
大猿叶虫滞育品系和非滞育品系交配行为的比较
2017-03-16 08:02周睿琦谈倩倩王小平
环境昆虫学报 2017年1期
周睿琦,谈倩倩,王小平,刘 文
(华中农业大学植物科学技术学院,昆虫资源利用与害虫可持续治理湖北省重点实验室,武汉 430070)
大猿叶虫滞育品系和非滞育品系交配行为的比较
周睿琦,谈倩倩,王小平,刘 文*
(华中农业大学植物科学技术学院,昆虫资源利用与害虫可持续治理湖北省重点实验室,武汉 430070)
大猿叶虫ColaphellusbowringiBlay是一种重要的十字花科蔬菜害虫,以成虫越冬和越夏。明确大猿叶虫非滞育品系生殖力下降的现象是否与交配行为的改变相关,将促进该非滞育品系在滞育遗传、滞育调控分子机制研究中的应用。本研究在25℃和L ∶D=12 ∶12 h条件下,测定了大猿叶虫滞育品系和非滞育品系的交配节律、交配持续时间和交配频次。结果表明,大猿叶虫滞育品系和非滞育品系交配高峰均出现在光期开始后2 h,非滞育品系的交配比率显著低于滞育品系。滞育品系和非滞育品系第1次交配持续时间无显著差异,分别为122.7±7.4 min和100.1±10.2 min;但非滞育品系的总交配持续时间显著低于滞育品系。非滞育品系的交配频次为2.2 ±0.3次,显著低于滞育品系的3.8±0.5次。滞育品系和非滞育品系交配行为的比较发现,大猿叶虫非滞育品系生殖力降低与其交配比率下降、交配持续时间缩短和交配频次减少显著相关,本研究为进一步利用非滞育品系为材料解析滞育遗传、滞育调控机制奠定了基础。
大猿叶虫;非滞育品系;交配节律;交配持续时间;交配频次
滞育(Diapause)是昆虫应对外界不良环境的一种季节性适应策略,主要表现为营养代谢模式的转变和发育停滞(Tauberetal.,1986)。滞育昆虫具有遗传多样性,其滞育能力(diapause capacity)可被自然或人工选择改变。通过自然选择或人工筛选已在40多种昆虫和螨类中获得非滞育品系(Tauberetal.,1986;Riihimaa and Kimura,1988;Ito,2009)。有的昆虫非滞育品系已被用于滞育遗传研究的材料,例如,比较始红蝽Pyrrhocorisapterus滞育品系和非滞育品系杂交、回交后亲代和子代的光周期反应,最终探明了始红蝽的滞育综合征(diapause syndrome)是显性性状(Socha and Hodková,1994)。有的昆虫非滞育品系可被用于害虫遗传防治,田间释放害虫的非滞育品系,非滞育品系个体与野生品系个体杂交后,可改变害虫的基因型,在不利环境到来时,害虫无法进入滞育而死亡(Hoy and Knop,1978)。然而,很多昆虫非滞育品系存在适应性退化的现象,其中生殖力降低最为明显(Tauberetal.,1986)。例如,在守瓜Atrachyamenetriesi中,非滞育品系雌成虫的产卵量下降,卵的孵化率降低(Ando and Miya,1968)。交配是两性生殖昆虫繁衍后代的重要环节,交配节律、交配持续时间和交配频次均可能影响昆虫的生殖力(Arnqvist and Nilsson,2000;Herbersteinetal.,2010)。有研究表明,昆虫非滞育品系的生殖力降低可能与交配行为的改变有关(Herbersteinetal.,2010),但非滞育品系的哪些交配行为发生改变进而影响到其生殖力尚不清楚。
大猿叶虫ColaphellusbowringiBaly是一种重要的十字花科蔬菜害虫,低温或长光照会诱导其成虫入土滞育(薛芳森等,2002;Xueetal.,2002)。前期报道过一个对光周期不敏感的大猿叶虫非滞育品系,该非滞育品系在温度高于20℃时,无论何种光周期均不能诱导个体进入滞育(Maetal.,2011;Tanetal.,2016)。大猿叶虫非滞育品系是研究滞育遗传的良好材料,利用大猿叶虫滞育品系与非滞育品系杂交,已探明光周期诱导的滞育是不完全显性遗传且受母本效应影响较大(Kuangetal.,2011;Maetal.,2011)。然而,大猿叶虫非滞育品系的生殖力明显降低,其雌成虫寿命、产卵量和卵孵化率均显著低于正常品系(Tanetal.,2016)。前期观察发现,大猿叶虫交配行为中的交配频次和交配持续时间显著影响着其生殖力,交配频次与其产卵量正相关,较长的交配持续时间可提高产卵量(Liuetal.,2010;Liuetal.,2013)。因此,推测大猿叶虫非滞育品系生殖力的降低可能与其交配行为的改变有关,但非滞育品系的交配行为与滞育品系相比存在哪些差异仍不清楚。
为明确大猿叶虫非滞育品系生殖力降低的原因,在25℃和L ∶D=12 ∶12 h条件下,分别测定大猿叶虫滞育品系和非滞育品系的交配节律、交配持续时间和交配频次。研究结果可明确大猿叶虫非滞育品系生殖力降低与其交配行为发生改变的相关性,为以非滞育品系为材料研究昆虫滞育遗传、滞育调控机制提供科学依据。
1 材料与方法
1.1 供试昆虫
2008年11月在江西省修水县(29°1′N,114°4′E)采集超过1000头的大猿叶虫成虫,在自然条件下的玻璃缸中入土滞育(Xueetal.,2002)。大猿叶虫成虫滞育解除后子代有明显的光周期反应,温度高于20℃时长光照诱导其进入滞育,视为滞育品系(high-diapause strain,HD strain)。大猿叶虫滞育解除后,将成虫转移到塑料养虫盒(16 cm × 7.5 cm × 6 cm)内繁殖,用于试验。
2006年11月在江西省修水县(29°1′N,114°4′E)采集500-600头大猿叶虫成虫,将滞育解除后的成虫饲养在非滞育诱导条件下(22℃和L ∶D=13 ∶11),部分成虫未进入滞育。将未滞育成虫饲养在滞育诱导条件22℃和L ∶D=16 ∶8下,持续饲养筛选22代后,滞育比率低于5%,对光周期诱导失去敏感性,获得了较稳定的非滞育品系(non-diapause strain,NDS)(Maetal.,2011),该非滞育品系在室内22℃和L ∶D=16 ∶8 h条件下连续饲养保存。
1.2 试验条件
所有幼虫或成虫放入塑料养虫盒内,以新鲜萝卜Raphanusstativusvar.longipinnatus叶饲养。饲养条件为温度25℃±1℃、光周期L ∶D=12 ∶12 h、相对湿度50%-60%、光照强度500-700 Lx(薛芳森等,2002)。饲养的全过程均在光照培养箱(HP250GS,武汉瑞华仪器设备有限责任公司)中进行,暗期处理采用人工套袋(在养虫盒外加套不透光的黑布袋,至光期再移除)的方法进行(Xueetal.,2002)。
1.3 交配行为观察
将单头滞育品系或非滞育品系5日龄未交配雌成虫放入垫有保湿滤纸和新鲜萝卜叶的培养皿(直径 5.0 cm)中,适应10 min后,引入1头同品系的5日龄未交配雄成虫。大猿叶虫交配多发生在光期,雌雄成虫可多次交配,且第1次交配持续时间显著长于之后的交配持续时间(Liuetal.,2010)。配对后立即观察成虫的交配情况,观察完整的光期,从9 ∶00开始至21 ∶00结束,连续观察12 h。每对雌雄成虫为1次重复,滞育品系和非滞育品系观察样本分别为30对和40对雌雄成虫。记录每对雌雄成虫的交配起始时间、交配结束时间、交配频次(Liuetal.,2010),同一对雌雄成虫的总交配持续时间为其多次交配持续时间之和。
1.4 数据分析
试验中的数据均采用SPSS 11.5进行分析。对大猿叶虫滞育品系和非滞育品系雌雄成虫观察对数和交配对数进行卡方检验,比较二者间交配比率的差异;交配持续时间、交配频次的平均值差异通过t测验(t-test,P<0.05)进行分析。

图 1 大猿叶虫滞育品系和非滞育品系的交配节律Fig.1 The mating rhythm of the diapause strain (DS)and non-diapause strain (NDS)of Colaphellus bowringi
2 结果与分析
2.1 交配节律
在25℃和L ∶D=12 ∶12 h条件下,大猿叶虫滞育品系和非滞育品系的交配高峰均出现在光期开始后2 h,交配率分别为76.7%和30.0%。滞育品系的交配率为90.0%,非滞育品系为47.5%,大猿叶虫滞育品系的交配率显著高于非滞育品系(χ2=42.128,P<0.01)(图1)。
2.2 交配持续时间

图2 大猿叶虫滞育品系和非滞育品系交配持续时间的分布Fig. 2 The distribution of the mating duration of diapause strain (DS)and non-diapause strain (NDS)of Colaphellus bowringiA,第一次交配持续时间;B,总交配持续时间。A, mating duration for the first time; B, total mating duration.
大猿叶虫滞育品系和非滞育品系第1次交配持续时间的分布相似,超过60%的个体第1次交配持续时间大于90 min(图2A)。大猿叶虫滞育品系和非滞育品系总交配持续时间分布存在明显差异,滞育品系总交配持续时间最短为73 min,最长达到417 min;而非滞育品系总交配持续时间集中在90-150 min(图2B)。滞育品系第1次交配持续时间为122.7±7.4 min,非滞育品系的第1次交配持续时间为100.1±10.2 min,两品系第1次交配持续时间无显著性差异(t=1.847,P=0.072)(图3A);大猿叶虫滞育品系总交配持续时间为232.6±18.7 min,非滞育品系总交配持续时间为152.3±20.1 min,滞育品系总交配持续时间显著长于非滞育品系(t=2.877,P<0.01)(图3B)。
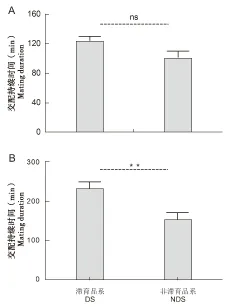
图3 大猿叶虫滞育品系和非滞育品系交配持续时间的比较Fig.3 The comparison of mating duration between diapause strain (DS)and non-diapause strain (NDS)of Colaphellus bowringi注:A,第一次交配持续时间;B,总交配持续时间。Note:A, mating duration for the first time; B, total mating duration.
2.3 交配频次
大猿叶虫滞育品系和非滞育品系交配频次的分布存在明显差异,大猿叶虫滞育品系大部分个体1天中交配频次大于3次,而非滞育品系大部分个体1天中交配1-2次。大猿叶虫滞育品系总交配频次为3.8±0.5次,而非滞育品系总交配频次为2.2±0.3次,滞育品系交配频次显著多于非滞育品系(t=2.481,P< 0.001)(图4)。

图4 大猿叶虫滞育品系和非滞育品系平均交配频次Fig.4 Meanfrequency of mating between diapause strain (DS) and non-diapause strain (NDS)of Colaphellus bowringi
3 结论与讨论
昆虫非滞育品系在滞育遗传研究和害虫生物防治中有潜在应用价值,但大部分昆虫非滞育品系存在生殖力降低的现象(Tauberetal.,1986)。本研究对大猿叶虫滞育品系和非滞育品系交配行为的比较发现,滞育品系和非滞育品系的交配节律无明显差异,但非滞育品系的交配率显著下降、总交配持续时间显著缩短,交配频次显著减少,说明非滞育品系的交配能力较之滞育品系显著降低,这为明确大猿叶虫非滞育品系生殖力降低的内在机制奠定了基础。
大猿叶虫交配多发生在光期,雌雄成虫可多次交配,第1次交配持续时间显著长于之后的交配持续时间(Liuetal.,2010)。本研究表明,大猿叶虫滞育品系和非滞育品系第1次交配持续时间无显著差异,但非滞育品系的总交配持续时间显著短于滞育品系。在昆虫中,交配持续时间的长短对其生殖适应性有重要影响(Herbersteinetal.,2010)。在多数昆虫中,雄成虫通过延长交配持续时间来增加可向雌成虫转移的精子数,同时延长交配持续时间可作为一种配偶保护行为,避免精子竞争(Thornhill and Alcock,1983;Simmons,2001);对于雌成虫,延长交配持续时间有利于贮存更多的精子和随精子一同转移的营养物质进而提高生殖成功的几率(Arnqvist and Nilsson,2000)。交配持续时间对产卵量的影响在许多昆虫中已有报道。例如对于茄28星瓢虫Henosepilachnavigintloctopunctata,较长的交配持续时间导致更高的产卵量和卵孵化率(周康念等,2012)。因此,大猿叶虫非滞育品系总交配持续时间的缩短,可能导致雌成虫从雄成虫精液获得的营养物质和精子数减少,不利于雌成虫的生殖,最终导致其非滞育品系雌成虫产卵量的下降。
许多昆虫在一个生殖季节中能发生多次交配(multiple mating),这种多次交配常有利于提高雌成虫的生殖适应性(Arnqvist and Nilsson,2000)。雌成虫通过多次交配能获得更多的营养进而刺激生殖行为,提高生殖产量;同时,多次交配也有利于雌成虫储存足够的精子,确保所有卵的受精(Thornhill and Alcock,1983;Arnqvist and Nilsson,2000;孟玲和李保平,2006)。在大猿叶虫中,雌成虫交配频次与其产卵量呈正相关(Liuetal.,2013)。大猿叶虫非滞育品系1天内交配频次显著少于滞育品系,交配频次的减少与其总交配持续时间的缩短有直接关系。因此,交配频次减少可能是导致大猿叶虫非滞育品系生殖力降低的关键因素。
本研究表明大猿叶虫非滞育品系较之滞育品系,具有低的交配率、短的交配持续时间、少的交配频次,这些均有可能导致非滞育品系雌成虫的生殖力下降,从而导致非滞育品系适应性降低。明确大猿叶虫非滞育品系生殖力降低的原因与其交配行为发生改变的相关性,为进一步利用大猿叶虫非滞育品系为材料分析昆虫滞育遗传、滞育调控的机制提供了依据。
References)
Ando Y, Miya K.Diapause character in the false melon beetle,AtrachyamenetriesiFaldermann, produced by crossing between diapause and non diapause strains [J].BulletinoftheFacultyofAgricultureIwateUniversity, 1968, 9: 87-96.
Arnqvist G, Nilsson T. The evolution of polyandry: Multiple mating and female fitness in insects [J].AnimalBehavior, 2000, 60 (2): 145-164.
Herberstein ME, Schneider JM, Harmer AMT,etal. Sperm storage and copulation duration in a sexually cannibalistic spider [J].JournalofEthology, 2011, 29 (1): 9-15.
Hollingsworth CS, Capinera JL. Comparison of nymphal development times in diapausing and nondiapausing strains ofMelanoplussanguinipes(F.)(Orthoptera: Acrididae)[J].JournaloftheKansasEntomologicalSociety, 1983, 56 (4): 612-616.
Hoy MA, Knop NF. Development, hatch dates, overwintering success, and spring emergence of a “non-diapause” gypsy moth strain (Lepidoptera: Orgyiidae)in field cages [J].CanadianEntomologist, 1978, 110 (9): 1003-1008.
Ito K. Does artificial selection on diapause incidence cause correlational changes in other life-history traits? Case study in a spider mite population[J].EntomologiaExperimentalisetApplicata, 2009, 130 (3): 266-274.
Kuang XJ, Xu J, Xia QW,etal. Inheritance of the photoperiodic response controlling imaginal summer diapause in the cabbage beetle,Colaphellusbowringi[J].JournalofInsectPhysiology, 2011, 57 (5): 614-619.
Liu XP, He HM, Kuang XJ,etal. Mating behavior of the cabbage beetle,Colaphellusbowringi(Coleoptera: Chrysomelidae)[J].InsectScience, 2010, 17 (1): 61-66.
Liu XP, He HM, Xue FS.The effect of mating frequency and mating pattern on female reproductive fitness in cabbage beetle,Colaphellusbowringi[J].EntomologiaExperimentalisetApplicata, 2013, 146 (3): 379-385.
Ma CH, Ding N, Wang XP,etal. Examination of parental effect on the progeny diapause by reciprocal cross test in the cabbage beetle,Colaphellusbowringi[J].JournalofInsectScience, 2011, 11 (1): 145.
Meng L, Li BP. Mating and oviposition behaviors of exotic leafbeetle,Ophraellacommuna[J].ChineseBulletinofEntomology, 2006, 43 (6): 806-809. [孟玲,李保平.豚草条纹萤叶甲的交配和产卵行为[J].昆虫知识,2006, 43 (6): 806-809]
Riihimaa AJ, Kimura MT. A mutant strain ofChymomyzacostata(Diptera: Drosophilidae)insensitive to diapause-inducing action of photoperiod [J].PhysiologicalEntomology, 1988, 13 (4): 441-445.
Simmons LW. Sperm Competition and its Evolutionary Consequences in the Insects [M]. Princeton: Princeton University Press, 2001.
Socha R, Hodkova M. Selection for non-diapause in the HeteropteranPyrrhocorisApterus[J].Hereditas, 1994, 120 (1): 81-85.
Tan QQ, Zhou RQ, Liu W,etal. Biological characteristics of a non-photoperiodic-diapause strain of the cabbage beetleColaphellusbowringi(Coleoptera: Chrysomelidae)[J].EntomologicalScience, 2016, doi: 10.1111/ens.12226.
Tauber MJ, Tauber CA, Masaki S. Seasonal Adaptations of Insects [M]. New York: Oxford University Press, 1986.
Thornhill R, Alcock J. The Evolution of Insect Mating Systems [M]. Cambridge: Harvard University Press, 1983.
Xue FS, Spieth HR, Li AQ,etal. The role of photoperiod and temperature in determination of summer and winter diapause in the cabbage beetle,Colaphellusbowringi(Coleoptera: Chrysomelidae)[J].JournalofInsectPhysiology, 2002, 48 (3): 279-286.Xue FS, Li AQ, Zhu XF,etal. Diversity in life history of the leaf beetle,ColaphellusbowringiBaly [J].ActaEntomologicaSinica, 2002, 45 (4): 494-498. [薛芳森,李爱青,朱杏芬,等.大猿叶虫生活史的研究[J].昆虫学报,2002, 45 (4): 494-498]
Zhou KN, Zhang JL, Deng L,etal. Effects of copulation duration on fitness consequences of lady beetles,HenosepilachnavigintloctopunctataFabricius [J].BiologicalDisasterScience, 2012, 35 (1): 40-44. [周康念,张爵龙,邓礼,等.交配持续时间对茄28星瓢虫生殖适应性的影响[J].生物灾害科学,2012, 35 (1): 40-44]
Differences inmating behavior between diapause and non-diapause strains of the cabbage beetleColaphellusbowringi(Coleoptera: Chrysomelidae)
ZHOU Rui-Qi, TAN Qian-Qian, WANG Xiao-Ping, LIU Wen*
(Hubei Insect Resources Utilization and Sustainable Pest Management Key Laboratory, College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, China)
The cabbage beetleColaphellusbowringiBaly (Coleoptera: Chrysomelidae)is a serious pest of crucifers and goes into summer or winter diapause as adults. Compared to the diapause strain (DS), the reduced fecundity of non-diapause strain (NDS)is possibly related to mating behavior. The NDS ofC.bowringiis a promising model for investigating the mechanism of diapause inheritance and regulation. In this study, we observed the mating rhythm, mating duration and mating frequency of the DS and NDS inC.bowringiat 25℃ and L ∶D=12 ∶12 h. The results showed that mating rhythm of both strains peaked at 2 h after light period began, and the mating rate of NDS was significant lower than that of DS. The mating durations of the first time of DS and NDS were 122.7±7.4 min and 100.1±10.2 min, respectively, and there is no significant difference. However, the total mating duration of NDS was significant lower than that of DS. The mating frequency of DS was 3.8±0.5 times which was significant higher than that of NDS (2.2 ± 0.3 times). Comparison of the mating behavior between DS and NDS shows that the reduced fecundity of NDS is associated with the mating behavior variation, including decreased mating rate, shortened mating duration and reduced mating frequency. The application of NDS can promote the study of the mechanism of insect diapause inheritance and regulation.
Colaphellusbowringi; non-photoperiodic-diapause strain; mating rhythm; mating duration; mating frequency
国家自然科学基金(31272045, 31572009);中央高校基本科研业务费专项资金(2014BQ012)
周睿琦,女,1991年生,湖北武汉人,农学硕士,研究方向为昆虫生态学,E-mail:richchou19911017@163.com
*通讯作者 Author for correspondence,E-mail: liuwen@mail.hzau.edu.cn
Received:2016-08-16; 接受日期Accepted: 2016-10-22
Q968.1;S433
A
1674-0858(2017)01-0167-05
周睿琦,谈倩倩,王小平,等.大猿叶虫滞育品系和非滞育品系交配行为的比较[J].环境昆虫学报,2017,39(1):167-171.
猜你喜欢
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
园林科技(2020年2期)2020-01-18
石油炼制与化工(2019年9期)2019-09-18
山东林业科技(2019年2期)2019-06-03
云南农业科技(2019年2期)2019-04-04
科技与创新(2016年24期)2017-03-30
中国马铃薯(2015年3期)2015-12-19
第二课堂(课外活动版)(2015年5期)2015-10-21
