
气候变化对阔叶红松林潜在地理分布区的影响
2017-02-08 06:13:58马芳芳周旺明于大炮代力民
生态学报 2017年2期
贾 翔,马芳芳,周旺明,周 莉,于大炮, 秦 静, 代力民,*
1 中国科学院沈阳应用生态研究所,森林生态与管理重点实验室,沈阳 110016 2 中国科学院大学,北京 100049 3 辽宁省林业种苗管理总站,沈阳 110036
气候变化对阔叶红松林潜在地理分布区的影响
贾 翔1,2,马芳芳1,2,周旺明1,周 莉1,于大炮1, 秦 静3, 代力民1,*
1 中国科学院沈阳应用生态研究所,森林生态与管理重点实验室,沈阳 110016 2 中国科学院大学,北京 100049 3 辽宁省林业种苗管理总站,沈阳 110036
物种地理分布主要取决于它对气候、地形等环境因子的适应性。基于22个环境因子和阔叶红松林的4类主要建群树种——红松、紫椴、水曲柳和蒙古栎的地理分布数据,采用最大熵模型模拟了阔叶红松林的潜在分布区域,并分析决定阔叶红松林地理分布的主要气候和地形因子,最后利用政府间气候变化专门委员会(IPCC)发布的3种排放场景(SRES-A2、SRES-A1B、SRES-B1)下2020、2050、2080年的气候数据预测阔叶红松林的未来潜在分布区。结果表明:各树种的受试者工作特征曲线下面积(AUC值)都大于0.8,说明模型有很好的预测能力;影响阔叶红松林分布的主导环境因子是年降雨量、季节性降雨量、海拔、年平均温度、最湿季度的平均温度。在基准气候条件下,阔叶红松林的高度适宜分布区主要分布在长白山和小兴安岭地区,占研究区总面积的11.69%,低度适宜区面积、不适宜区面积分别占研究区总面积的23%和65.31%。模型预测结果显示,未来在A2、A1B和B1气候情景下,阔叶红松林高度适宜区的南界与北界都向北移动,其面积有缩减的趋势,而低度适宜区的面积有增加的趋势。
阔叶红松林;气候变化;最大熵模型;潜在地理分布;气候变化情景
分析物种-环境之间的关系已经成为生态学、生物地理学中的一个焦点问题[1],物种在各种各样的生物因子和非生物因子的共同作用下,都有其暂时的地理分布范围。物种地理分布主要取决于它对气候、地形等环境因子的适应性,特别是森林群落优势物种的地理分布与气候有着密切的关系,其地理分布受到气候变化的深刻影响[2]。而很多大气环流模型预测研究表明,到21世纪末,地表平均温度将升高1.1—6.4℃[3],因此,全球气候变暖将会对物种地理分布产生极大的影响。
关于物种地理分布的研究方法已有很多,其中比较热门的研究方法是物种分布模型(Species Distribution Models, SDMs)。物种分布模型主要是利用物种的分布数据与环境数据,依据特定的算法估计物种的生态位,并投影到景观中,以概率的形式反映物种对生境的偏好程度[4]。在所有物种分布模型中,最大熵模型(Maxent)相比而言预测效果最好[5-6]。
阔叶红松林树种资源丰富,红松(Pinuskoraiensis)为主要成林树种,并与其他针阔叶树种如鱼鳞云衫(Piceajezoensis)、臭冷杉(Abiesnephrolepis)、紫椴(Tiliaamurensis)、水曲柳(Fraxinusmandshurica)和蒙古栎(Quercusmongolica)等形成针阔混交林[7],是东北东部山区典型的地带性植被,也是第四纪冰川期后保存完好的原始裸子植物群落[8],其分布中心在东北长白山、张广才岭、小兴安岭和完达山山地。由于历史上的过度采伐,阔叶红松林遭到严重破坏,其分布面积急剧萎缩。为了实现阔叶红松林的生态恢复及可持续经营,研究阔叶红松林在未来气候变化情况下的潜在分布区显得至关重要。目前,关于阔叶红松林地理分布的研究还局限于单一的树种[9-10],而单一树种的分布区域不能直接代替阔叶红松林群落的分布区域,并且这些研究仅仅探讨了为数不多的环境因子,如温度累积指标生长度·日(GDD)和水分指标可能蒸散率(PER)对其产生的影响[11]。本文以阔叶红松林的4类主要建群树种——红松、紫椴、水曲柳和蒙古栎为研究对象,通过最大熵模型分析决定阔叶红松林地理分布的主要气候和地形因子,并分别模拟4类树种的潜在分布区,根据它们的共同分布区域来决定阔叶红松林的分布区,最后利用气候变化下的未来环境数据预测阔叶红松林的未来潜在分布区,为阔叶红松林的生态恢复和经营管理提供理论依据。
1 材料与方法
1.1 研究区域
阔叶红松林主要分布于我国东北地区,其地处亚欧大陆东缘,地理坐标38°43′—53°23′N,118°50′—135°05′E,包括辽宁、吉林、黑龙江3省以及内蒙古自治区东部的呼伦贝尔市、兴安盟、通辽市、赤峰市,南北长1600多km,东西宽1400多km[12],南临黄海和渤海,东北与俄罗斯接壤,东南以鸭绿江为界与朝鲜民主主义人民共和国隔江相望,西部与内蒙古自治区接壤[13]。
1.2 数据采集
1.2.1 树种地理分布数据
通过中国数字植物标本馆(http://www.cvh.org.cn/)、教学标本资源共享平台(http://mnh.scu.edu.cn/)、中国自然保护区标本资源共享平台(http://www.papc.cn/)等数据库获得标本采集地信息,同时查阅《中国植物志》和东三省植物志以及相关研究文献确定其现在分布点。去除模糊记录的分布点信息,对具有详细信息的分布点,利用百度拾取坐标系统(http://api.map.baidu.com/lbsapi/getpoint/index.html)确定其分布点的经纬度。另外,为提高预测的可靠性,剔除引种栽培的树种分布点。
1.2.2 气候因子
气候基准数据来源于WorldClim数据库(http://www.worldclim.org),该数据库收集了1950—2000年全球各地气象站的每月气象数据,采用插值法生成全球气候栅格数据,将2000年作为基准年。本文基于ArcGIS 10.0平台,提取了东北地区的19个气候因子(Bio1—Bio19)(表1),空间分辨率为30″(约1km2)。徐影等(2002)对国际上较流行的5种大气环流模式(HadCM、GFDL、ECHAM、CSIRO以及CGCM)进行比较研究,研究结果表明ECHAM4和HadCM2两个模式对东亚和中国地区的气候模拟效果最好[14],本文采用的未来气候数据由大气环流模型HadCM模拟,版本为HadCM3。研究涉及的3种排放场景来自《IPCC排放情景特别报告》,分别代表了碳排放高速增长(SRES-A2)、中速增长(SRES-A1B)和低速增长(SRES-B1)。其中A2情景是描述了一个非均衡的世界:各地域间生产力方式的趋同异常缓慢,由此导致人口持续增长,经济发展主要面向区域,人均经济增长和技术变化是不连续的,并低于其他情景的发展速度;A1情景描述的世界是:经济增长迅速、全球人口峰值将出现在本世纪中叶,新的更高效的技术被迅速引进,A1情景进一步划分为3组情景,而A1B是能源使用较为平衡的一种类型;B1情景是描述了一个趋同的世界:全球人口数量与A1相同,但经济结构向服务和信息经济方向迅速调整,伴之以材料密集程度的下降,以及清洁和资源高效技术的引进[15]。以上3种排放场景、3个未来时段(2020、2050、2080年)的未来气候数据由国际热带农业中心(CIAT, http://www.ccafs-climate.org)提供,空间分辨率为30″(1km2)。
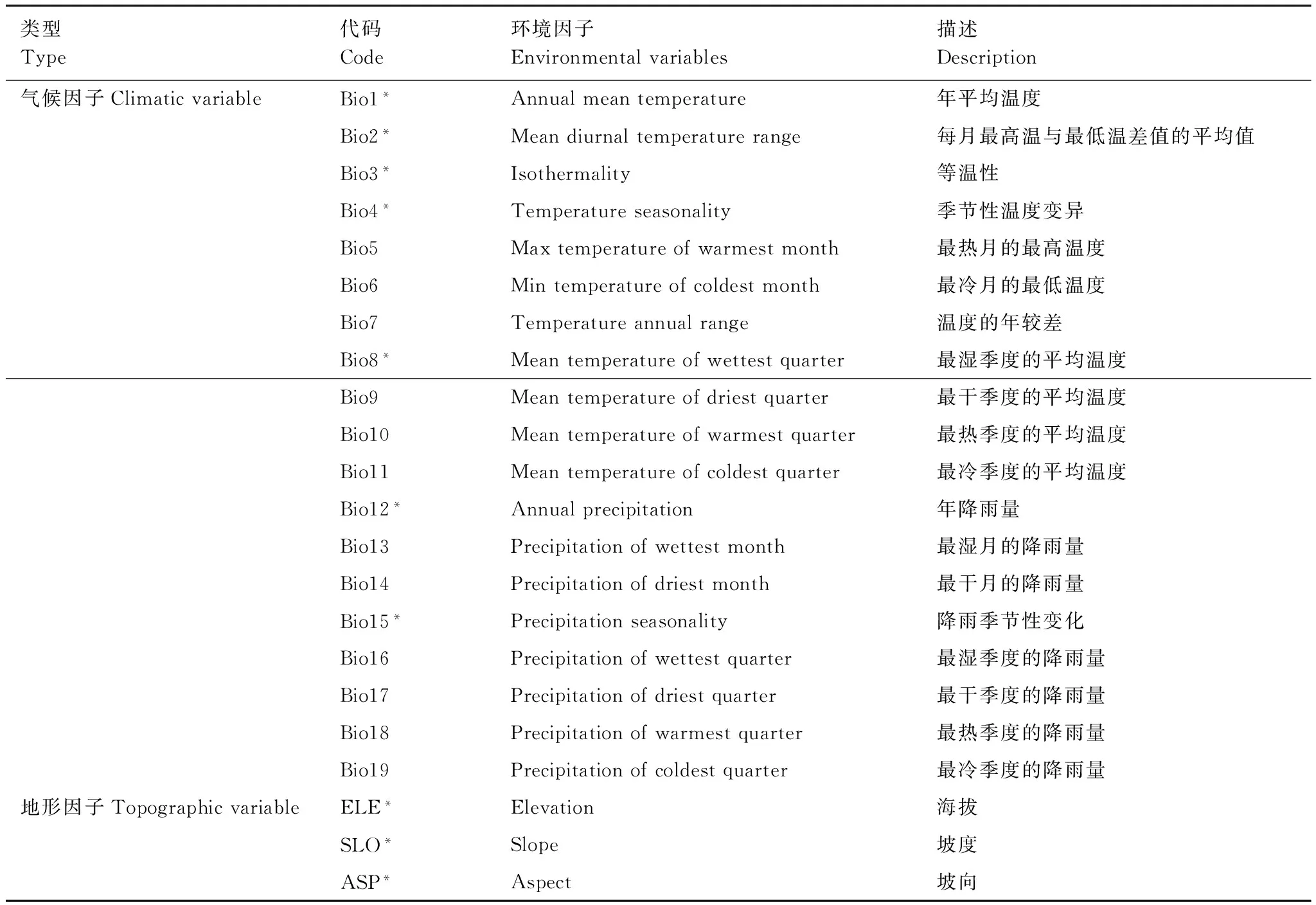
表1 研究采用的环境因子
* 经筛选后输入模型的因子
1.2.3 地形因子和底图
地形数据来源于中国科学院计算机网络信息中心国际科学数据镜像网站(http://www.gscloud.cn)。下载分辨率为90m的数字高程地图(DEM),从中提取海拔、坡度、坡向(表1),而后进行重采样生成分辨率为30″的栅格数据图层。中国地图和中国省级行政区划图来自国家基础地理信息系统网站(http://nfgis.nsdi.gov.cn/)。
1.3 数据分析
1.3.1 树种地理分布区
将树种分布点整理到EXCEL中,共获取红松分布点167个,蒙古栎分布点210个,紫椴分布点177个,水曲柳分布点106个(图1)。陈新美等对Maxent模型预测物种分布所需样本量的研究表明,样本量大小对Maxent模型预测物种空间分布的精度影响不大[16],所以本文数据个数可以进行模拟。
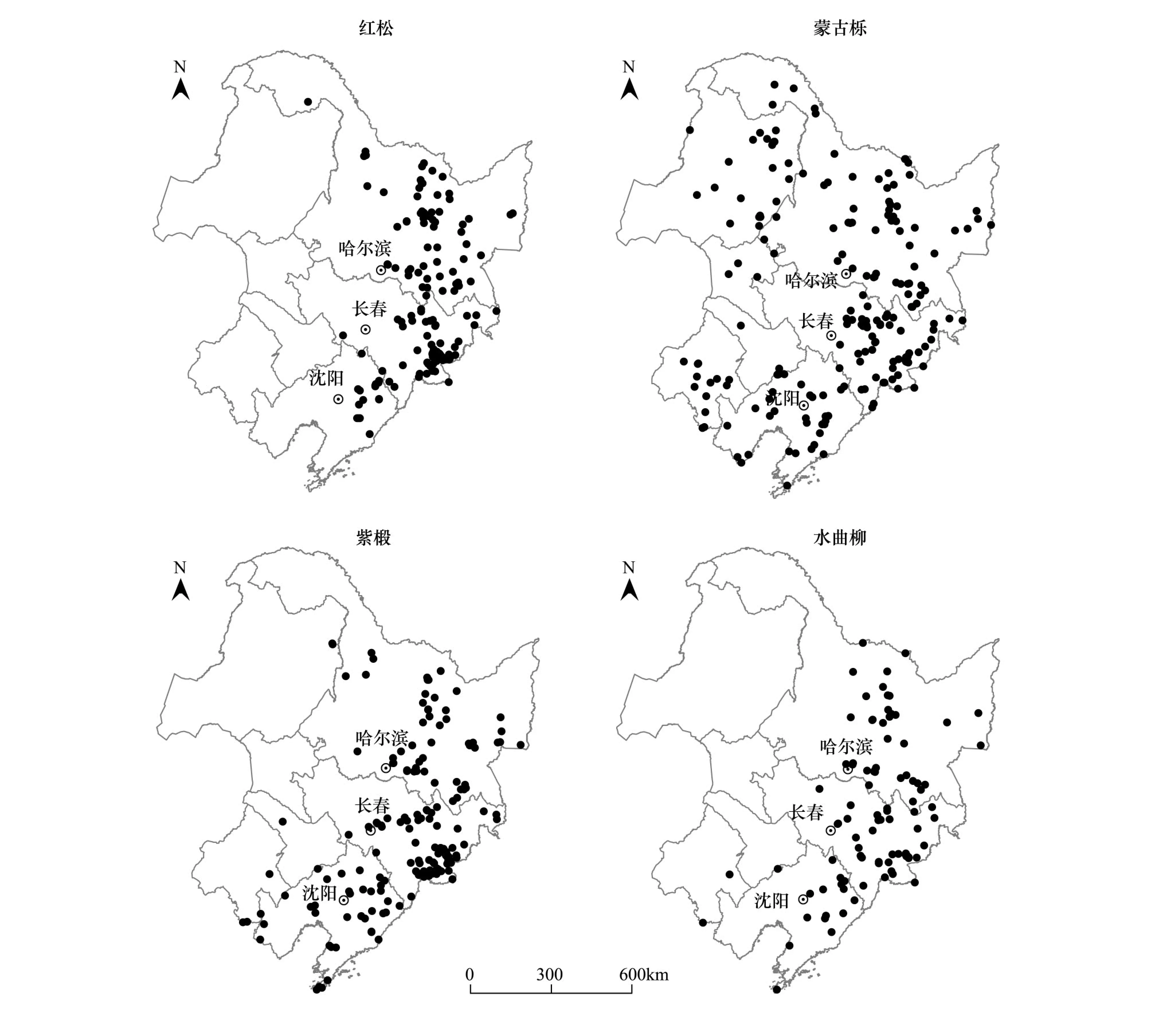
图1 红松、蒙古栎、紫椴和水曲柳分布点的地理位置Fig.1 Geographic locations of Pinus koraiensis, Quercus mongolica, Tilia amurensis and Fraxinus mandshurica
1.3.2 环境因子的筛选
由于各环境因子之间具有一定的相关性,如果直接应用于模型,可能会产生过度拟合现象,所以对环境因子进行相关性分析后才可以应用于物种分布模型[17]。参照Yang等筛选环境因子的方法[1],对22个环境因子图层进行多重共线性分析(SPSS 19.0)来检验图层之间的相关性,若两个环境因子之间的Pearson指数|r|≥0.8,那么只能有一个因子选入模型,最后筛选得到10个环境因子,包括7个气候因子(Bio1、Bio 2、Bio 3、Bio 4、Bio 8、Bio 12和Bio 15)和3个地形因子(海拔、坡度、坡向)(表1)。
1.3.3 物种分布模型
Maxent模型是物种分布模型中表现较好的一种模型,它基于可以免费使用的Maxent软件V3.3.3.k(http://www.cs.princeton.edu/—schapire/maxent/)。Maxent软件运行输入数据包括研究区域的一组环境图层和一个物种在该区域的分布数据。参照Moreno等研究方法[18],随机选取25%的分布点作为测试集(testing data),重复运算10次进行建模,其他参数为默认值。采用受试者工作特征曲线(receiver operating characteristic curve, ROC)下面积(Area under curve, AUC)来评价模型的拟合程度。选择AUC值最大的一次运算结果进行物种地理分布的预测,AUC值越大表明拟合越好,目前以AUC值0.5—0.6为较差,0.6—0.7为一般,0.7—0.8为较准确,0.8—0.9为很准确,0.9—1为极准确。在环境参数设置中开启刀切法(Jackknife)来评价各个环境变量的权重,结合每个环境因子的贡献百分比来判定主导因子。模型的输出格式为ASCLL格式文件,利用ASCII to Raster转换为栅格数据,按栅格数值的大小将研究区域分为3个适宜等级,具体参考政府间气候变化专门委员会(IPCC)关于评估可能性的划分标准[19]:P<0.05为不适宜区;0.05≤P<0.33为低度适宜区;P≥0.33为高度适宜区。本文将4类树种共同具有的高度适宜区作为阔叶红松林的高度适宜区,用4类树种的不适宜区求并集作为阔叶红松林的不适宜区,其余作为低度适宜区。
2 结果与分析
2.1 模型模拟的准确性评价
目前,ROC曲线分析法已经广泛应用于准确评价物种分布模型的预测能力。本文对4类树种的分布进行模拟,各树种的AUC值如下(表2)。其中红松表现最好,其次是水曲柳、紫椴、蒙古栎。当Test AUC>0.8时,Maxent模型具有很好的预测能力,模拟结果保持稳定,可以预测物种分布区域[20]。
2.2 影响阔叶红松林分布的主导环境因子
通过对4类树种的环境因子贡献百分比叠加分析(图2),可知影响阔叶红松林分布的主导环境因子是年降雨量(Bio12)、季节性降雨量(Bio15)、海拔(ELE)、年平均温度(Bio1)和最湿季度的平均温度(Bio8),对于红松、蒙古栎、水曲柳,它们的累计贡献率都大于85%,对于紫椴,它们的累计贡献率大于80%,其中年降雨量为最主要的因子。基于Jackknife模块的输出结果也能很好地反映这些因子对各树种分布的重要性。

表2 各树种模型模拟的AUC值
2.3 阔叶红松林适宜分布区的空间变化趋势分析
2.3.1 阔叶红松林潜在分布区的模拟
运行Maxent模型后输出4类树种的潜在分布区,利用Reclassy工具划分等级,然后叠加得到阔叶红松林分布图(图3)。阔叶红松林的高度适宜分布区主要分布在长白山和小兴安岭地区,东起吉林珲春市、黑龙江东宁县,北界位于黑龙江伊春市、鹤岗市、萝北县,向南经小兴安岭、张广才岭、大青山、吉林哈达岭、辽宁本溪,到辽宁岫岩满族自治县、凤城市北部、桓仁满族自治县。阔叶红松林的低度适宜区主要分布在东北地区的东南面,北界位于黑龙江黑河市,向南延伸至辽宁普兰店市、瓦房店市。阔叶红松林的高度适宜区面积占研究区总面积的11.69%,低度适宜区面积占研究区总面积的23%,不适宜区面积占研究区总面积的65.31%(表3)。
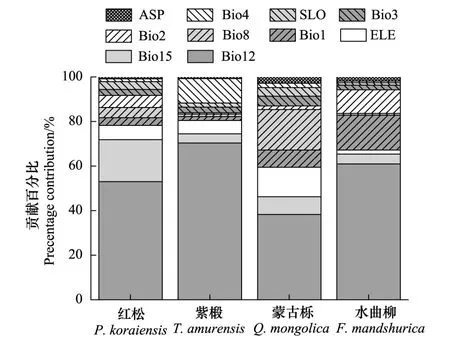
图2 各树种的环境因子贡献百分比 Fig.2 Percentage contribution of environmental variables to the species distribution models
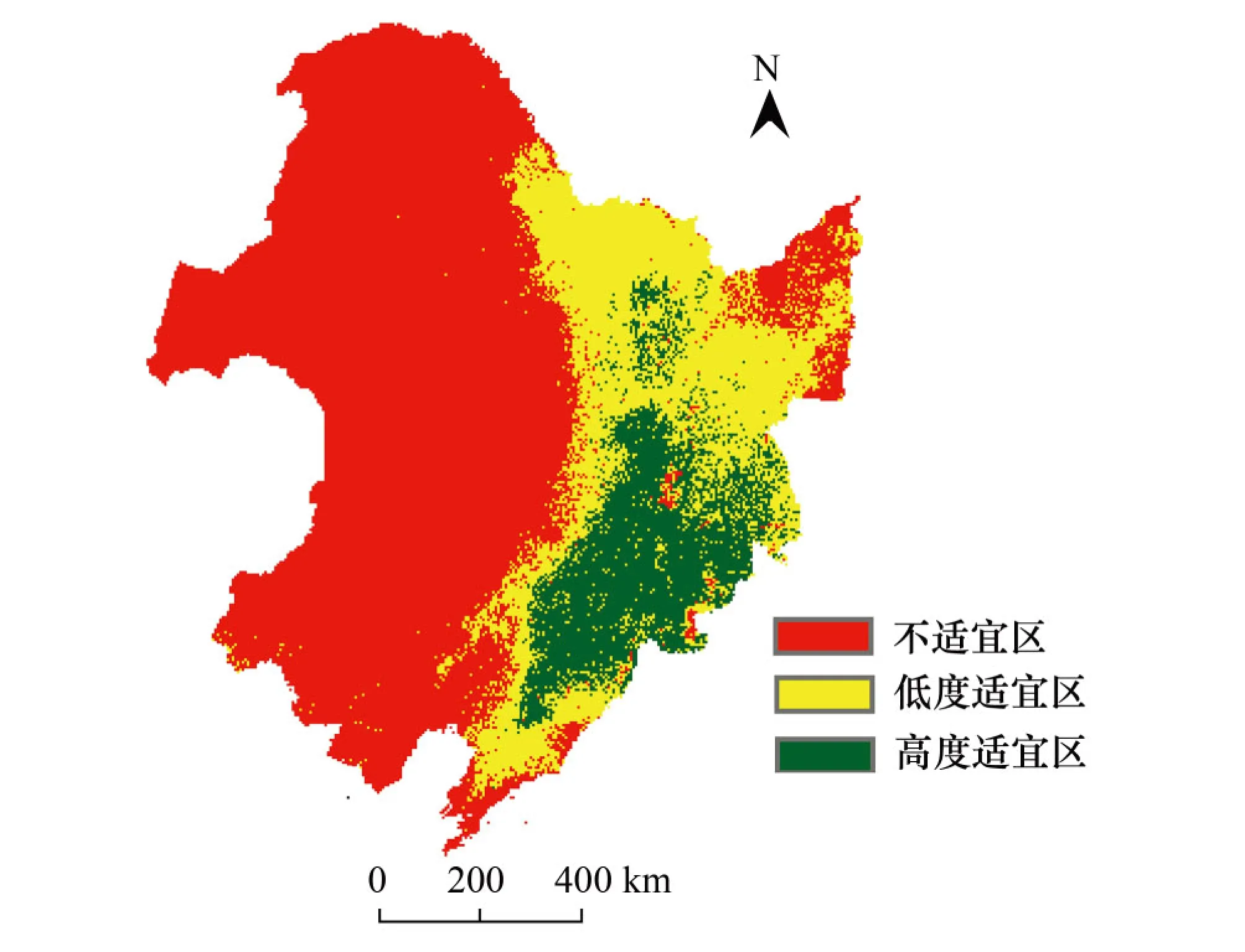
图3 阔叶红松林的潜在分布图Fig.3 Potential distribution of broadleaved Pinus koraiensis forest
2.3.2 未来阔叶红松林潜在分布区的预测
由基准环境数据与物种分布数据建立的模型,预测效果良好。在此基础上,将物种和环境因子之间的联系投影到未来气候情景中用于预测4类树种的未来分布,得到阔叶红松林在3种气候情景下,2020、2050、2080年的预测分布图(图4)。
在A2、A1B、B1气候情景下,阔叶红松林高度适宜区将主要集中分布于长白山自然保护区和小兴安岭部分地区。其分布南界与北界都向北移动,在A2、A1B情景下的2080年尤为明显。
在A2气候情景下,阔叶红松林的高度适宜区面积波动不定,由原来的11.69%骤减到2020年的4.77%,到2050年增加到5.66%,而后2080年又减少为0.60%;阔叶红松林的低度适宜区面积从2000年的23%呈现增加的趋势,到2080年为37.18%(图4,表3)。
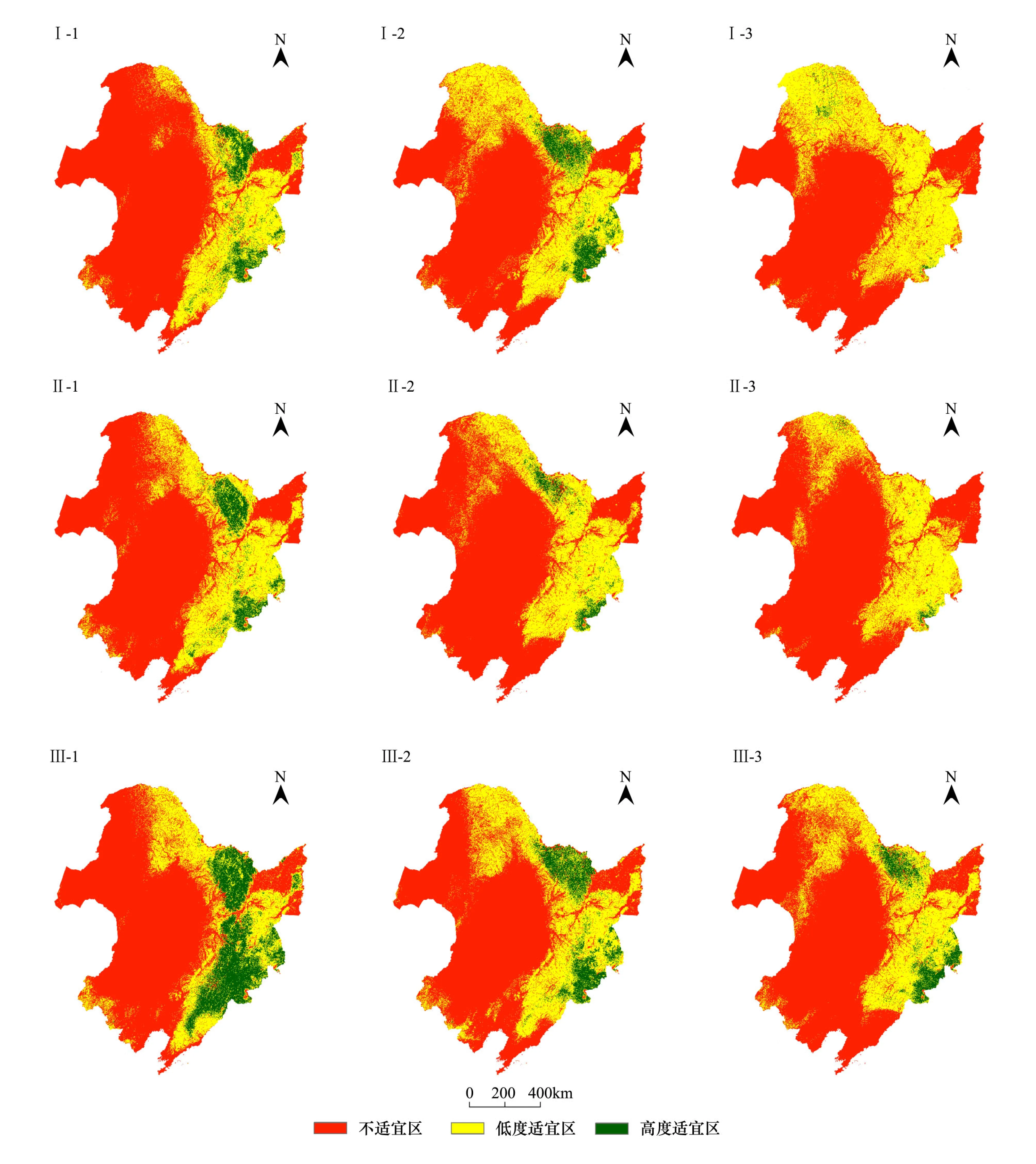
图4 不同气候情景下,阔叶红松林的潜在分布预测图Fig.4 Predicted potential distribution of broadleaved Pinus koraiensis forest under different climate change scenariosⅠ: A2气候情景;Ⅱ: A1B气候情景;Ⅲ: B1气候情景;1: 21世纪20年代;2: 21世纪50年代;3: 21世纪80年代。如:Ⅱ—2,指A1B气候情景下21世纪50年代
在A1B气候情景下,阔叶红松林的高度适宜区面积表现为减少趋势,由基准年的11.69%骤减到2020年的4.71%,到2050年减小到2.17%,而2080年缩少至0.46%;阔叶红松林的低度适宜区面积从2000年的23%增加到2020年的28.01%,至此接近稳定。
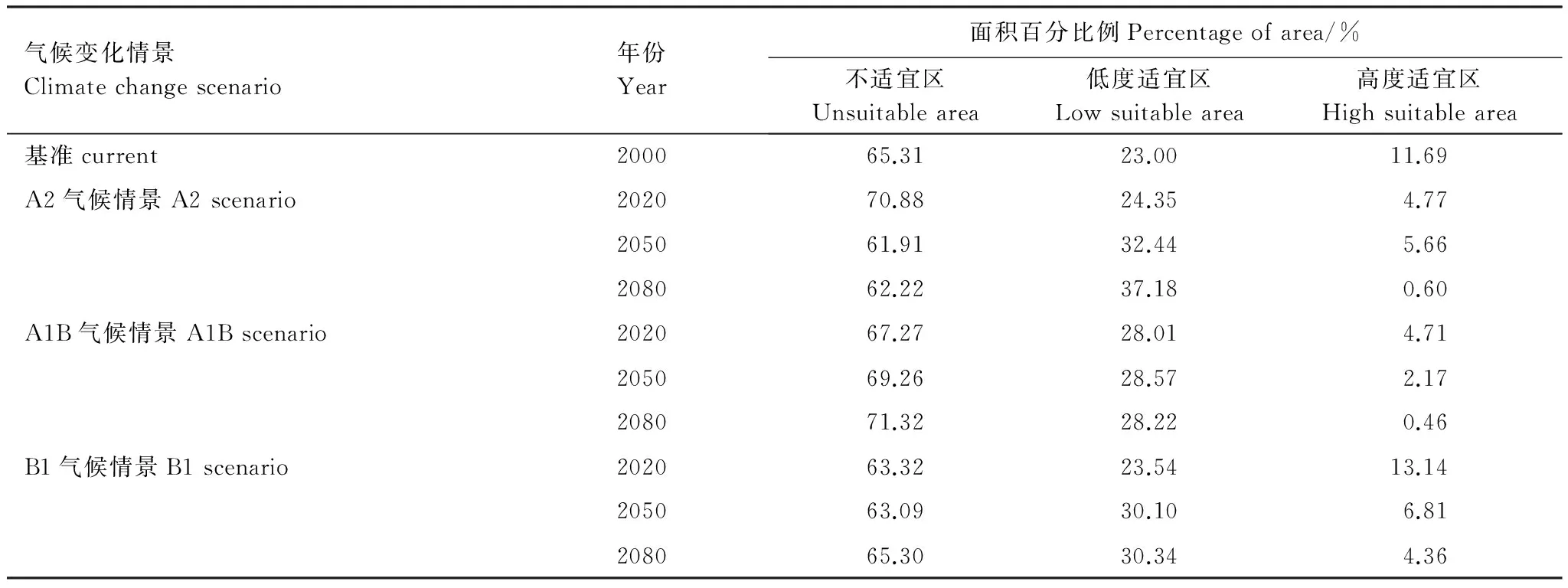
表3 阔叶红松林的不同适宜等级面积百分比
而在B1气候情景下,阔叶红松林的高度适宜区面积开始稍有增加而后减少,由11.69%增长至2020年的13.14%,但在2050和2080年其面积分别降为6.81%和4.36%;阔叶红松林的低度适宜区面积从2000年(23%)到2020年(23.54%)几乎没有变化,但是到2050年增加为30.10%,而后保持稳定。
3 讨论
已有研究表明,限制植物地理分布的主要因子是植物的耐寒性、完成生活史所需的热量供应和可利用水[21]。殷晓洁等研究表明影响蒙古栎的最主要因子是年均降水量[10],本文得到与其相同的结论。孙晓红研究表明影响红松地理分布的最主要气候因子是最冷月最热月平均温度差,其次是年降水量[9],与本研究结果有一定的差异。其主要原因是不同的研究所采用的环境因子不同,本文增加了地形因子作为环境因子,此外,由于环境因子的筛选过程不同,也会将不同的环境因子选入模型。
对东北地区气温和降水变化的研究表明,在A2、A1B、B1三种排放情景下未来100年的气温和降水总体趋势均呈逐渐增加的趋势。此外,在A2情景下气温和降水的增幅最高,其次是A1B情景,变化幅度最小的是B1排放情景[22]。有研究表明红松地理分布区面积与降水量呈正相关关系,与年平均温度呈负相关关系[9],而全球气候变化将导致东北地区温度明显升高,温度带北移,降水量有所增加,但抵消不了由温度升高造成的蒸散量的增加,因此全球气候变化将使东北东部山区面临暖干的气候变化趋势,所以实际上气候的暖干化促使阔叶红松林分布区面积有减少的趋势,生态适宜性显著下降[11]。程肖侠等研究表明,在气候变暖降水增加的背景下,红松有成为大兴安岭地区森林主要树种的趋势[23],而本文中在3种气候情景下,低度适宜区都有显著的北移,到2020年,其分布区域扩展到大兴安岭地区,在A2、A1B气候情景下到2080年,大兴安岭的部分地区也将会成为阔叶红松林的高度适宜区。但对于小兴安岭地区的研究却有所不同,当未来年降水增加10%左右、年气温增幅大于5℃时,林窗模型的模拟结果表明阔叶红松林将被蒙古栎、紫椴和裂叶榆为主组成的阔叶林所取代[24],在CGCM2 情景下,也能够得到相同的结论[25],而本文选取SRES-A2、SRES-A1B、SRES-B1三种气候情景,结果有很大的差别。原因是物种分布模型目前有很多不确定性,首先,在模型选取上,现在有很多种类的物种分布模型,不同的物种分布模型预测结果有所差异,其次,环境因子的选取也有很大变化,选取不同的气候情景,预测结果会有所不同[26]。此外,在A2气候情景下,从2020年到2050年阔叶红松林的高度适宜区面积增加,其原因是阔叶红松林的适宜性等级分布是采取4类树种的叠加方式得到的,因此可能会造成累计误差。
物种分布区是物种生态与进化历史的复杂表达形式,在不同的时空尺度上受到多种因素、不同强度的控制[27-28]。通常认为决定物种分布区共有四种因素:环境因素、生物因素、物种的扩散能力、物种适应新环境的进化能力[29-31]。而本文仅仅考虑环境因素中的部分环境因子作为预测指标进行预测,与实际还有一定的差距,还需要做进一步的研究。
4 结论
目前,阔叶红松林主要分布于长白山和小兴安岭地区。但是在A2、A1B、B1气候情景下预测阔叶红松林的分布,结果有很大的差异,其分布界线北移、高度适宜分布区域面积减小,其中长白山地区的阔叶红松林高度适宜分布区更集中于长白山自然保护区。此外,气候变化程度越严重,对阔叶红松林的分布影响也越大,尤其是在A2、A1B气候情景下,到2080年阔叶红松林的高度适宜区面积所占研究区域面积不足1%,说明如果不加以保护环境而使气候得以改善,那么阔叶红松林在未来有可能逐渐消失于东北地区。而影响阔叶红松林分布的主导环境因子主要是年降雨量(Bio12)、季节性降雨量(Bio15)、海拔(ELE)、年平均温度(Bio1)和最湿季度的平均温度(Bio8)。
[1] Yang X Q, Kushwaha S P S, Saran S, Xu J C, Roy P S. Maxent modeling for predicting the potential distribution of medicinal plant,JusticiaadhatodaL. in Lesser Himalayan foothills. Ecological Engineering, 2013, 51: 83-87.
[2] 张新时, 周广胜, 高琼, 倪健, 唐海萍. 中国全球变化与陆地生态系统关系研究. 地学前缘, 1997, 4(1/2): 137-144.
[3] IPCC(Intergovernmental Panel on Climate Change). Climate change 2007: Impacts, adaptation and vulnerability//Parry M L, Canziani O F, Palutikof J P, van der Linden P J, Hanson C E, eds. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2007.
[4] 李国庆, 刘长成, 刘玉国, 杨军, 张新时, 郭柯. 物种分布模型理论研究进展. 生态学报, 2013, 33(16): 4827-4835.
[5] Elith J, Graham C H, Anderson R P, Dudík M, Ferrier S, Guisan A, Hijmans R J, Huettmann F, Leathwick J R, Lehmann A, Li J, Lohmann L G, Loiselle B A, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton J M M, Peterson A T, Phillips S J, Richardson K, Scachetti-Pereira R, Schapire R E, Soberón J, Williams S, Wisz M S, Zimmermann N E. Novel methods improve prediction of species′ distributions from occurrence data. Ecography, 2006, 29(2): 129-151.
[6] Elith J, Graham C H. Do they? How do they? Why do they differ? On finding reasons for differing performances of species distribution models. Ecography, 2009, 32(1): 66-77.
[7] 代力民, 谷会岩, 邵国凡, 王庆礼. 中国长白山阔叶红松林. 沈阳: 辽宁科学技术出版社, 2004.
[8] 代力民, 邵国凡. 森林经营决策——理论与实践. 沈阳: 辽宁科学技术出版社, 2005.
[9] 孙晓红. 中国红松地理分布与气候的关系研究[D]. 北京: 中国科学院研究生院, 2012.
[10] 殷晓洁, 周广胜, 隋兴华, 何奇瑾, 李荣平. 蒙古栎地理分布的主导气候因子及其阈值. 生态学报, 2013, 33(1): 103-109.
[11] 吴正方. 东北阔叶红松林分布区生态气候适宜性及全球气候变化影响评价. 应用生态学报, 2003, 14(5): 771-775.
[12] 王绍强, 周成虎, 刘纪远, 李克让, 杨晓梅. 东北地区陆地碳循环平衡模拟分析. 地理学报, 2001, 56(4): 390-400.
[13] 王新闯. 东北地区森林生态系统碳储量空间分布格局[D]. 沈阳: 中国科学院研究生院, 2011.
[14] 徐 影, 丁一汇, 赵宗慈. 近30年人类活动对东亚地区气候变化影响的检测与评估. 应用气象学报, 2002, 13(5): 513-525.
[15] 秦大河, 陈振林, 罗勇, 丁一汇, 戴晓苏, 任贾文, 翟盘茂, 张小曳, 赵宗慈, 张德二, 高学杰, 沈永平. 气候变化科学的最新认知. 气候变化研究进展, 2007, 3(2): 63-73.
[16] 陈新美, 雷渊才, 张雄清, 贾宏炎. 样本量对Maxent模型预测物种分布精度和稳定性的影响. 林业科学, 2012, 48(1): 53-59.
[17] Sillero N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecological Modelling, 2011, 222(8): 1343-1346.
[18] Moreno R, Zamora R, Molina J R, Vasquez A, Herrera M A. Predictive modeling of microhabitats for endemic birds in South Chilean temperate forests using Maximum entropy (Maxent). Ecological Informatics, 2011, 6(6): 364-370.
[19] IPCC(Intergovernmental Panel on Climate Change). Climate change 2007: The physical science basis//Solomon S Qin D, Manning M, Chen Z, Marquis M, Averyt K B, Tignor M, Miller H L, eds. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2007.
[20] Fitzpatrick M C, Gove A D, Sanders N J, Dunn R R. Climate change, plant migration, and range collapse in a global biodiversity hotspot: theBanksia(Proteaceae) of Western Australia. Global Change Biology, 2008, 14(6): 1337-1352.
[21] Woodward F I. Climate and Plant Distribution. Cambridge: Cambridge University Press, 1987.
[22] 王翼, 娄德君, 曲金华, 张凯. IPCC-AR4模式资料对东北地区气候及可利用水资源的预估研究. 自然资源学报, 2009, 24(9): 1647-1656.
[23] 程肖侠, 延晓冬. 气候变化对中国大兴安岭森林演替动态的影响. 生态学杂志, 2007, 26(8): 1277-1284.
[24] 邓慧平, 吴正方, 周道玮. 全球气候变化对小兴安岭阔叶红松林影响的动态模拟研究. 应用生态学报, 2000, 11(1): 43-46.
[25] 周丹卉, 贺红士, 李秀珍, 周春华, 王绪高, 陈宏伟. 小兴安岭不同年龄林分对气候变化的潜在响应. 北京林业大学学报, 2007, 29(4): 110-117.
[26] Xu D Y, Yan H. A study of the impacts of climate change on the geographic distribution ofPinuskoraiensisin China. Environment International, 2001, 27(2/3): 201-205.
[27] Brown J H. Macroecology. Chicago: University of Chicago Press, 1995.
[28] Franklin J. Mapping Species Distributions: Spatial Inference and Prediction. Cambridge: Cambridge University Press, 2009.
[29] Soberon J, Peterson A T. Interpretation of models of fundamental ecological niches and species′ distributional areas. Biodiversity Informatics, 2005, 2: 1-10.
[30] Soberón J, Nakamura M. Niches and distributional areas: concepts, methods, and assumptions. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(S2): 19644-19650.
[31] Soberón J M. Niche and area of distribution modeling: a population ecology perspective. Ecography, 2010, 33(1): 159-167.
Impacts of climate change on the potential geographical distribution of broadleaved Korean pine (Pinuskoraiensis) forests
JIA Xiang1,2, MA Fangfang1,2, ZHOU Wangming1, ZHOU Li1, YU Dapao1, QIN Jing3, DAI Limin1,*
1KeyLaboratoryofForestEcologyandManagement,InstituteofAppliedEcology,ChineseAcademyofSciences,Shenyang110016,China2UniversityofChineseAcademyofSciences,Beijing100049,China3SeedsandSeedlingsofWoodsManagementStation,Shenyang110036,China
Species-environment relationships have always been a central issue in ecology and biogeography. The adaptation of species to a changing natural environment, including topographical and climatic factors, determines its geographic distribution. Accumulating evidence has revealed that Northeast China has experienced the greatest temperature increase since the 1950s, which in turn affected species distribution in this region. Broadleaved Korea pine (Pinuskoraiensis) mixed forest (BKF) is the native forest type in the southern part of Northeast China. The distribution area of this species has shrunk substantially due to historical overexploitation. It is important for forest managers to be able to predict the potential geographic distribution of BKF based on species-environment relationships. Although the distributions of individual species in BKF such asP.koraiensisandQuercusmongolicahave been reported previously, few studies have focused on the potential geographic distribution of BKF. In this study, four dominant tree species -P.koraiensis,Q.mongolica,TiliaamurensisandFraxinusmandshurica, which together account for more than 80% of the growing stock in primary BKF, were selected to represent this forest type. Nineteen climatic and three topographic variables in Northeast China that are considered to be most likely influences on the geographic distribution of tree species were selected as environmental factors. To identify the major climatic and topographic factors controlling BKF distribution and simulate the potential geographic distribution of BKF under current climatic condition, the geographic distribution records of the dominant tree species, together with the environmental factors, were used in the Maxent model. The future geographic distributions of BKF were consequently predicted for the 2020s, 2050s, and 2080s, based on three kinds of climate change scenarios (SRES-A2, SRES-A1B, SRES-B1) published by the Intergovernmental Panel on Climate Change (IPCC). For the four dominant tree species, the simulations showed that the area under the curve indexes (AUC) were 0.925, 0.890, 0.859, and 0.847, respectively. All these values exceeded 0.8, which indicates that the models had a good predictive performance. The major environmental factors affecting the distribution of BKF included annual precipitation, precipitation seasonality, elevation, annual mean temperature and mean temperature of wettest quarter. For the entire region, 11.69% of the total area was identified to be of high suitability for BKF distribution, 23% was of low suitability, and 65.31% of the area was unsuitable. Under the A2, A1B, and B1 scenarios, the model predicted that both the southern and northern boundary of the high suitability area for BKF will shift northward. Overall, the high suitability area in this region was predicted to decrease, with the extent of the decrease depending on the severity of climate change. For example, under the A2 and A1B scenarios, the high suitability area in Northeast China will be less than 1% of the region by the 2080s. Overall, the results indicate that if no effective measures are taken to mitigate climate change, there is a great possibility that BKF will disappear from Northeast China.
broadleaved Korea pine mixed forest; climate change; Maxent model; potential geographic distribution; climate change scenarios
科技基础性工作专项资助项目(2015FY210200-9);中国科学院特色研究所项目资助项目(Y5YZX151YD)
2015-08-10;
日期:2016-06-13
10.5846/stxb201508101680
* 通讯作者Corresponding author.E-mail: lmdai@iae.ac.cn
贾翔,马芳芳,周旺明,周莉,于大炮, 秦静, 代力民.气候变化对阔叶红松林潜在地理分布区的影响.生态学报,2017,37(2):464-473.
Jia X, Ma F F, Zhou W M, Zhou L, Yu D P, Qin J, Dai L M.Impacts of climate change on the potential geographical distribution of broadleaved Korean pine (Pinuskoraiensis) forests.Acta Ecologica Sinica,2017,37(2):464-473.
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
小哥白尼(野生动物)(2021年9期)2022-01-17 02:31:00
安徽农学通报(2021年22期)2021-12-11 20:37:03
安徽农业科学(2021年14期)2021-09-03 09:55:59
中国林副特产(2020年5期)2020-10-09 08:24:14
炎黄地理(2019年5期)2019-10-20 01:59:19
防护林科技(2017年7期)2017-09-01 01:08:43
安徽农业科学(2016年9期)2016-10-14 22:12:23
建筑与预算(2014年7期)2014-10-14 13:28:52
满族文学(2014年5期)2014-09-22 23:50:36
