
雌性圈养大熊猫在卧龙和雅安繁殖习性的比较研究
2017-02-18 07:05:10黄炎刘洋吴代福张明春黄金燕周世强张贵权李德生张和民
四川动物 2017年1期
黄炎, 刘洋, 吴代福, 张明春, 黄金燕, 周世强, 张贵权, 李德生, 张和民
(中国保护大熊猫研究中心,四川卧龙623006)
雌性圈养大熊猫在卧龙和雅安繁殖习性的比较研究
黄炎, 刘洋, 吴代福, 张明春, 黄金燕, 周世强, 张贵权, 李德生, 张和民*
(中国保护大熊猫研究中心,四川卧龙623006)
本研究通过对比研究卧龙和雅安圈养大熊猫Ailuropodamelanolenca来探讨环境变化对雌性圈养大熊猫繁殖习性的影响。中国保护大熊猫研究中心卧龙核桃坪基地(卧龙基地)和雅安碧峰峡基地(雅安基地)的海拔分别是1 848 m±20 m和1 080 m±42 m,2个基地的海拔相差近800 m。通过对比研究10年间13只雌性圈养大熊猫在不同基地(各5年)的繁殖习性,发现其从卧龙基地到雅安基地以后,每胎产仔数的差异有高度统计学意义(P<0.01),在卧龙基地的平均每胎产仔数为1.66±0.27,雅安基地为1.28±0.24;在卧龙基地的平均发情时间为90.05 d,雅安基地为75.54 d,在雅安基地的平均发情时间比卧龙基地早14.5 d (P<0.05);雌激素峰值在卧龙基地平均为136.1 ng/mg Cr,雅安基地为129.7 ng/mg Cr(P=0.47);在卧龙基地的平均孕娠期为120.3 d,雅安基地为115.3 d,差异无统计学意义(P=0.38)。本研究结果显示,在圈养大熊猫的管理中需要重视环境因子的影响,将来也需要进一步加强光周期和温度对大熊猫发情影响的研究。
大熊猫;发情;海拔;气温
大熊猫Ailuropodamelanoleuca是中国特有的珍稀濒危物种。据全国第四次大熊猫调查结果显示,目前野生大熊猫分布在秦岭、岷山、邛崃山、大相岭、小相岭和凉山六大山系中,受保护力度加大等因素的影响,其种群数量近20年来保持持续缓慢增长态势,但生境破碎化现象仍十分严重,整个种群未摆脱濒危状况,个别局域种群甚至面临灭绝的风险(四川省林业厅,2015)。制约大熊猫种群数量增长的一个重要因素是其繁殖能力较低(胡锦矗,2001)。大熊猫是季节性发情动物,在自然分布区每年仅在春季发情一次(胡锦矗,1990;王雄清,1990;冯文和等,1991),圈养大熊猫也有相同的发情现象(黄炎等,2001;Huangetal.,2011,2012)。从中国保护大熊猫研究中心卧龙核桃坪基地1986—2000年的圈养大熊猫繁育情况来看,圈养大熊猫一般每年3—4月发情,8—9月产仔,每胎1~3仔,多胞胎率为48%。大熊猫幼仔初生体质量和母体妊娠期变化很大,初生幼仔平均体质量143 g,最轻的只有53.5 g,最重的216 g,即使是双胞胎,其体质量也可能有很大差异(黄炎等,2001);大熊猫妊娠期平均137 d,受其受精卵延迟着床(Zhangetal.,2009)的影响,最短的妊娠期只有70 d,最长的却达到了328 d(大熊猫“吉妮”在2006年4月2日人工授精,2007年2月23日产仔),但幼仔发育程度受妊娠时间的影响较小(Zhangetal.,2009)。
动物的繁殖能力与其周边的环境条件关系密切,一些研究表明哺乳动物的胎仔数和鸟类的产卵数受纬度和海拔的影响很大(Conawayetal.,1974;Fleming & Rauscher,1978;Innes,1978;Cockburnetal.,1983;Bilencaetal.,1994;Reinhardt,1997),推测可能是高纬度地区的夏季白昼时间更长,动物用更多时间摄食,为繁殖更多数量的后代提供营养保障(Pianka,1978)。大熊猫的生活环境在发生较大改变以后,其发情时间也会发生相应改变,如借展至日本和歌山的大熊猫“梅梅”于9月中旬发情,并在12月17日产仔(兰景超等,2003);日本神户王子动物园的大熊猫“爽爽”11月发情;美国孟菲斯动物园的大熊猫“丫丫”也曾多次出现秋季发情的情况;泰国清迈(热带)的大熊猫“林惠”2009年2月发情,5月27日产仔,时间比卧龙和雅安都早,但在2013年9月底又发情;大熊猫“网网”从雅安基地转移到澳大利亚阿德莱德市,该地季节与雅安相反,“网网”在次年出现了3次非典型发情,经过了1年的适应期后又调整到每年的9月发情(当地的春季)。
2008年5月12日的汶川大地震破坏了通往卧龙基地的道路,以及基地的通讯和电力设施,迫使中国保护大熊猫研究中心将大熊猫转移至雅安基地和其他动物园。这样的条件客观上为研究环境变化对大熊猫繁殖造成的影响提供了基础,我们也希望通过收集到的地震前后各5年(2004年1月—2008年7月;2008年8月—2013年12月)的数据,揭示环境改变对大熊猫繁殖习性造成影响的规律,为圈养大熊猫管理者制定大熊猫繁殖措施提供理论依据,以促进圈养大熊猫种群的健康发展。
1 研究方法
1.1 实验对象
参与实验的大熊猫均为在卧龙核桃坪基地和雅安碧峰峡基地都参与过繁殖的雌性成年大熊猫,各大熊猫的基本情况如表1所示。
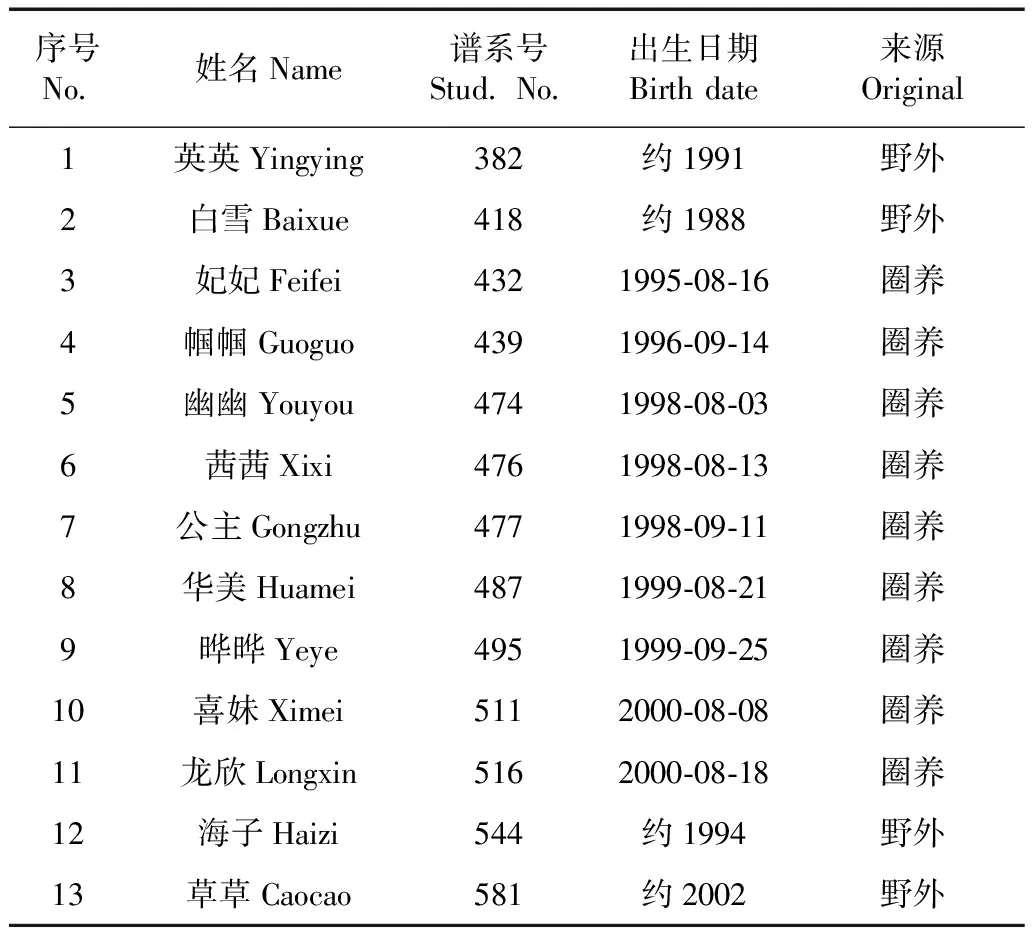
表1 实验对象个体基本信息
1.2 数据收集
收集中国保护大熊猫研究中心卧龙核桃坪基地(103°06′36″E,31°02′24″N,海拔1 848 m)和雅安碧峰峡基地(102°59′24″E,30°04′16″N,海拔1 080 m)各月的气温数据。在保证食物和圈舍环境基本一致的情况下,记录发情配种的第一天与当年1月1日之间的天数来计算发情时间,同时为避免半岁前断奶的雌性大熊猫在产仔后次年再次发情造成的发情时间推迟,只选择受试大熊猫前一年没有带仔的年份来记录发情时间,大熊猫雌激素的检测方法见Huang等(2012)。
1.3 统计分析
将2个基地的月平均气温进行单因素方差分析(One-Way ANOVA)(数据经检验符合正态分布),将每只大熊猫在卧龙基地和雅安基地的平均胎仔数和发情时间进行成对数据的t检验(双尾),且当P<0.05时差异有统计学意义,P<0.01时差异有高度统计学意义。
2 结果
2.1 两地的气温差别
由于海拔较低,雅安基地各月平均气温均高于卧龙基地,一般相差1~6 ℃(图1),但差异无统计学意义(F=3.23,P=0.09)。
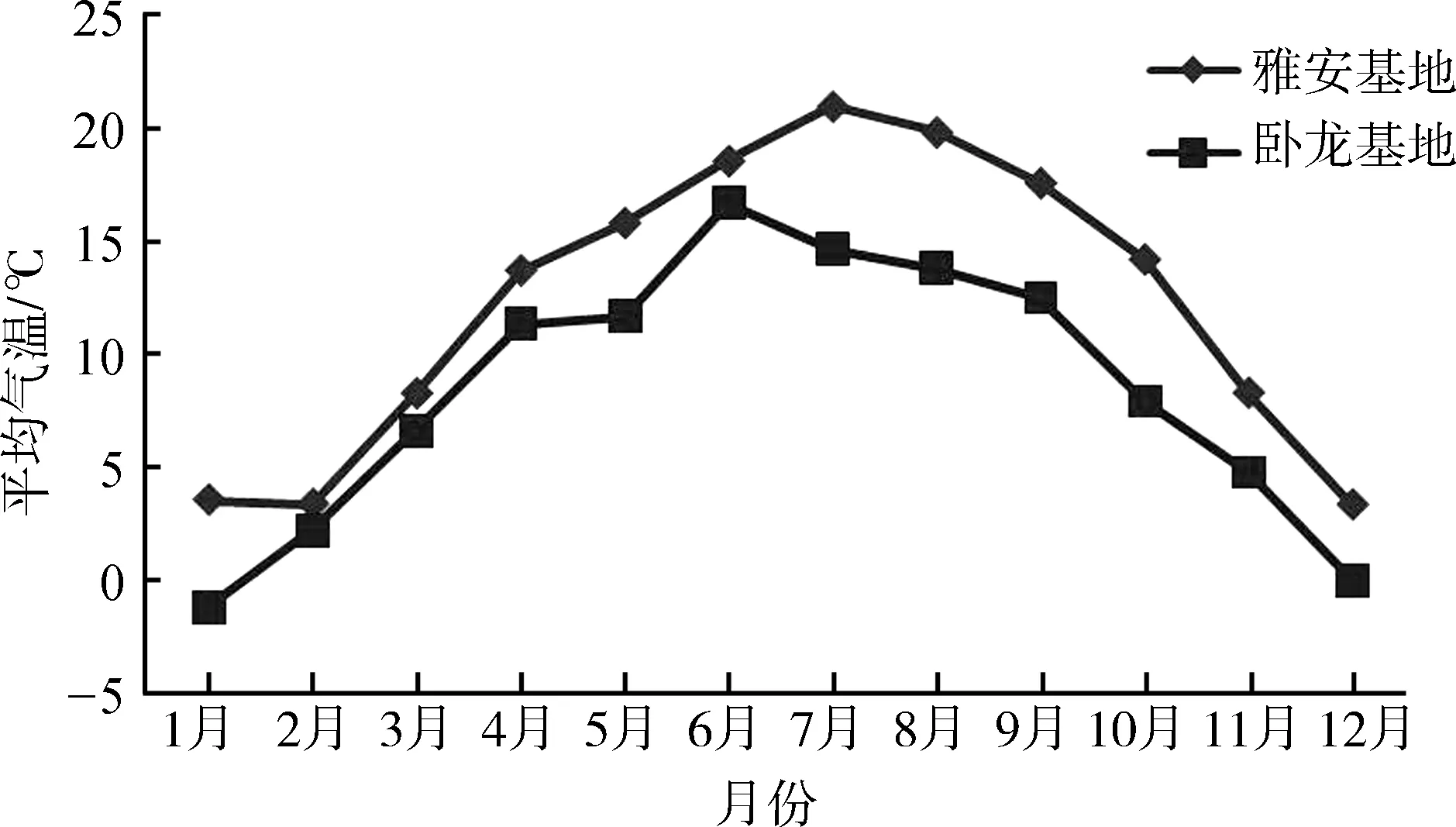
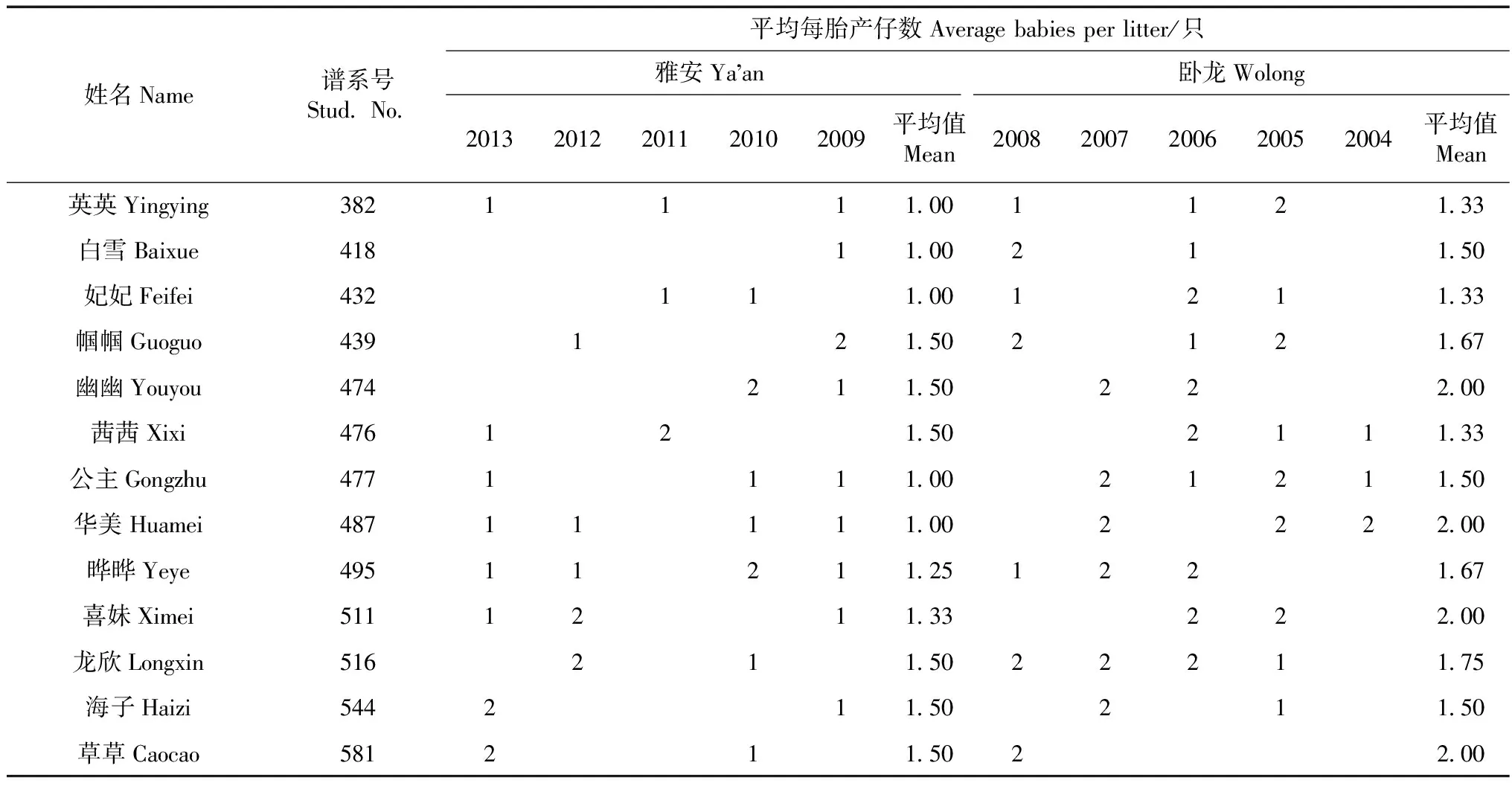
图1 卧龙核桃坪基地和雅安碧峰峡基地的气温变化
2.2 平均每胎产仔数
对13只雌性大熊猫在卧龙基地和雅安基地期间平均每胎产仔数进行对比发现,在卧龙基地的平均每胎产仔数(1.66只±0.27只)高于雅安基地(1.28只±0.24只),两者之间的差异具有高度统计学意义(P<0.01;表2)。
2.3 发情期雌激素峰值
比较这些大熊猫在卧龙基地和雅安基地期间的发情期雌激素峰值,发现在卧龙基地时的雌激素平均峰值(136.1 ng/mg Cr)仅比在雅安基地期间的(129.7 ng/mg Cr)略高,两者之间差异无统计学意义(P=0.47;表3)。
2.4 发情时间
大熊猫“公主”在2003—2007年连续产仔,因此没有统计它在卧龙基地的自然发情时间,其他12只大熊猫在卧龙基地期间的平均发情时间为90.05 d,明显大于它们在雅安基地的平均发情时间(75.54 d,P=0.01;表4)。所以大熊猫在雅安基地的发情时间比在卧龙基地早,事实上大熊猫在雅安基地的发情时间主要集中在1月(3.2%)、2月(9.7%)、3月(67.7%)和4月(19.4%),而在卧龙基地的主要在2月(4%)、3月(48%)和4月(48%)。
2.5 孕娠期
大熊猫在卧龙基地的平均孕娠期为120.3 d,比雅安基地的115.3 d略高,但两者之间差异无统计学意义(P=0.38;表5)。
3 讨论
繁殖力较差一直是制约大熊猫种群数量增长的一个重要原因(张和民等,2003)。大熊猫在现有的

表3 雌性大熊猫在卧龙和雅安发情期雌激素峰值
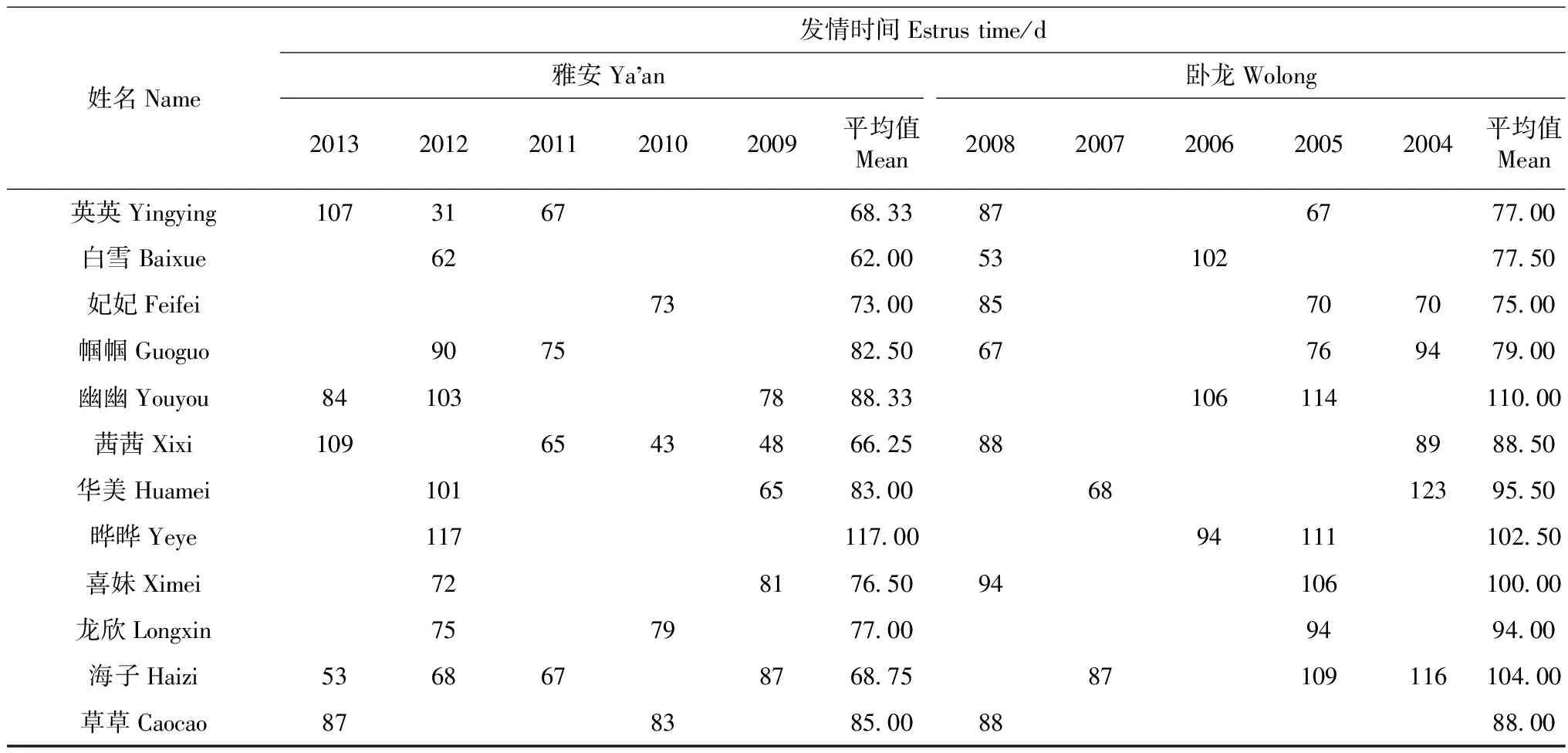
表4 雌性大熊猫在卧龙和雅安的发情时间
分布区内,均生活在海拔较高的高山密林中,环境温度相对较低(胡锦矗,2001)。圈养大熊猫从卧龙迁移至雅安后,不仅纬度减少了1°,海拔也下降了700多米,这些都导致了大熊猫生活的环境温度升高(曹广才等,1990;图1)。而气温会对哺乳动物的繁殖行为产生影响(孙儒泳,2001),因此这些雌性大熊猫在雅安的发情时间比在卧龙期间提前了10多天。大熊猫的季节性发情规律是其长期适应环境的结果,本研究也显示圈养雌性大熊猫的繁殖习性可以根据环境条件的改变而进行适应性调整,如大熊猫“网网”到澳大利亚后就将发情期调整为9月(当地的春季),但在调整过程中会产生非典型发情和发情周期紊乱的情况,同时也影响了大熊猫的总产仔数量。所以管理人员应根据大熊猫繁殖特点制定适宜性措施,减少环境条件改变对大熊猫繁殖能力的影响。由于大熊猫的年龄也会对其繁殖习性产生影响(张和民等,2003),所以我们也将在以后的研究中,着重针对大熊猫年龄对繁殖的影响开展进一步的工作。
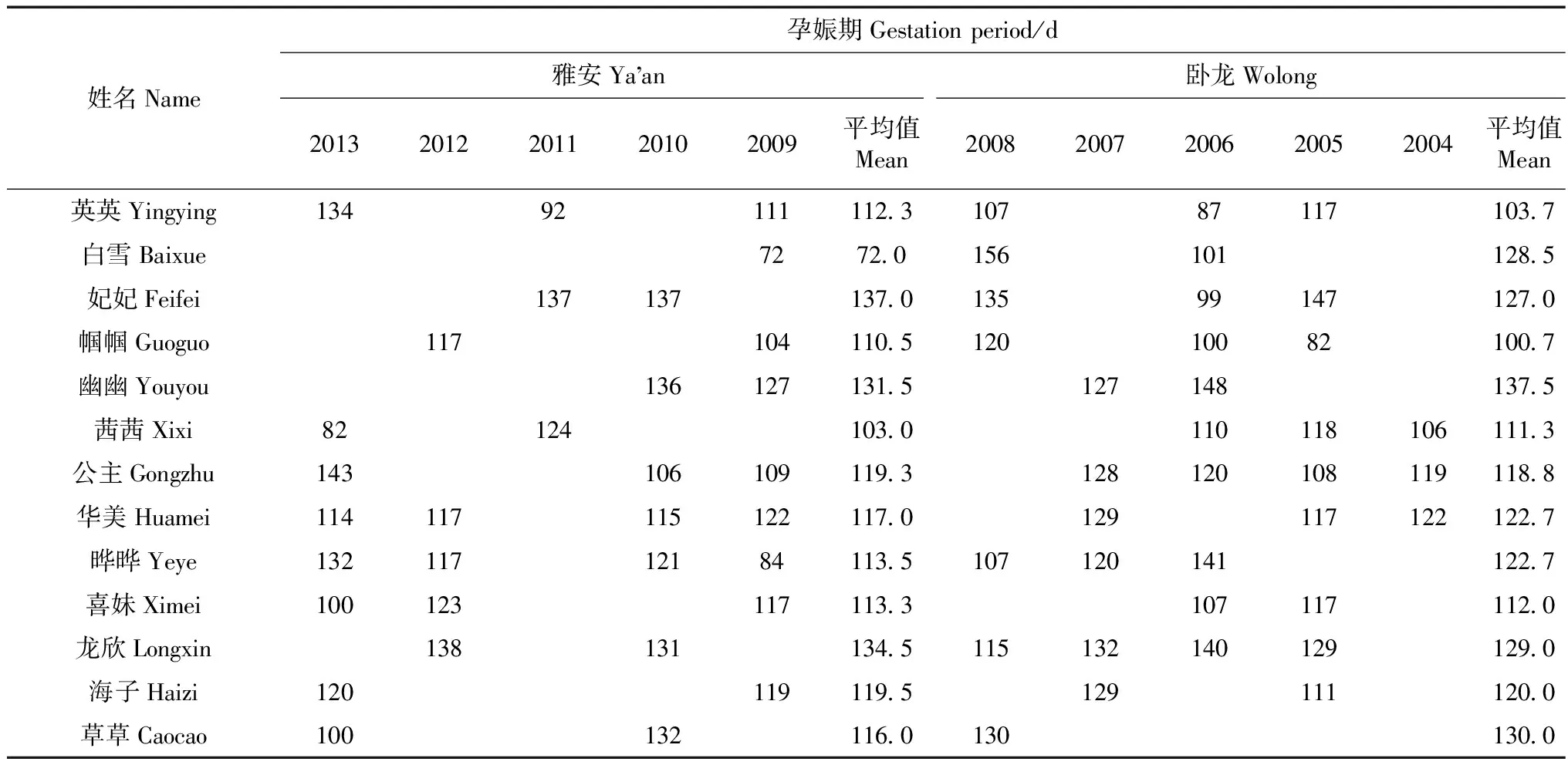
表5 卧龙和雅安圈养大熊猫的孕娠期
光照是影响动物繁殖行为的另一个重要因素(孙儒泳,2001),而不管是卧龙基地还是雅安基地都具有四季分明的特点,但热带地区气候的特点却是全年气温较高,四季界限不明显,日温度变化大于年温度变化,光照强度在一年四季的变化也不大。因此,一旦大熊猫的生活环境变成了热带地区,如泰国、新加坡和马来西亚等,就会影响大熊猫的生理周期。在这些地区的大熊猫场馆中添加设施,模拟卧龙基地的环境温度和光周期可能会减少环境变化对这些大熊猫的影响。当然,进一步加强光周期和温度对大熊猫发情行为影响的研究,或从分子水平弄清雌性大熊猫发情的机理,将更有助于提高其繁殖能力。
致谢:感谢蔡水花同志协助进行数据整理工作。
曹广才, 吴东兵, 金陵, 等. 1990. 纬度对内陆低海拔地区一些冬型小麦品种生育天数的影响[J]. 安徽农业科学, 44(2): 123-127.
冯文和, 叶志勇, 何光昕. 1991. 大熊猫生育能力的研究[J]. 四川大学学报(自然科学版), 28: 15-21.
胡锦矗. 1990. 大熊猫生物学研究与进展[M]. 成都: 四川科学技术出版社: 19-23.
胡锦矗. 2001. 大熊猫研究[M]. 上海: 上海科技教育出版社.
黄炎, 张贵权, 张和民. 2001. 大熊猫的繁殖研究[J]. 四川师范学院学报(自然科学版), 22(3): 203-208.
蓝景超, 吕文其, 张志和, 等. 2003. 首例大熊猫秋季发情、配种、产仔[J]. 四川动物, 22(1): 9-11.
四川省林业厅. 2015. 四川的大熊猫——四川省第四次大熊猫调查报告[C]. 成都: 四川科学技术出版社.
孙儒泳. 2001. 动物生态学原理[M]. 北京: 北京师范大学出版社.
王雄清. 1990. 大熊猫繁殖研究进展[J]. 四川师范学院学报(自然科学版), 11(2): 118-123.
张和民, 王鹏彦, 张贵权, 等. 2003. 大熊猫繁殖研究[M]. 北京: 中国林业出版社.
Bilenca DN, Zuleta GA, Kravetz FO. 1994. Latitudinal variationin litter size and length of the breeding season in populations of pampas mice,Akodonazarae[J]. Mammalia, 58: 563-568.
Cockburn A, Lee AK, Martin RW. 1983. Macrogeographic variation in litter size inAntechinus(Marsupialia, Dasyuridae)[J]. Evolution, 37: 86-95.
Conaway CH, Sadler KC, Hazelwood DH. 1974. Geographic variation in litter size and onset of breeding in cottontails[J]. Journal of Wildlife Manage, 38: 473-481.
Fleming TH, Rauscher RJ. 1978. On the evolution of litter size inPeromyscusleucopus[J]. Evolution, 32: 45-55.
Huang Y, Li DS, Zhou YM,etal. 2011. Factors affecting the outcome of artificial insemination using cryopreserved spermatozoa in the giant panda (Ailuropodamelanoleuca)[J]. Zoo Biology, 30: 1-13.
Huang Y, Zhang HM, Li DS,etal. 2012. Relationship of the estrogen surge and multiple mates to cub paternity in the giant panda (Ailuropodamelanoleuca): implications for optimal timing of copulation or artificial insemination[J]. Biology of Reproduction, 87(5): 517-527.
Innes DGL. 1978. A reexamination of litter size in some north American microtines [J]. Canadian Journal of Zoology, 56: 1488-1496.
Pianka ER. 1978. Evolutionary ecology, 2nd ed[M]. New York: Harper & Row.
Reinhardt K. 1997. Breeding success of southern hemisphere skuasCatharactaspp.: the influence of latitude[J]. Ardea, 85: 73-82.
Zhang HM, Li DS, Wang CD,etal. 2009. Delayed implantation in giant pandas: the first comprehensive empirical evidence[J].Reproduction, 138: 979-986.
Comparison of Reproductive Characteristics of Captive FemaleAiluropodamelanoleucain Ya’an and Wolong
HUANG Yan, LIU Yang, WU Daifu, ZHANG Mingchun, HUANG Jinyan, ZHOU Shiqiang,ZHANG Guiquan, LI Desheng, ZHANG Hemin*
(China Conservation and Research Center for the Giant Panda, Wolong, Sichuan Province 623006, China)
This study analyzed the relationship between captive female giant pandas’ reproduction characteristics and environmental factors. China Conservation and Research Center for the Giant Panda has 2 bases, Wolong Hetaoping Base and Ya’an Bifengxia Base, which are located at 1 848 m±20 m and 1 080 m ±42 m in elevation, respectively. In the past10 years, 13 captive female giant pandas had been raised in both bases (5 years in each base). Comparing their reproductive characteristics in this period, statistics showed that there was a significant difference to the litter size and timing of estrus between 2 bases, but had no difference to peak estrogen and pregnancy. In detail, the average litter size was 1.66±0.27 in Wolong Hetaoping Base, and this was extreme significantly different from that of Ya’an Bifengxia Base (1.28±0.24,P<0.01). Additionally, beginning on every January 1, the captive female giant pandas in Ya’an Bifengxia Base experienced a 75.54 days’ rutting period on average, and this was about 15 days earlier than that in Wolong Hetaoping Base (P<0.05). Furthermore, the peak estrogen value of the giant panda in Wolong Hetaoping Base was 136.1 ng/mg Cr and 129.7 ng/mg Cr in Ya’an Bifengxia Base (P=0.47). Besides, the pregnancy period of captive female giant pandas in Wolong Hetaoping Base was 120.3 days on average, and about 115.3 days in Ya’an Bifengxia Base (P=0.38). These findings indicated that environmental factors were important and should be taken into consideration in captive giant panda management. Investigations on the relationship between giant pandas’ oestrus and photoperiod illumination and temperature could be carried out in further studies, as well as analysis of the mechanisms driving female giant pandas’ oestrus at the molecular level.
Ailuropodamelanoleuca; oestrus; elevation; temperature
2016-06-20 接受日期:2016-11-16
香港海洋公园保育基金项目(GP02_1415); 国家林业局国际合作项目(SD0628,KB1401)
黄炎(1964—), 男, 教授, 研究方向:繁殖生理和保护生物学,E-mail: pandayard@hotmail.com
*通信作者Corresponding author, E-mail:wolong_zhm@126.com
10.11984/j.issn.1000-7083.201600165
Q959.8
A
1000-7083(2017)01-0014-05
猜你喜欢
今日农业(2021年11期)2021-08-13 08:53:40
中华环境(2021年9期)2021-04-09 05:26:37
今日农业(2020年14期)2020-12-14 19:47:34
下一代英才(酷炫少年)(2017年10期)2017-11-22 02:16:08
快乐语文(2016年10期)2016-11-07 09:44:58
知识经济·中国直销(2016年5期)2016-11-07 09:35:19
小说月刊(2015年11期)2015-04-23 08:47:34
电机与控制应用(2015年3期)2015-03-01 03:50:03
航天器工程(2014年5期)2014-03-11 16:36:02
河南科技(2014年23期)2014-02-27 14:19:07
