
遮荫对元宝枫花色素苷含量的影响
2017-03-27 09:13韦忠刚
天津农业科学 2017年3期
关键词:元宝枫
韦忠刚
摘 要:为研究遮荫强度对元宝枫花色素苷含量的影响,以秋叶变红元宝枫为试验材料,通过进行不同强度的遮荫处理,测定元宝枫不同位置叶片中花色素苷含量和PAL活性的变化。结果表明:随着遮荫时间的延长,元宝枫上部和中部叶片花色素苷含量呈现出持续下降趋势,下部叶片呈现出先升高后降低的趋势;PAL酶活性均呈现出持续降低趋势。在园林应用中适当对元宝枫进行修剪整枝有利于秋季叶片着色。
关键词:元宝枫;花色素苷;苯丙氨酸解氨酶
中图分类号:S792.35 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2017.03.002
Effects of Shading on the Content of Anthocyanin in Acer truncatum
WEI Zhonggang
(Agricultural Integrated Service Center of Tai'an, Tai'an, Shandong 271000, China)
Abstract: In order to study the effects of shading intensity on the content of anthocyanin in Acer truncatum, the changes of anthocyanin content and PAL activity in leaves of A. truncatum were studied by different shading treatments. The results showed that the content of anthocyanin in the upper and middle leaves of A. truncatum showed a decreasing trend with increasing the shading time, while the lower leaves showed a trend of increasing first and then decreasing. The activities of PAL decreased continuously.
Key words: A. truncatum;anthocyanin;PAL
元寶枫(Acer truncatum),槭树科槭树属,我国特有树种,分布范围广,变异丰富。元宝枫以其独特的叶形、叶色等特点受到广泛关注,是我国重要的园林绿化树种和彩色叶树种[1]。研究发现,元宝枫的种子含油量较高,如何高效提取和利用元宝枫籽油也成为当下研究热点,被视为重要的木本油料树种,发展前景广阔。目前,元宝枫已被视为集观赏价值、经济价值于一身的优良树种。
植物的叶色变化是遗传因素和外界环境共同作用的结果,就生态因子来说,主要集中在光照、温度、水分、栽培方式几个方面,基于不同物种的适应性展现出不同的表现形式。研究发现:黄金榕随着光强的增加颜色愈加鲜艳,而紫叶黄栌随着光强的增加叶色则逐渐暗淡;蓝光有利于美人梅叶片的着色,紫外光有利于高山花卉的着色;紫叶矮樱需要适度的干旱才能很好地显色,而天南星科植物却需要在高度湿润的环境下才能表现良好[2]。研究表明,在植物的变色过程中花色素苷种类与含量发生了变化,这是导致叶色呈现不同形式的内在因素[3]。同时,花色素苷在提高植物抗性方面也起到了重要作用,在寒冷、干旱等胁迫条件下,植物可以合成花色素苷抵御逆境[4]。
在植物叶色变化过程中,植物的光合作用是关键,而光强起到了不可替代的作用[5]。随着雾霾越来越严重,高楼耸立的城市中遮阴度不断升高,城市绿化中的观赏树木很少在全光照下生长,这对城市观赏植物的生长起到了抑制作用,尤其是对变色叶植物的生长,降低了彩色植物的美化功能。本试验通过对元宝枫光照强度的控制,研究了不同光照条件下元宝枫叶片花色素苷含量及其关键酶苯丙氨酸解氨酶活性的变化,探索光照强度与元宝枫叶色变化的关系,以期为元宝枫的引种利用提供参考。
1 材料和方法
1.1 试验时间、地点
本试验于2015年10月在泰安市泰山区邱家店邱家店苗圃进行。
1.2 试验材料
供试材料为课题组选育的元宝枫秋叶变红无性系植株。
1.3 试验方法
1.3.1 材料处理 将盆栽嫁接苗放于试验地,用遮阳网进行遮阳处理。处理1,一层遮阳网;处理2,两层遮阳网;以全光照为对照。每处理20盆,经光强测定仪测定,一层遮阳网的光强约为全光强的65%,两层遮阳网的光强约为全光强的30%。分别在处理后的第0,7,14天,取植株的上、中、下三个部位的叶片进行花色素苷含量和苯丙氨酸解氨酶活性的测定。
1.3.2 花色素苷含量的测定 取叶片鲜样0.3 g,加入 10 mL 1%盐酸乙醇提取液,放置24 h,过滤,用紫外光分光光度计在波长 535 nm 下测定吸光度。
1.3.3 苯丙氨酸解氨酶活性测定 参照文献[6],取叶片鲜样0.5 g,加 5 mL 0.1 mol·L-1 硼酸缓冲液(pH值 8.8)、巯基乙醇 2 mmol·L-1、0.5 g PVP,冰浴研磨匀浆,12 000 r·min-1 4 ℃离心 15 min ,取上清夜。吸取 1 mL 0.02 mol·L-1 的 L-PAL 和 3 mL pH值8.8 硼酸缓冲液(对照为失活的酶液),置于试管中,水浴中保温 30 min,加0.2 mL 酶液,混合均匀, 30 ℃恒温水浴中反应 30 min,在冰浴中终止反应,于分光光度计 290 nm 上测其 OD 值。计算公式如下:
PAL 活性(U)=OD×1 000
1.4 数据处理
数据采用EXCEL、SPSS13.0进行分析处理。
2 结果与分析
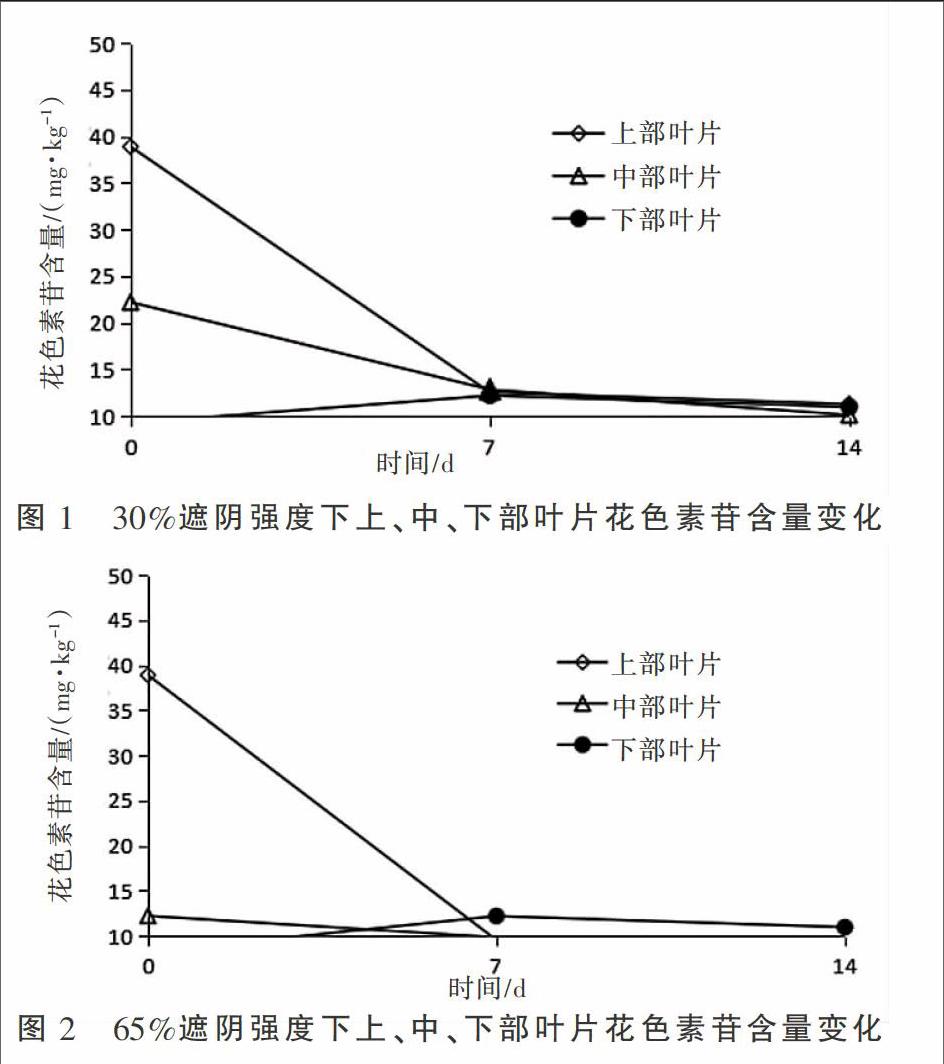
2.1 30%遮阴条件下花色素苷含量的变化
元宝枫在30%遮阴条件下花色素苷含量的变化见图1。随着遮阴时间的延长,元宝枫上部和中部叶片花色素苷含量均呈现出持续下降趋势,同时,叶片受光强的影响从上到下逐渐减轻。其中元宝枫上部叶片花色素苷含量在处理第7,14 天分别下降了67.8%和71.1%,中部叶片花色素苷含量在处理第7,14天分别下降了42.3%和54.5%,而下部叶片则呈现出先升高再降低的趋势。
2.2 65%遮阴条件下花色素苷含量的变化
元宝枫在65%遮阴条件下花色素苷含量的变化见图2。与元宝枫在30%遮阴条件下花色素苷含量的变化相似,随着遮阴时间的延长,元宝枫上部和中部叶片花色素苷含量均呈现出持续下降趋势,降低强度更大,叶片受光强的影响从上到下逐渐减轻。其中元宝枫上部叶片花色素苷含量在处理第7,14 天分别下降了75.5%和78.7%,中部叶片花色素苷含量在处理第7,14 天分别下降了19.5%和41.5%,而下部叶片也呈现出先升高再降低的趋势。可见,在高遮阴条件下,上部叶片受影响较大,当植物适应光强后,花色素苷含量变化较小,植物合成花色素苷含量随光强降低而下降,达到一定程度后会稳定在某一水平。
2.3 30%遮阴条件下PAL酶活性的变化
元宝枫在30%遮阴条件下PAL酶活性变化见图3。随着遮阴时间的延长,元宝枫上、中部叶片PAL酶活性呈现出先升高后降低的趋势,下部叶片呈现持续降低趋势,其中,上部和中部叶片中苯丙氨酸解氨酶活性显著高于下部叶片。同时,叶片受光强的影响从上到下逐渐减轻,其中元宝枫上、中、下部叶片PAL酶活性在处理第7 天分别上升了32.3%,1.3%和12.6%,在处理第14 天分别下降了40.1%,20.5%和24.9%。可见,植物在遭受光强不足时,会合成PAL酶以应对不良环境条件,当超过一定承受阈值时,保护功能降低,酶活性减弱。
2.4 65%遮阴条件下PAL酶活性的变化
元宝枫在65%遮阴条件下PAL酶活性变化见图4。随着遮阴时间的延长,元宝枫上、中、下部叶片PAL酶活性呈现持续降低的趋势,其中,上部和下部叶片中苯丙氨酸解氨酶活性降低趋势最明显,中部叶片變化较小。元宝枫上、中、下部叶片PAL酶活性在处理第7 天分别下降了24.7%,4.5%和16.8%,第14 天分别下降了54.6%,9.7%和55.7%。
3 结论与讨论
变色叶植物在园林绿化应用中起到了重要作用,而花色素苷则是植物变色过程中不可或缺的因素。研究还发现,花色素苷在植物的逆境胁迫中也起到了重要作用[7]。Moscovici[8]研究发现,植物在光强下,可以通过合成花色素苷消耗过多的能量,避免强光辐射带来的不利影响。Parker[9]和Krol等[10]研究发现,常春藤在一年的生长中,其叶片POD、SOD等抗氧化酶活性和花色素苷含量均随温度的降低而升高,可以推测,花色素苷在植物的抗寒过程中也起到了一定作用。已有研究证明,花色素苷也有利于植物的抗旱和抵抗渗透胁迫的能力[11]。
苯丙氨酸解氨酶是植物苯丙氨酸代谢途径的关键酶,对花色素苷的合成有重要的调节作用,然而,对于两者的关系尚有争议。Chalker-scott等[12]研究认为PAL活性与花色素苷含量呈正相关,而Sherwin和Farraont[13]则认为两者并无明确的联系,其原因可能是不同植物在不同生长期和环境胁迫下没有表现出明确的联系。本研究通过试验表明,元宝枫变色过程中PAL活性与花色素苷含量没有明确的定量关系。
光照条件是影响花色素苷形成的重要因素,其中光强最为重要。梁峰[5]研究发现,元宝枫在全光照条件下颜色最鲜艳,观赏效果最好,叶绿素含量和花色素苷含量均最高。一般认为,低温加速了叶绿素的降解,植物在形成一系列抗氧化酶的过程中花色素苷含量也不断升高,花色素苷在抵抗低温胁迫中起到了一定作用。本研究发现,随着遮荫时间的延长,元宝枫叶片的花色素苷含量和PAL酶活性均下降,上、中、下部叶片降低程度具有差异性。
笔者研究了不同光照条件下元宝枫叶片中花色素苷含量和苯丙氨酸解氨酶活性的变化,同时对枝条不同层次的叶分别进行了比较,研究发现,随着遮阴时间的增加,二者皆表现出下降的趋势,然而不同位置的叶片变化速率不同。因此,通过筛选适宜的彩叶植物、对树木进行修剪、补光等可以有效延长彩叶植物叶片变色期,提高植物的观赏特性。
参考文献:
[1]朱书香,杨建民,王中华,等.4种李属彩叶植物色素含量与叶色参数的关系[J].西北植物学报,2009,29(8):1663-1669.
[2]洪丽,王金刚,龚束芳.彩叶植物叶色变化及相关影响因子研究进展[J].东北农业大学学报,2010,41(6):152-156.
[3]杨科家,丰震,朱红梅,等.元宝枫叶片花色素苷的提取及其稳定性研究[J].中国农学通报,2009,25(24):334-337.
[4]姜卫兵,庄猛,韩浩章,等.彩叶植物呈色机理及光合特性研究进展[J].园艺学报,2005,32(2):352-358.
[5]梁峰,蔺银鼎.光照强度对彩叶植物元宝枫叶色表达的影响[J].山西农业大学学报(自然科学版),2009,29(1):41-45.
[6]赵世杰.植物生理学实验指导[M].北京:中国农业科技出版社,1998:55-57.
[7]唐前瑞,陈友云,周朴华.红继木花色素苷稳定性及叶片细胞液 pH 值变化的研究[J].湖南林业科技,2003,30(4):24-25.
[8]MANCINELLI A L,YANG C P,RABINO I,et al.Photocontrol of anthocyanin synthesis:V.further evidence against the involvement of photosynthesis in high irradiance reaction anthocyanin synthesis of young seedlings[J].Plant Physiology, 1976, 58(2):214-217.
[9]PARKER J. Relationships among cold hardiness, water soluble protein, anthocyanins and free sugar in Hedera helix L[J]. Plant physiol, 1962, 37: 809-813.
[10]KROL M, GRAY G, HURRY V. Low-temperature stress and photoperiod affect an increased tolerance to photoinhibition in Pinus banksiana seedlings[J]. Can bot, 1995, 73:1119-1127.
[11]冯立娟,苑兆和,尹燕雷,等.美国红枫叶色表达期间相关物质的研究[J].山东林业科技,2008(4):9-10.
[12] CHALKER-SCOTT L. Environmenttal significance of anthocyanins in plant stress responses[J].Photochemistry and photobiology, 1999, 70(1): 1-9.
[13]SHERWIN H W,FARRANT J M.Protection mechanisms against excess light in the resurrection plants Craterostigma wilmsii and Xerophyta viscosa[J].Plant Growth Regulation,1998,24(3):203-210.
猜你喜欢
中国饲料(2022年5期)2022-04-26
中国油脂(2022年2期)2022-03-10
中国林业产业(2021年5期)2021-12-14
绿色科技(2021年3期)2021-11-28
中国生物防治学报(2021年1期)2021-04-18
现代园艺(2020年19期)2020-12-20
山东林业科技(2019年2期)2019-06-03
中国粮油学报(2018年12期)2018-03-19
农村.农业.农民(2016年24期)2016-04-02
