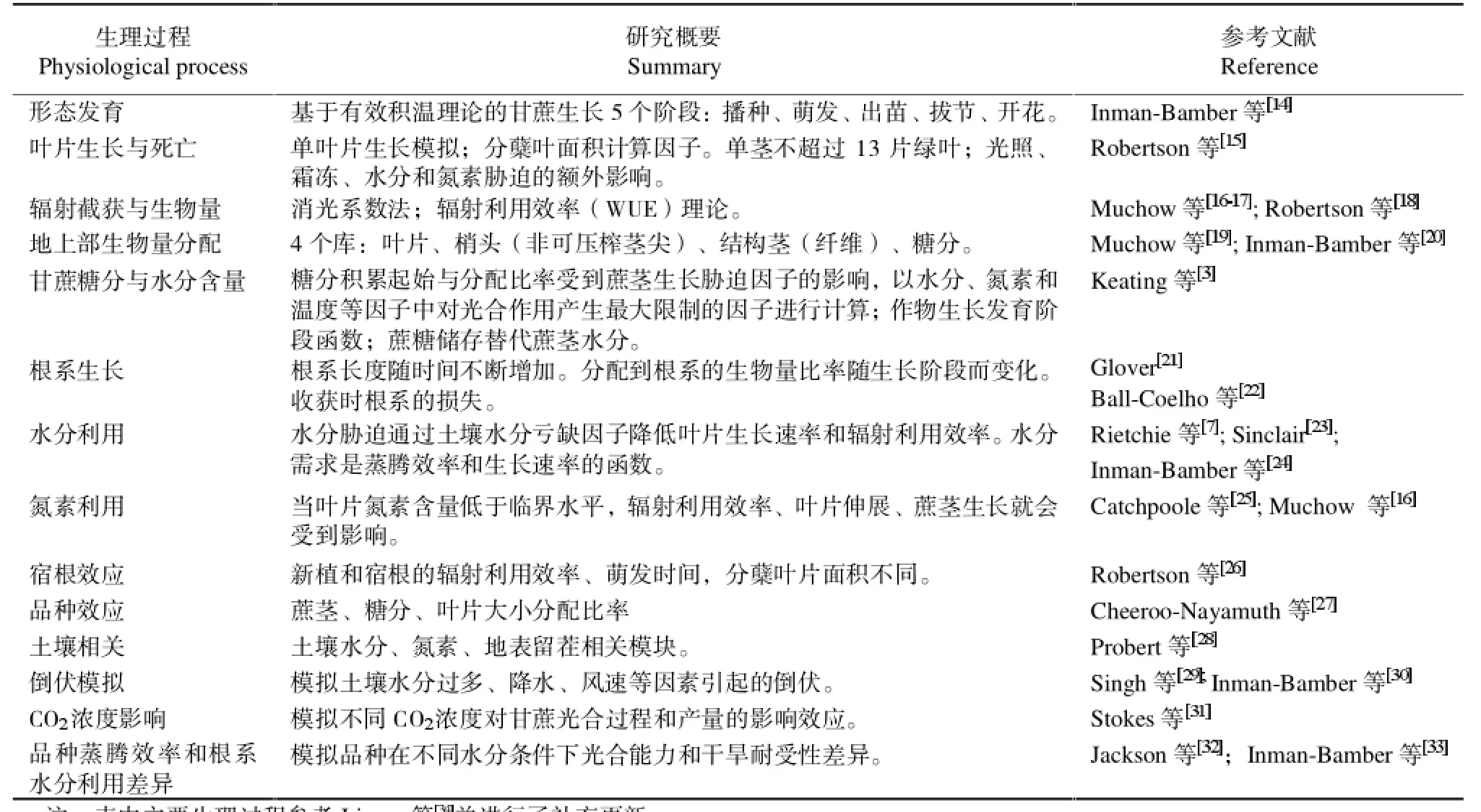
表1 APSIM-Sugar 模型核心模拟过程与相关苷蔗生理学研究概要Table 1 The main physiological process and summary simulated in APSIM-Sugar
2 APSIM-Sugar模型甘蔗品种遗传参数设置
APSIM-Sugar模型甘蔗品种主要遗传参数约有17个,通常是在甘蔗模块品种配置文件(sugar.xml)中进行设定。默认品种Q117各参数取值及说明见表2,其中包括冠层扩展相关的形态学参数(1,2,12,13,14);生育期相关参数 (9,10,11);生物量、糖分积累和分配系数 (3,4);环境胁迫对产量糖分的影响效应系数(5,6,7,8);叶片蒸腾水分利用效率TE(15);太阳辐射利用效率RUE(16);根系提水系数kL(17)等。
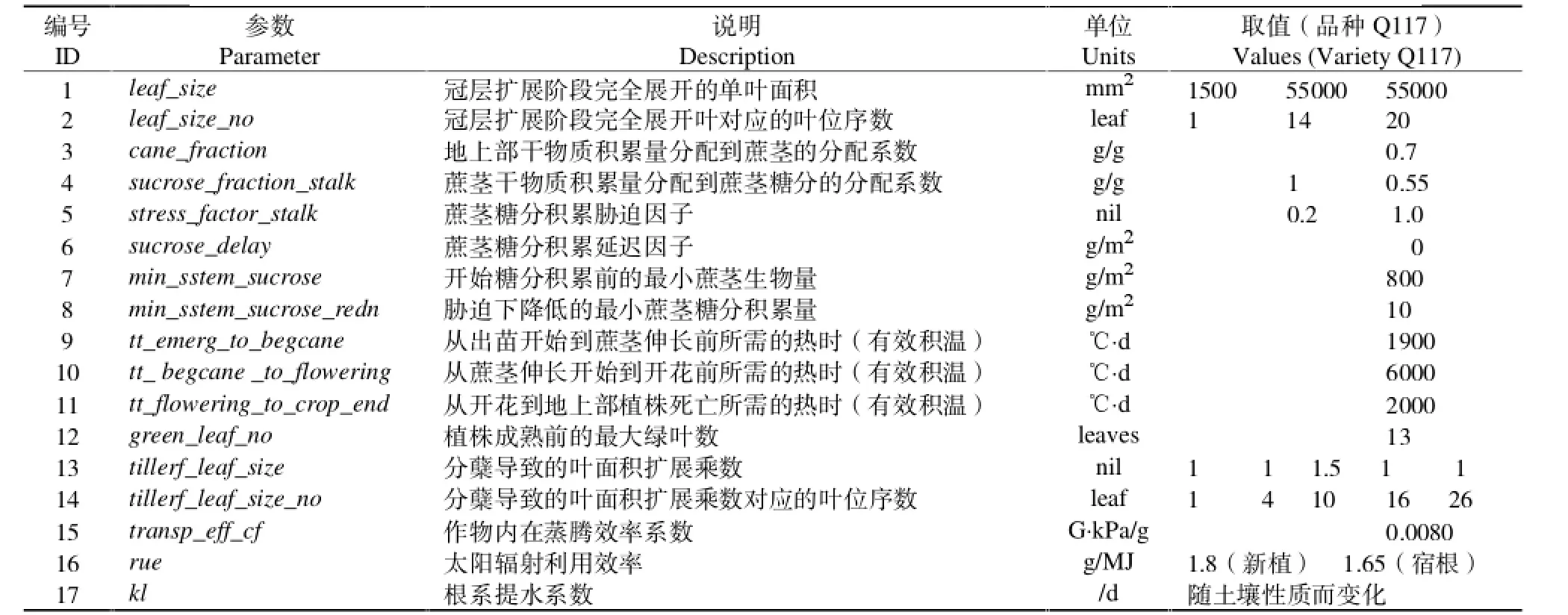
表2 APSIM-Sugar 模型苷蔗品种遗传参数Table 2 Cultivar genetic parameters in APSIM-Sugar
APSIM-Sugar模型中一个完整的甘蔗生育期包括6个主要的物候学事件:播种、萌芽、出苗、蔗茎伸长、开花和地上植株死亡。但在大田实际生产中,甘蔗品种开花只是零星发生,蔗茎通常在开花前就可达到生理成熟并收获。另外对于宿根蔗而言无需再次播种,直接由存活的地下蔗桩发株出苗。因此目前的甘蔗模块中仅模拟了3个物候期,即出苗期(从萌芽到出苗),分蘖期(从出苗到蔗茎伸长前),蔗茎伸长期(从蔗茎伸长开始到收获)。生育期参数也仅对出苗到蔗茎伸长(表2参数9),蔗茎伸长到开花(表2参数10),开花到地上植株死亡(表2参数11)所需的积温进行了粗略的描述,出苗时间需要通过管理模块的播种日期和播种深度等参数进行调整,而与花期相关的物候学参数调整并没有实际效果[3,36]。热时(有效积温)对甘蔗植株的生长发育和冠层扩展影响很大,在APSIM模型中,热时-温度关系是根据冠层三基点温度进行计算的。模块设定甘蔗正常发育的下限温度为9℃,最适温度为32℃,上限温度为45℃。新植蔗由播种到萌芽需要350℃·d的有效积温时滞,而宿根蔗发株只需100℃·d的有效积温时滞。甘蔗萌芽后将以0.8mm/(℃·d)的速率生长直至出土。甘蔗由出苗至分蘖结束进入蔗茎伸长期所需的有效积温因甘蔗品种差异在1200~1900℃范围内变化。通常蔗茎总数在分蘖开始后1400℃·d的有效积温时滞内快速增加至最高数量,然后逐渐下降到相对稳定的蔗茎总数。宿根蔗通常比新植蔗较早达到最高蔗茎总数,因此其早期冠层扩展很快。冠层叶片扩展的模拟是通过用户定义的品种叶面积发育函数和分蘖模式经验校正系数进行计算。如甘蔗单株总叶面积是由其全部绿叶的完全展开总叶面积乘以分蘖校正系数得到。叶片长出率是有效积温的连续函数,模型通过线性插值的方法推算新植蔗和宿根蔗的叶片生长。分蘖初期每生长一片蔗叶需80℃·d的有效积温,进入蔗茎伸长期生长一片蔗叶需150℃·d的有效积温。蔗田甘蔗总叶面积的增减主要由4个因素决定:甘蔗发育、光照竞争、水分胁迫和霜冻。当甘蔗植株的完全展开叶达到13片后进入冠层扩展成熟期,开始启动对叶片衰老和死亡的模拟。当蔗田内甘蔗叶片的太阳辐射截获率超过0.85或土壤水分亏缺因子低于1.0时,由于植株间的光竞争和土壤水分胁迫,可能引起总叶面积的减少和降低有效光合叶面积。当气温低于0℃时产生的霜冻可导致总叶面积减少10%,当气温低于-5℃时,叶片将全部死亡[3,14]。
甘蔗模块对光合同化作用的模拟是基于太阳辐射利用效率(RUE)和水分利用效率(WUE)即蒸腾效率(TE)相关理论进行的[37]。首先计算冠层叶片每天截获的太阳辐射量,然后根据作物的RUE经验系数转化为每日同化的干物质量,并按一定比例分配到不同的组织器官,最后根据物质能量平衡计算总体生物量累积。正常生长情况下,新植蔗和宿根蔗分别以1.8g/(MJ·m2)和1.65 g/(MJ·m2)的RUE经验系数将冠层截获的有效太阳辐射量换算为每天生物量。在极端气温、土壤水分短缺或过量、氮素亏缺限制光合作用等情况下,RUE将会减小。例如平均气温在15~35℃之间,RUE取最大值。平均气温低于5℃或超过50℃,RUE降为零。甘蔗生物量以不同分配系数被分配到5个储存库中:根系、展开叶、茎秆、梢头幼叶和蔗糖分[1]。通常地上部生物量的70%被分配到茎秆中,当甘蔗进入伸长期以后,原来分配到茎秆的生物量会重新分配到蔗茎纤维用于结构生长和蔗茎糖分用于养分存储。如果分到叶片中的生物量不能满足生长需求,则叶片生长速率会降低。如果分到叶片中的生物量超过生长需求,多余的生物量会转移到糖分库和茎秆纤维中。根的生物量增加是独立的,每天的全部生物量以固定比例分配给根系,分配比例随着植株由分蘖至成熟,由最大30%逐渐减少到20%。根系生物量以18mm/g系数换算为根系总长。植株分蘖后,若土壤水分充足,根系将以0.8mm/(℃·d)的速率伸长。蔗茎鲜重是产量构成的重要指标,甘蔗模块设定植株茎秆含水量与茎秆干物重的比例由甘蔗生长初期的9∶1过渡到后期的5∶1[38]。
3 APSIM-Sugar模型土壤水氮模块的参数设置
3.1 土壤水分模块
APSIM-Sugar模型可选用两个模块(SoilWater和SWIM)来模拟土壤水分剖面的动态变化。SoilWater模块的核心是土壤分层阶梯式渗透模型[39],其原理主要来源于CERES模型和PERFECT模型,整个土壤剖面中水分再分配的算法也是继承CERES模型。SWIM模块基本原理基于Richard方程的土壤水动力学模型[5]。例如,SoilWater模块以日为时间单位进行模拟,各个模拟进程都是连续的。土壤剖面每层的水分特征可由萎蔫系数(LL15)、田间持水量(DUL)和饱和含水量(SAT)等土壤参数来表述。LL15表示水吸力15巴时的土壤含水量,是作物根系正常吸水的最低土壤含水量下限;DUL即田间持水量,表示排除重力水后仍然能够保持的土壤含水量;SAT为土壤吸收水分至完全饱和时的含水量。在APSIM模型土壤参数设置中,主要通过输入土壤剖面每层的土壤体积含水量进行土壤水分特征的初始化[40]。土壤剖面水分平衡模型的模拟计算过程包括:各层土壤含水量变化,降雨和灌溉,地表径流、饱和渗漏、非饱和入渗和水分扩散,根系水分吸收等[28]。灌溉量和降水量是作为模型的初始参数输入,灌溉量在管理模块中进行定义,降水量通过气象文件(后缀名为met的文本文件)设定。降水所导致的地表径流量R0则是利用径流曲线数方法(USDA-Curve Number)来估算。SoilWater模块根据前期降水状况,首先计算出湿径流曲线(高潜在径流量)和干径流曲线(低潜在径流量),然后根据每天的土壤含水量变化和两种极端径流曲线估算实际径流量。土壤表层植被密度和地表残留物覆盖也会制约土壤表面产生径流的能力,对此,SoilWater模块中通过设置一个参数(CNcov)来计算土壤表层覆盖的影响。土壤饱和流(Flux)发生在当某土层土壤含水量(SW)大于DUL,通过一个给定的比例系数(SWcon)将多余的水分排至下一土层。非饱和流发生于某土层含水量小于 DUL时,作为对降水和蒸发的反应,在相邻的两个土层间以重力水或者水分扩散形式移动。模块将非饱和流作为相邻两个土层的平均含水量的函数进行计算。在SoilWater模块初始化中,可设置两个参数(diffus_const和diffus_slope)定义土层间的水分扩散力[38]。
土壤-作物蒸散(Evapotranspiration,ET)包括土壤蒸发(Evaporation)和植物蒸腾(Transpiration)两个过程,是农田水分循环和能量平衡的关键环节。作物蒸散量与不断变化的气象要素、根系吸水量和土壤含水量密切相关。APSIM模型采用参考作物蒸散量(ET0)和作物系数(Kc)估算农田潜在蒸散量(ETp),农田实际蒸散量(ETa)则取决于农田潜在蒸散量以及制约蒸散过程的土壤水分胁迫状况。在SoilWater模块中对土壤-作物蒸散的模拟基于土壤潜在蒸散原理和Penman-Monteith公式,并根据地表有机物残留和作物生长状况进行了修正[39]。在甘蔗全生育期中,从前期(稀疏冠层)到中期(冠层封闭)Kc由0.4增至1.25,随后下降至收获末期的0.7[40]。
3.2 土壤氮素模块
APSIM-Sugar模型对土壤氮素变化的模拟同样基于CERES模型[49]。甘蔗对土壤氮的吸收和转化是通过被动的蒸腾流和主动的根系吸收两个过程来进行模拟。所有与土壤-作物有机碳库和有机氮相关的氮素的变化都是需要利用SoilN模块进行计算。很多土壤氮素模型都力图较为全面地考虑土壤氮素的物理、生物化学以及物理化学等变化状态,以描述土壤氮素的流通转化状况。但复杂的机理性模型由于考虑的方面太多,反而可能影响到模型的模拟精度[42]。因此,SoilN模块采用了CERES-Maize模型的简化处理方式,将土壤中的各种类型有机质都设为具有相同的矿化率,降低了复杂运算可能造成的模拟精度误差,缺点是对土壤有机质的长期变化状况模拟欠佳[28]。
SoilN模块可描述土壤中碳素和氮素的动态变化特征,包括碳氮转化、有机质分解、硝化、反硝化、尿素水解等物理过程。该模块将土壤有机质分割为两个库:BIOM库包括易矿化有机物、土壤微生物及其代谢产物;HUM库则由剩下的有机物组成。两个库之间的碳通量可通过计算求得,而氮通量则由碳氮比来换算。BIOM的碳氮比被设定为一个常数,而HUM的碳氮比则是由输入的土壤属性初始值设定。BIOM和HUM的一级分解过程中,分解速率的稳定性取决于各土层的土壤温度和土壤水分。新鲜有机质(FOM)的分解速率则取决于碳氮比。有机氮的矿质化与腐殖化过程、微生物固定之间的平衡关系决定了土壤氮素的矿质化和固定的数量。如果土壤无机态氮(硝态氮和铵态氮)不能满足腐殖化和微生物的要求,则有机氮的分解速率也会下降。有机质分解产生的碳素一方面以CO2向大气中释放,另一方面则储存在BIOM和HUM中。土壤中的碳通量被定义为效率系数,或者描述为模块系统中的分配比例系数,土壤中的碳以不同的比例系数被合成到BIOM库中。由于土壤微生物的代谢作用,BIOM库中也存在内部的碳循环。在模拟过程中,每个土层的HUM和BIOM的数量可由设置输入值计算而来[38]。
4 APSIM-Sugar模型调参与验证
APSIM-Sugar模型的校验是通过一系列适合当地生产环境的气候、土壤、水分、养分、品种参数来阐明。为了使APSIM-Sugar模型能够正确模拟不同甘蔗品种在不同气候、土壤和水肥管理条件下的生长发育过程和产量形成过程,需要根据研究区域的甘蔗生长发育情况和气候土壤数据对甘蔗品种遗传参数和农田土壤初始水分氮素含量等进行必要的修正,即对模型进行本地化。
品种遗传参数由作物内在的生物学特性决定,通常是利用试验站点实测的作物生育期、产量、土壤以及同期气象数据,采用试错法(Trial and error),广义似然不确定性估计法(Generalized Likelihood Uncertainty Estimation,GLUE)或马尔可夫链蒙特卡洛法(Markov Chain Monte Carlo,MCMC)在计算机上进行多次调试来确定品种遗传参数的取值[35-36]。无论采用哪种算法,其基本原理方法均为:挑选一个参数而固定其他参数不变,在模型给定或根据参考文献确定的该参数范围内,每次增加或减少1%~10%,反复运行作物模型,直到模型模拟值(生育期、叶面积、生物量、产量等)与实测值的均方根误差(RMSE)或相对均方根误差(NRMSE)最小,且决定系数R2最接近1时,确定该参数值;然后再采用相同的方法确定其他参数,最后确定模型最优的参数组合。在APSIM模型土壤模块中,土壤水分的模拟采用了CERES的土壤分层阶梯式渗透模型和潜在水势基础模型[1,39]。土壤水分剖面、径流曲线、容重、饱和含水量、田间持水量、凋萎系数、排水系数、初始氮素含量等描述土壤水分和养分状况的参数是根据实测结果和试验区土壤特性来确定的。运行模型需要输入的土壤初始含水量和氮素含量,一般采用作物播种前后大田土壤水分和养分实测数据,如果没有实测数据,可以根据当地的土壤类型及组分进行估算或参考相关文献和模型默认值。
5 APSIM-Sugar模型评价与应用研究进展
综合前人研究,经过模型参数的本地化和适应性评价,APSIM-Sugar模型对地上部分甘蔗植株的叶面积、生物量、蔗茎和糖分产量模拟的预测值与实测值的相对均方根误差可以控制在5%~24%之间,决定系数R2可以控制在0.7~1之间[3,36]。目前,APSIM-Sugar模型已被广泛应用于不同气候带的甘蔗种植制度、种植结构、栽培管理、气候变化对甘蔗农田系统生产力和水肥利用效率的影响分析。
甘蔗产量预测和气候影响方面,Keating等[3]利用从世界范围内(如澳大利亚、南非、斯威士兰和美国夏威夷等)不同气候、不同管理、不同甘蔗品种的大田试验中获取的35个数据集,针对APSIM-Sugar模型的生物量和糖分模拟性能进行了评价,结果表明除了糖分积累对水分胁迫的响应不够精确以外,其他叶面积指数和生物量的模拟结果均比较准确,模型预测值与实测值的R2为叶面指数0.79,生物量0.93,蔗茎糖分产量0.83,氮素积累0.86。O’Leary等[10]的甘蔗模型比较研究显示APSIM-Sugar模型对绿叶干物重,新鲜蔗茎产量和蔗茎糖分的模拟值和实测值的RMSE分别为6.1、28.7和4.9 t/hm2。Cheeroo-Nayamuth等[27]利用校验后的APSIM-Sugar模型评价了毛里求斯的甘蔗潜在产量和可获得产量。2015年,Lisson等[34]基于澳大利亚昆士兰州两个蔗区的多年大田试验数据对APSIM-Sugar模型的10个品种遗传参数进行了敏感性分析,结果表明模拟蔗糖产量对太阳辐射利用效率、蔗茎糖分分配系数和绿叶数3个参数最为敏感,同时地区间的气候和土壤差异也会影响模型输出结果对品种遗传参数的敏感程度。2016年,Sexton等[35]进一步采用两种贝叶斯算法(GLUE和MCMC)对APSIM-Sugar模型品种遗传参数进行了调参验证,认为APSIM-Sugar模型可应用于甘蔗新品种的生态适应性风险评估。
甘蔗水肥利用效率方面,Inman-Bamber等[24,33,40,43-44]详细阐述了不同条件下甘蔗对土壤水分的利用情况,并利用APSIM-Sugar等甘蔗模型对甘蔗不同品种形态生理指标的变化对水分胁迫的响应进行了系统的研究。Stewart等[45]应用APSIM-Sugar模型评价了澳大利亚昆士兰州北部地区蔗田中硝态氮淋失的情况。Thorburn等[46-47]通过APSIM-Sugar模型模拟了氮肥的施用对澳大利亚甘蔗产业和生态环境的影响,提出了氮肥施用的优化策略。
6 总结与展望
作物生产系统是一个复杂的多因子系统,受气候、土壤、作物及栽培管理技术等因素的影响。在综合考虑这些因子的相互作用,预测和分析作物生长趋势等方面,基于土壤-作物系统和计算机信息技术的作物生长模拟模型有着其它工具不可替代的优势。现代作物生长模型的发展趋势,是越来越重视系统性和应用性的表现。APSIM模型作为农业生产系统模拟模型的代表,在世界范围内得到了广泛的应用。APSIM模型区别于其他模型的关键理念在于其模块化的“即插即用”的灵活设计,用户可以自行选择一系列的作物、土壤以及其他子模块来配置自己的作物模型。通过不同模块的分离与整合,使得APSIM能够很好地模拟不同土壤-作物系统的连作、轮作、间作以及农林混作效应。模块化设计的另一优势在于可以把零散的研究结果方便地集成到模型之中,使得某一学科领域的最新研究成果能方便地应用到别的学科领域。
以甘蔗模型为例,APSIM-Sugar模型的开发历程充分利用了APSIM模型的模块化设计优势,通过吸收和借鉴其他作物模型或模块的优点,并将甘蔗生理学、农艺栽培学、遗传育种学等相关学科的最新研究成果不断整合到模型当中,优化模块参数,改进模型模拟效果。根据前人研究,从总体上看,APSIM-Sugar模型模拟产量对太阳辐射、温度和土壤水分和氮肥的响应比较理想,特别是在海拔较低的平原丘陵地形和适宜的水肥条件下模拟效果较好,但是在高海拔山地和极端的水分、氮素和温度胁迫条件下,模拟效果不佳[3,24,40]。另外,由于国内甘蔗模型研究数据的缺乏,APSIM-Sugar模型对某些甘蔗品种(如我国的自育品种)和管理措施(如地膜覆盖、磷肥和钾肥的施用等)的模拟效果较差或没有进行模拟。需要通过更多的大田试验和实测数据对模型进行参数优化和模块改进。
综上所述,以APSIM模型为代表的土壤-作物农业生产系统模拟模型未来的发展趋势,一方面是与土壤学、气象学、遥感技术、地理信息系统(GIS)的结合[48],建立更加完善的作物生长模拟与农业生产计算机决策管理系统;另一方面是进一步与植物生理学、基因组学结合[49-50],将生理指标、表型性状和分子标记、等位基因信息通过数学模型和模型参数联系起来,预测遗传基因性状对环境和管理因子的响应。此外,近年来兴起的多种作物模型联合比较与改进国际合作项目(Agricultural Model Intercomparison and Improvement Project,AgMIP)也提供了一条非常好的发展道路[51-52]。总之,随着生态农业和数字化农业的发展,土壤-作物农业生产系统模拟模型的应用和发展将具有更加广阔的前景。
(References):
[1]McCown R L,Hammer G L,Hargreaves J N G,Holzworth D P,Freebairn D M.APSIM:a novel software system for model development,model testing and simulation in agricultural systems research[J].Agr Syst,1996,50:255-271
[2]Keating B A,Carberry P S,Hammer G L,Probert M E,Robertson M J,Holzworth D,Huth N I,Hargreaves J N G,Meinke H, Hochman Z.An overview of APSIM,a model designed for farming systems simulation[J].Eur J Agron.,2003,18(3):267-288
[3]Keating B A,Robertson,M J,Muchow,R C,Huth,N I.Modelling sugarcane production systems I:Development and performance of the sugarcane module[J].Field Crop Res.,1999,61:253-271
[4]Lisson S N,Robertson M J,Keating B A,Muchow R C.Modelling sugarcane production systems II:Analysis of system performance and methodology issues[J].Field Crop Res.,2000,68:31-48
[5]Monteith J L,Greenwood DJ.How do crops manipulate water supply and demand?[J]Philos Trans R Soc Lond A:Math Phys Eng Sci,1986,316(1537):245-259
[6]Sinclair T R.Water and nitrogen limitations in soybean grain production I.Model development[J].Field Crop Res.,1986,15:125-141
[7]Ritchie J T,Kiniry J R,Jones C A,Dyke P T.Model inputs.In:Jones,C A,Kiniry,J R(Eds.),CERES-Maize:A Simulation Model of Maize Growth and Development[M].Texas A&M University Press,College Station,1986:37-48
[8]Ritchie J T.Specifications of the ideal model for predicting crop yields.In:Muchow R C,Bellamy J A(Eds.),Climatic Risk in Crop Production:Models and Management for the Semi-arid Tropics and Subtropics[M].CAB International,Wallingford,1991:329-358
[9]Jones,C A;Wegener,M K;Russell,J S;McLeod,I M,Willians,J R.AUSCANE,Simulation of Australian Sugarcane with EPIC[M]. CSIRO,Brisbane,Australia,1989
[10]O'Leary G J.A review of three sugarcane simulation models with respect to their prediction of sucrose yield[J].Field Crop Res., 2000,68:97-111
[11]Holzworth D P,Huth N I,Devoil P G,Zurcher E J,Herrmann N I,Mclean G,Chenu K,Oosterom E J V,Snow V,Murphy C. APSIM-Evolution towards a new generation of agricultural systems simulation[J].Environ Modell Softw.,2014,62:327-350
[12]黄智刚,谢晋波.我国亚热带地区甘蔗产量的模型模拟[J].中国糖料,2007,29(1):8-12
[13]杨昆,蔡青,刘家勇,毛钧,范源洪.甘蔗生长模型研究进展[J].湖南农业大学学报:自然科学版,2015,41(1):29-34
[14]Inman-Bamber,N G,Temperature and seasonal effects on canopy development and light interception of sugarcane[J].Field Crop Res,1994,36:41-51
[15]Robertson M J,Bonnett G D,Hughes,R M,Muchow R C,Campbell J A.Temperature and leaf area expansion of sugarcane: integration of controlled-environment,field and model studies[J].Funct Plant Biol,1998,25:819-828
[16]Muchow R C,Spillman,M F,Wood A W,Thomas M R,Radiation interception and biomass accumulation in a sugarcane crop grown under irrigated tropical conditions[J].Australian Journal of Agricultural Research,1994,45:37-49
[17]Muchow,R C,Evensen C I,Osgood R V,Robertson M J.Yield accumulation in irrigated sugarcane II:Utilization of intercepted radiation[J].Agron J,1997,89:646-652
[18]Robertson M J,Wood A W,Muchow R C.Growth of sugarcane under high input conditions in tropical Australia I:Radiation use, biomass accumulation and partitioning[J].Field Crop Res,1996,48:11-25
[19]Muchow R C,Robertson,M J,Wood,A W,Growth of sugarcane under high input conditions in tropical Australia II:Sucrose accumulation and commercial yield[J].Field Crop Res,1996,48:27-36
[20]Inman-Bamber N G,Muchow R C,Robertson M J.Dry matter partitioning of sugarcane in Australia and South Africa[J].Field Crop Res,2002,76:71-84
[21]Glover J,The simultaneous growth of sugarcane roots and tops in relation to soil and climate[J].Proc S Afr Sug Technol,1967,46: 143-159
[22]Ball-Coelho B,Sampaio E V S B,Tiessen H,Stewart J W B,Root dynamics in plant and ratoon crops of sugar cane[J].Plant& Soil,1992,142:297-305
[23]Sinclair T R.Water and nitrogen limitations in soybean grain production I:Model development[J].Field Crops Res,1986,15: 125-141
[24]Inman-Bamber N G,Mcglinchey M G.Crop coefficients and water-use estimates for sugarcane based on long-term Bowen ratio energy balance measurements[J].Field Crop Res,2003,83:125-138
[25]Catchpoole V R,Keating B A.Sugarcane yield and nitrogen uptake in relation to profiles of mineral-nitrogen in the soil[C].//Proc Aust Soc Sugar cane Technol,1995:187-192
[26]Robertson M J,Muchow R C,Wood A W,Campbell J A.Accumulation of reducing sugars by sugarcane:effects of crop age, nitrogen supply and cultivar[J].Field Crop Res,1996,49:39-50
[27]Cheeroo-Nayamuth F C,Robertson M J,Wegener M K,Nayamuth A R H,Using a simulation model to assess potential and attainable sugar cane yield in Mauritius[J].Field Crop Res,2000,66:225-243
[28]Probert M E,Dimes J P,Keating B A,Dalal R C,Strong W M.APSIM's water and nitrogen modules and simulation of the dynamics of water and nitrogen in fallow systems[J].Agr Syst,1998,56:1-28
[29]Singh G,Chapman S C,Jackson P A,Lawn R J,Lodging reduces sucrose accumulation of sugarcane in the wet and dry tropics[J]. Australian Journal of Agricultural Research,2002,53:1183-1195
[30]Inman-Bamber N G,Attard S J,Spillman M F,Hogarth D M.Can lodging be controlled through irrigation?[C].//Conference of the Australian Society of Sugar Cane Technologists,2004:1-11
[31]Stokes C J,Inman-Bamber N G,Everingham Y L,Sexton J.Measuring and modelling CO2effects on sugarcane[J].Environ Modell Softw,2016,78:68-78
[32]Jackson P A,Basnayake J,Inman-Bamber N G,Lakshmanan P,Natarajan S,Stokes C,Genetic variation in transpiration efficiency and relationships between whole plant and leaf gas exchange measurements in Saccharum spp.and related germplasm[J].J Exp Bot,2016,39:44-56
[33]Inman-Bamber N G,Jackson P A,Stokes C,Verall S,Lakshmanan P,Basnayake J.Sugarcane for water-limited environments: Enhanced capability of the APSIM sugarcane model for assessing traits for transpiration efficiency and root water supply[J].Field Crop Res,2016,TBA
[34]Lisson S N,Inman-Bamber N G,Robertson M J,Keating B A,The historical and future contribution of crop physiology and modelling research to sugarcane production systems[J].Field Crop Res,2005,92:321-335
[35]Sexton J,Everingham Y L,Inman-Bamber N G.A global sensitivity analysis of cultivar trait parameters in a sugarcane growth model for contrasting production environments in Queensland,Australia[J].Eur J Agron,2015
[36]Sexton J,Everingham Y L,Inman-Bamber N G.A theoretical and real world evaluation of two Bayesian techniques for the calibration of variety parameters in a sugarcane crop model[J].Environ Modell Softw,2016,83:126-142
[37]Monteith J L,Unsworth M H.Principles of Environmental Physics(second edition),in Legg B J(Eds).Experimental Agriculture[M]. Sevenoaks,Kent,Edward Arnold,London,1990
[38]Model,Crop and Soil[EB/OL]in Agricultural Production Systems Simulator(APSIM)official website:http://www.apsim.info/ Documentation/Model,CropandSoil.aspx
[39]Ritchie J T.Model for Predicting Evaporation From a Row Crop With Incomplete Cover[J].Water Resour Res,1972,8:1204-1213 [40]Inman-Bamber N G,Smith D M.Water relations in sugarcane and response to water deficits[J].Field Crop Res,2005,92:185-202
[41]Allen R G,Pereira L S,Raes D,Smith M.FAO.Crop Evapotranspiration:Guidelines for Computing Crop Water Requirements[M]. In:Irrigation and Drainage Paper 56.FAO,Rome,1998
[42]Willigen P D,Nielsen N E,Claassen N,Castrignanò A M.Modelling Water and Nutrient Uptake[M].Root Methods.Springer Berlin Heidelberg,2000
[43]Inman-Bamber N G,Everingham Y,Muchow R C.Modelling water stress response in sugarcane:Validation and application of the APSIM-Sugarcane model[J].Australian Agronomy Conference,2001:1030-1200
[44]Inman-Bamber N G,Lakshmanan P,Park S.Sugarcane for water-limited environments:Theoretical assessment of suitable traits[J]. Field Crop Res,2012,134:95-104
[45]Stewart L K.,Charlesworth P B,Bristow K L.Estimating deep drainage and nitrate leaching from the root zone under sugarcane using APSIM-SWIM[J].Agricultural Water Management,2006,81:315-334
[46]Thorburn P,Biggs J,Bristow K,Horan H,Huth N,Unkovich M,O'Leary G.Benefits of sub-surface application of nitrogen and water to trickle irrigated sugarcane[C].//Solutions for A Better Environment:Proceedings of the Australian Agronomy Conference, Geelong,Victoria,Australia,2003:2-6
[47]Thorburn P J,Biggs J S,Attard S J,Kemei J.Environmental impacts of irrigated sugarcane production:Nitrogen lost through runoff and leaching[J].Agr Ecosyst Environ,2011,144:1-12
[48]Davis R J,Baillie C P,Schmidt E J.Precision agriculture technologies-relevance and application to sugarcane production[J]. Agricultural Technologies in a Changing Climate,2009:13-16
[49]Yin X,Struik P C,Kropff M J.Role of crop physiology in predicting gene-to-phenotype relationships[J].Trends in Plant Science, 2004,9(9):426-432
[50]Hoogenboom G,White J W,Messina C D.From genome to crop:integration through simulation modeling[J].Field Crop Res,2004, 90(1):145-163
[51]Rosenzweig C,Hatfield J,Jones J W,Ruane A C.The Agricultural Model Intercomparison and Improvement Project(AgMIP): Overview and Progress[J].American Geophysical Union,2012,170:166-182
[52]Rosenzweig C,Jones J W,Hatfield J L,Ruane A C,Boote K J,Thorburn P,Antle J M,Nelson G C,Porter C,Janssen S.The Agricultural Model Intercomparison and Improvement Project(AgMIP):Protocols and pilot studies[J].Agricultural&Forest Meteorology,2013,170:166-182
Modular Design and Application of Agricultural System Simulating Model for Sugarcane(APSIM-Sugar)
MAO Jun1,2,N.G.Inman-Bamber3,YANG Kun1,2,LU Xin1,2,LIU Jia-yong1,2,P.A.Jackson4,FAN Yuan-hong1,2*
(1.Sugarcane Research Institute,Yunnan Academy of Agricultural Sciences,Kaiyuan 661699;2 Yunnan Key Laboratory of Sugarcane Genetic Improvement,Kaiyuan 661699;3 College of Science,Technology and Engineering,James Cook University,Townsville, Australia;4 CSIRO Agriculture,Australian Tropical Science and Innovation Precinct,Townsville,Australia)
Using typical case of APSIM-sugar as an example,the Agricultural Production System simulator (APSIM)developed by the Agricultural Production Systems Research Unit(APSRU)in Australia was introduced, including the core crop and soil modules,the related process theories,parameter calibration and model validation and application research progress.We aimed to provide some references for learning,understanding and using the APSIM in soil-plant system simulating researches.
sugarcane;agricultural production system simulator;physiological process;module;parameter
S566.3
B
1007-2624(2017)01-0044-07
10.13570/j.cnki.scc.2017.01.016
2016-09-22
云南省高端科技人才引进计划项目(2012HA001)。
毛钧(1979-),男,硕士,副研,主要从事甘蔗种质资源利用与甘蔗模型研究。E-mail:mj_raincat@163.com
范源洪,研究员,主要从事甘蔗种质资源和遗传育种研究。E-mail:fyhysri@vip.sohu.com
猜你喜欢
儿童故事画报·自然探秘(2022年12期)2022-11-24 10:10:12
中国糖料(2022年1期)2022-01-13 12:28:06
基层中医药(2021年3期)2021-11-22 08:08:04
甘蔗糖业(2020年1期)2020-03-04 03:48:26
特别健康(2018年3期)2018-07-04 00:40:08
农村百事通(2018年5期)2018-03-28 10:29:26
中国糖料(2016年1期)2016-12-01 06:48:57
中国糖料(2012年3期)2012-09-03 08:46:16
植物营养与肥料学报(2011年5期)2011-11-06 07:30:52
植物营养与肥料学报(2011年2期)2011-10-26 03:52:10
