
高寒草甸不同类型草地群落根土比、土壤养分变化
2017-01-09 07:21字洪标阿的鲁骥杨有芳王长庭
西南农业学报 2016年12期
字洪标,阿的鲁骥,马 力,胡 雷,陈 焱,杨有芳,王长庭*
(1.西南民族大学青藏高原研究院,四川 成都 610041;2.西南民族大学生命科学与技术学院,四川 成都 610041;3.南京大学生命科学学院,江苏 南京 210093)
高寒草甸不同类型草地群落根土比、土壤养分变化
字洪标1,阿的鲁骥1,马 力2,胡 雷3,陈 焱2,杨有芳2,王长庭2*
(1.西南民族大学青藏高原研究院,四川 成都 610041;2.西南民族大学生命科学与技术学院,四川 成都 610041;3.南京大学生命科学学院,江苏 南京 210093)
为了更好地保护和利用青藏高原高寒草地生态系统功能,了解高寒草甸不同草地类型生态系统结构与功能及生态过程的差异性,本研究对海北站小嵩草(Kobresiapygmaea)草甸、杂类草草甸、矮嵩草(K.humilis)草甸、藏嵩草(K.tibetica)沼泽化草甸植物群落的根土比、土壤容重和土壤养分状况进行了比较研究。结果表明,不同植被类型高寒草甸根土比均随着土层深度增加而显著降低,且藏嵩草沼泽化草甸根土比最高(P< 0.05);土壤容重随着土层深度而增加,其中藏嵩草草甸显著低于其他草甸(P< 0.05);土壤养分随着土层深度增加而显著降低,且在藏嵩草沼泽化草甸下养分含量最高(P< 0.05)。土壤养分与根土比主要呈正相关关系,而与土壤容重则主要呈负相关关系。因此,高寒草甸不同类型草地群落根土比大小、土壤养分含量反映了高寒草甸群落类型及稳定性,从而可作为判断高寒草甸生态系统功能可持续利用的关键指标之一。
高寒草甸;根土比,植被类型;土壤养分
高寒草甸作为青藏高原草地生态系统的主要类型,占青藏高原面积的19.2 %,是当地畜牧业发展的主要基础[1]。近年来,由于受全球气候变化和人类活动的双重干扰,青藏高原高寒生态系统也正在发生着剧烈的变化[2]。从而导致草地退化,生物多样性下降,水土流失日益严重等一系列生态问题,给当地生态环境与畜牧业的可持续发展造成了严重威胁[3]。维持高寒生态系统稳定以及可持续利用是当前生态学家面临的重要难题之一,同时也是制约当地经济发展的主要因素。因此,迫切需要对高寒草甸生态系统进行合理监测、保护和管理、利用[4]。
高寒草甸作为青藏高原典型的地带性植被类型,虽然其所处的大气环境属性基本相同,但是受土壤温度、湿度、地下水及土壤性状等综合作用的影响,形成了差异较大的不同植被类型草地[5]。目前,许多学者对高寒草甸生态系统结构和功能、土壤微环境[6-13]等方面进行大量的研究。但是对生态系统结构、功能与过程研究中最不确定的因素——地下部分关注相对较少,特别是高寒草甸不同类型草地生态系统根系生产、时空分布格局;根系与土壤微环境之间互作机制等,因而严重制约着高寒草甸生态系统的功能研究及理论拓展。因此,地上和地下的整合目前被认为是生态系统研究的最有效途径[15]。
根系作为连接地上与地下的桥梁[16]。在植物固定和支撑[17]、植被碳库与土壤碳库交换[18]等方面具有重要作用。其生存的“载体”土壤,不仅是生态系统中环境与生物相互作用的产物,而且也是陆地生态系统中物质与能量相互交换的重要场所,并储藏着碳、氮、磷等大量的营养物质[19],影响着植物群落物种组成、结构、功能和生态系统的稳定性。一方面,植物根系与土壤的接触面极大,土壤中的矿质营养和水分需要植物根系的吸收并输送到地上部分供自身利用[20];同时,在这个转移过程中植物根系也积累和储存了大量矿质元素,其死亡后,又归还给土壤,从而促进了植物与土壤的养分循环[21]。另一方面,根系在土壤中纵横穿梭,有利于土壤团粒结构的形成,改善土壤的物理性质,其死亡后又增加土壤的通气性和渗透压;同时根系还会分泌某些化学物质促进微生物的活动和营养元素的吸收,改善土壤化学和生物学特性[22]。土壤容重、水分以及C、N含量又与植物根系在土层中的垂直分布存在着一定的相关性[23],地下根系空间分布格局特征又决定着植物与土壤环境之间作用面的大小[24]。因此,研究高寒草甸不同类型草地植物根系、根系与土壤环境相互作用机制,为认知和确定高寒草地的群落稳定性和退化程度;防止草地和土壤进一步退化提供理论和实践依据。
1 材料与方法
1.1 研究区域概况
海北站地处青藏高原东北隅祁连山北支冷龙岭东段南麓,地理位置为37°29′~37°45′N,101°12′~101°23′E,海拔在3200~3600 m。该地区气候具有明显的高原大陆性气候,季风气候偏弱。气温极低,无明显四季之分,仅有冷暖季之别,冷季漫长、干燥而寒冷,暖季短暂、湿润而凉爽。年平均气温-1.7 ℃,最冷的1月平均气温-14.8 ℃,最暖的7月平均气温9.8 ℃。年平均降水量为600 mm左右,降水主要集中在5-9月,约占年降水量的80 %,蒸发量为1160.3 mm。特殊的自然环境及青藏高原隆起过程,造就了适应寒冷湿中生的多年生草本植物群落,形成了不同的植被类型,即杂类草草甸、矮嵩草(K.humilis)草甸、小嵩草(K.pygmaea) 草甸和藏嵩草 (K.tibetica) 沼泽化草甸等植被类型。主要土壤类型为高山草甸土(Typic alpine meadow soil)、高山灌丛草甸土(Alpine shrubby meadow soil)和沼泽土(Bog soil)[20]。
1.2 试验设计
于2010年8月中旬植物生长旺季(草盛期),选择典型的杂类草甸、矮嵩草草甸、小嵩草草甸、藏嵩草沼泽化草甸为试验样地,样地面积分别为50 m×50 m,以“S”型取样方法,用内径5 cm土钻随机分层(0~10 和10~20 cm)采集土壤样品,每个样地采集50钻,每10钻土壤混合为1个土壤样品,5次重复,拣去石子、根系和残体后编号,带回实验室,风干过2 和0.25 mm筛,用于土壤养分的测定。土壤根土体积比采用容量瓶法[21],用5 cm土钻在0~20 cm土层每10 cm取土柱,将土柱装入500 mL容量瓶中,加水V1至刻度,倒出;收集草根,将其洗净并用滤纸吸干水分,在装入500 mL容量瓶中,加水V2至刻度。得草根体积V根=500-V2,土体积V土=V2-V1,根土体积比=V根/V土,重复5次;根土重量比采用土钻法[22],在每个观测样方用内径5 cm土钻在0~20 cm土层每10 cm取样,共2层,每层取10钻,重复5次,样品过筛得到土壤样品并称重,清水冲洗得到根系样品,分别在60 和 105 ℃烘至恒量,称干重量,根土重量比=根重量/土壤重量。土壤容重用环刀法测定[25],每个样地重复5次。土壤养分的测试项目为样品的全磷(钼锑抗比色法)、速效磷(磷酸氢钠浸提—钼锑抗比色法)、全氮(凯氏法和康维皿法)、硝态氮[用2M KCl(土液比1∶4)浸提,流动分析仪测定]、全钾和速效钾(火焰光度法)[26]。
1.3 数据分析
采用Excel 2010 进行数据的基本统计和分析制图,SPSS 19.0统计软件进行单因素方差分析及多重检验法(LSD)检验各处理间的差异性,用Duncan进行显著性分析(P= 0.05),利用线性相关(Pearson)对根土比、土壤容重与土壤养分进行相关性分析(P= 0.05,0.01)。

表1 不同植被类型根土比和土壤容重的变化
注:不同大写字母代表不同植被类型的显著性差异(P<0.05);不同小写字母代表的是相同植被类型不同土层的显著性差异(P<0.05)。下同。 Note:The different lowercase letters indicated that vegetation types had significant differences at 0.05 level,and different capital letters showed that the same vegetation types in soil depth had significant differences at 0.05 level. The same as below.
2 结果与分析
2.1 不同植被类型高寒草甸根土比和土壤容重的变化
由表1可知,不同植被类型根土比表现为:藏嵩草沼泽化草甸>小嵩草草甸>矮嵩草草甸>杂类草草甸。藏嵩草沼泽化草甸根土比远远大于其他草甸的比值,是其他草甸的5倍以上,且与其他草甸存在显著差异(P<0.05)。在不同草甸类型其根土比均表现出随着土层深度的增加而降低,且不同土层之间差异不显著(P> 0.05)。土壤容重变化则表现出:杂类草草甸>矮嵩草草甸>小嵩草草甸>藏嵩草沼泽化草甸。不同植被类型土壤容重之间显著差异(P<0.05)。不同植被类型0~10 cm土壤容重显著低10~20 cm土层(P<0.05)。
2.2 不同植被类型高寒草甸土壤养分的变化
由表2可知,不同植被类型全氮含量变化:藏嵩草草甸>小嵩草草甸>矮嵩草草甸>杂类草草甸 (P<0.05),0~10 cm土层全氮明显高于10~20 cm土层(P<0.05)。全磷含量:藏嵩草草甸>矮嵩草草甸>杂类草草甸>小嵩草草甸(P<0.05)。全钾含量藏嵩草沼泽化草甸显著低于其他草甸,且不同土层之间存在显著差异(P<0.05)。硝态氮则表现出矮嵩草草甸>藏嵩草草甸>小嵩草草甸>杂类草草甸,且藏嵩草草甸、矮嵩草草甸明显高于小嵩草草甸、杂类草草甸(P<0.05)。速效磷含量矮嵩草草甸和藏嵩草草甸显著高于小嵩草草甸和杂类草草甸(P<0.05)且均表现出随土层深度的增加而降低。速效钾含量变化规律不明显,仅杂类草草甸0~10 cm土层含量较高。
2.3 不同植被类型高寒草甸根土比和土壤容重与土壤养分的相关性分析
在小嵩草草甸中,土壤全氮和速效钾与根土体积比和根土质量比均呈显著正相关(P<0.05); 土壤容重与全氮呈显著负相关(P<0.05)。在杂类草草甸中,土壤全氮、全磷和速效磷与根土体积比和根土质量比均呈显著正相关(P<0.05);速效磷和速效钾与土壤容重呈显著负相关(P<0.05)。在藏嵩草沼泽化草甸中,全氮、硝态氮、速效磷和速效钾与土壤容重呈显著负相关(P<0.05);仅土壤速效磷与根土质量比呈显著正相关(P<0.05)。在矮嵩草草甸,土壤全氮和含量与根土质量比呈显著正相关,速效钾与根土体积比呈显著正相关(P<0.05);土壤容重与全氮、速效磷、速效钾和全钾均呈显著负相关(P<0.05)。可以看出,土壤养分与根土比主要呈正相关关系,而与土壤容重呈负相关关系(表3)。

表2 不同植被类型高寒草甸土壤养分的变化
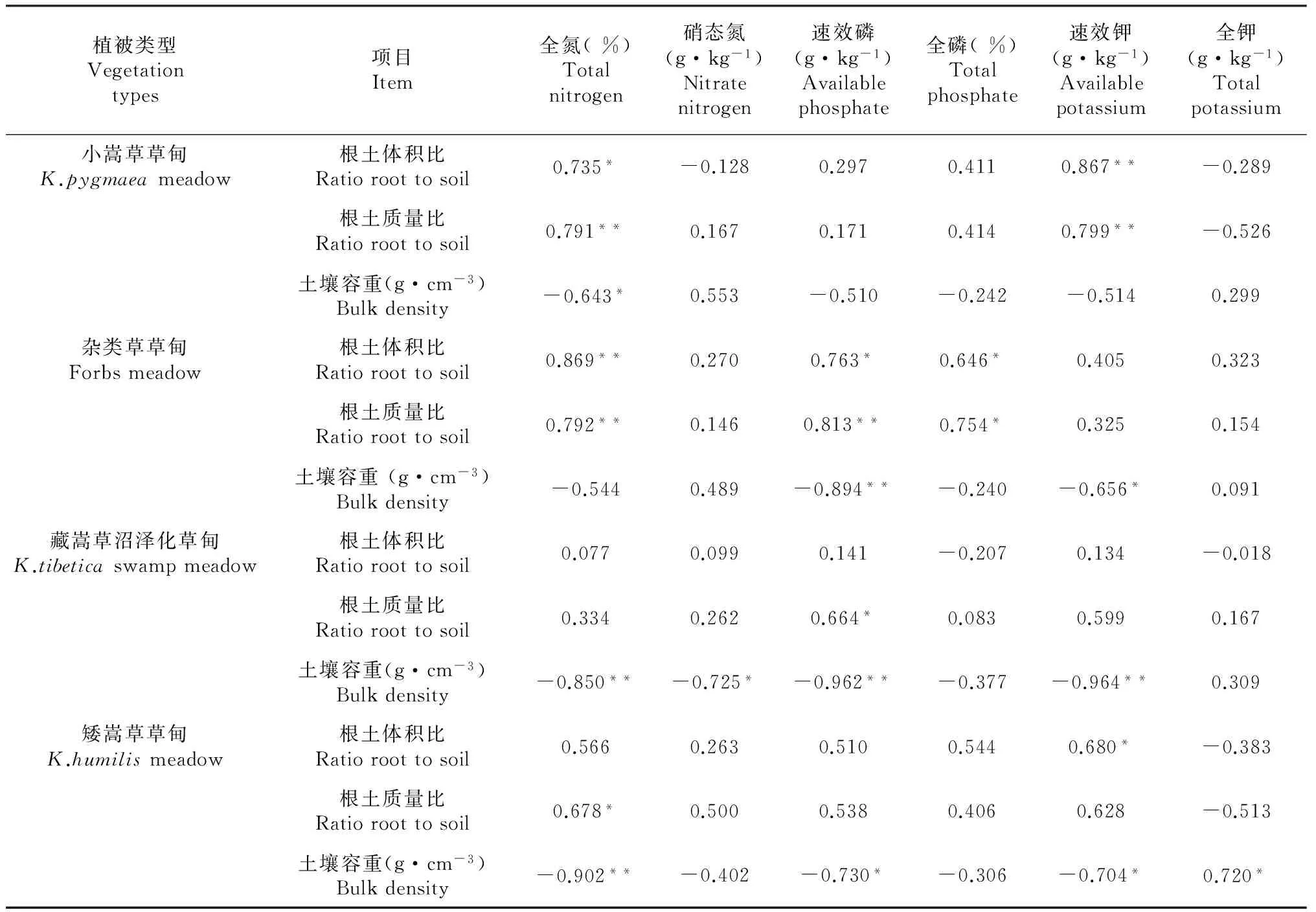
表3 不同植被类型高寒草甸根土比和土壤容重与土壤养分的相关性分析
注:* 表示显著相关(P< 0.05);** 表示极显著相关(P< 0.01)(双尾检验)。
Note:* indicated significant correlation at 0.05 level; ** indicated significant correlation at 0.01 level (2-tailed test).
3 讨论与结论
根系作为连接地上和地下的重要组成部分[16],是植物吸收养分及水分的主要器官[27],在养分合成、植物固定支撑等方面具有重要作用[28-29],对生态系统有效生态功能的发挥和地下部分生态位的分化产生重要影响[30]。植物根系分布范围和地下生态位的变化又反映出地上植物可利用资源的范围以及它在演替过程中的作用。而土壤是孕育根系的载体,对植被群落演替产生重要作用,不仅促进前一群落的灭亡,而且还为后续群落演替创造条件[31]。植物根系的时空分布格局与孕育根系土壤的量对于植物群落结构、功能以及生态系统的稳定性具有重要意义[24]。本研究发现高寒草甸不同类型草地群落的根系随着深度的增加而显著降低,且主要集中在0~10 cm土层中。这Wang等[32]研究结果相似。因为植物群落中资源分布从地表向下依次递减(表2),而草本植物根系细小[33],主要利用的是土壤表层的水分[34]和养分[35],从而促使大量根系集中在表层。此外,不同草地类型植物群落物种组成、多样性和生产力差异[5, 10, 36];土壤环境的异质性,特别是孕育根系的土壤量不同,进而引起根土比的改变。其中,藏嵩草沼泽化草甸根土比显著大于其他类型草甸,说明藏嵩草沼泽化草甸的植物根系十分发达,根系生物量较高,养分含量的高低影响着根土比的大小(表2)。而究其原因可能是由环境因子或是人为因子抑或是二者共同作用的结果[37],一方面,藏嵩草沼泽化草甸植物生长在沼泽地带,水分条件较好,生境相对均一,资源异质性小[38];另一方面,由于其特殊生境条件在雨季食草家畜、人以及其他食草类动物很难涉及其内,从而导致外界干扰低[39];还有藏嵩草沼泽化草甸的土壤环境特征具有明显的高湿和低温[40],作为藏嵩草沼泽化草甸优势植物藏嵩草又是耐寒喜湿的植物[41],正是这种适宜条件促使根系大量繁殖,从而表现出较高的根土比。
土壤容重是表征土壤紧实度的指标之一,它与土壤孔隙度和渗透率密切相关[42],其大小深受植物根系含量影响[22]。因为根系在土壤中纵横穿行,有利于形成团粒结构,改善土壤的物理性质,其死亡后留下的孔隙能增加土壤的透气性和渗透性[20]。本研究中,土壤容重在高寒草甸不同类型草地均表现出随着土层深度的增加而增加,这说明表层土壤根系丰富,深层土壤根系较匮乏,从而导致土壤容重在不同土层上的差异性。在不同类型草地由于其土壤类型、温湿度及植被种类等不同[5],土壤容重差异也比较大。
土壤作为生态系统中生物与环境相互作用的基质,贮存着大量的碳、氮、磷等营养物质,而土壤中氮、磷、钾等营养元素对植物生长起着至关重要的作用,直接影响着植物群落的种类组成与生活性特征,决定着生态系统的结构和功能[43]。本研究中,高寒草甸不同类型草地土壤全氮、全磷、速效磷、速效钾均随土层深度的增加而降低,全钾含量随着土层深度的增加而增加。这表明高寒草甸土壤营养成分在空间上是异质性分布的[44],且主要集中在土壤表层。这是由于表层土壤透气性良好,微生物活动强烈,加快了土壤养分转化[45];也可能够是植物凋落物影响了土壤基质的质量与数量、土壤水分等物理和化学性质[46]。还有最重要一方面是根系,因为土壤营养异质性分布将会影响植物根系的生长[43]。植物根系是陆地生态系统重要的碳汇和养分库,其中细根周转是地下部分的碳氮循环的核心环节,在生态系统中起着关键作用[18]。细根周转进入土壤的碳占土壤总碳输入碳量的14 %~50 %[47],在有些生态系统甚至是地上凋落物碳输入量的1.6倍[48]。而对于草本植物细根主要集中在表层,因此,养分含量呈现出浅层化的垂直分布格局[49]。在不同类型草地土壤养分由于植物根系深度[50]、植物对养分利用能力[51]以及相对吸收率[52]等不同而存在明显差异。
综上所述,植物根系空间结构不仅决定了根系对地下资源的利用效果及潜力,同时还反映了土壤中水分和养分的分配格局。高寒草甸植物根系与土壤量的比例(根土比)反映了高寒草甸植物根系生物量变化和空间分布规律,土壤养分和容重的变化与根土比密切相关,根土比大小反映出地下根系和土壤根系的“载体”量的变化过程,对判别不同自然环境条件下高寒草甸退化演替阶段、退化程度有着重要作用。课题组在分析和判断高寒草甸退化程度、演替阶段的量化指标——根土比时发现,其大小可能存在一个阈值,当超过或低于这个阈值时生态系统稳定性、生产力和生态服务价值等功能受到影响。因此,利用根系、土壤根系的“载体”量和根土比变化规律,能合理地判断草地现时演替阶段、草地演替的驱动力及其演替趋势。
[1]王启兰, 王 溪, 曹广民, 等. 青海省海北州典型高寒草甸土壤质量评价[J]. 应用生态学报, 2011, 22(6):1416-1422.
[2]王一博, 王根绪, 张春敏, 等. 高寒植被生态系统变化对土壤物理化学性状的影响[J]. 冰川冻土, 2007, 29(6):921-927.
[3]李文华. 青藏高原生态系统及优化利用模式[M]. 广州:广东科技出版社, 1998.
[4]喻小勇, 邵全琴, 刘纪远,等. 三江源区不同退化程度的高寒草甸光谱特征分析[J]. 地球信息科学学报, 2012, 14(3):398-404.
[5]杨时海, 李英年, 蒲继延,等. 三种高寒草甸植被类型植物群落结构及其土壤环境因子研究[J]. 草地学报, 2006,14(1):77-83.
[6]杨晓霞, 任 飞, 周华坤,等. 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应[J]. 植物生态学报, 2014,38(2):159-166.
[7]李 娜, 王根绪, 杨 燕,等. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响[J]. 生态学报, 2011, 31(4):895-905.
[8]高 洋, 王根绪, 高永恒. 长江源区高寒草地土壤有机质和氮磷含量的分布特征[J]. 草业科学, 2015(10):1548-1554.
[9]王长庭, 龙瑞军, 王根绪,等. 高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究[J]. 草业学报, 2010, 19(6):25-34.
[10]王长庭, 龙瑞军, 刘 伟,等. 高寒草甸不同群落类型土壤碳分布与物种多样性、生物量关系[J]. 资源科学, 2010, 32(10):2022-2029.
[11]胡 雷, 王长庭, 王根绪,等. 三江源区不同退化演替阶段高寒草甸土壤酶活性和微生物群落结构的变化[J]. 草业学报, 2014(3):8-19.
[12]Wang C, Long R, Wang Q, et al. Fertilization and litter effects on the functional group biomass, species diversity of plants, microbial biomass, and enzyme activity of two alpine meadow communities[J]. Plant & Soil, 2010, 331(1-2):377-389.
[13]Man Hou PENG, Quan Gang. Effects of warming and clipping on plant and soil properties of an alpine meadow in the Qinghai-Tibetan Plateau, China[J]. Journal of Arid Land, 2015, 7(2):189-204.
[14]Zhang B, Chen S Y, Zhang J F, et al. Depth-related respopnse of soil microbial communities to experimental warming in an alpine meadow on the Qinghai-Tibet Plateau[J]. European Journal of Soil Science, 2015, 66(3):496-504.
[15]André H, Ducarme X, Anderson J, et al. Skilled eyes are needed to go on studying the richness of the soil. Nature, 2001, 409:761.
[16]Norby R J, Joanne L, Reilly C D, et al. Fine-root production dominates response of a deciduous forest to atmospheric CO2enrichment.[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(26):9689-9693.
[17]周本智, 张守攻, 傅懋毅. 植物根系研究新技术Minirhizotron的起源、发展和应用[J]. 生态学杂志, 2007, 26(2):253-260.
[18]吴伊波, 车荣晓, 马 双,等. 高寒草甸植被细根生产和周转的比较研究[J]. 生态学报, 2014, 34(13):3529-3537.
[19]Robertson G P, Vitousek P M. Nitrification Potentials in Primary and Secondary Succession[J]. Ecology, 1981, 62(2):376-386.
[20]王长庭, 王启兰, 景增春,等. 不同放牧梯度下高寒小嵩草草甸植被根系和土壤理化特征的变化[J]. 草业学报, 2008, 17(5):9-15.
[21]李 鹏, 赵 忠, 李占斌,等. 植被根系与生态环境相互作用机制研究进展[J]. 西北林学院学报, 2002, 17(2):26-32.
[22]韦兰英, 上官周平. 黄土高原不同演替阶段草地植被细根垂直分布特征与土壤环境的关系[J]. 生态学报, 2006, 26(11):3740-3748.
[23]杨丽韫, 罗天祥, 吴松涛. 长白山原始阔叶红松(Pinuskoraiensis)林及其次生林细根生物量与垂直分布特征[J]. 生态学报, 2007, 27(9):3609-3617.
[24]王长庭, 王根绪, 刘 伟,等. 植被根系及其土壤理化特征在高寒小嵩草草甸退化演替过程中的变化[J]. 生态环境学报, 2012, 21(3):409-416.
[25]孔凡伟. 如何精测土壤容重[J]. 黑龙江农业科学, 2010(10):178-178.
[26]鲍士旦.土壤农化分析(第三版)[M]. 北京:中国农业出版社, 2000:30-107.
[27]张小全, 吴可红. 森林细根生产和周转研究[J]. 林业科学, 2001, 37(3):126-138.
[28]丁 红, 张智猛, 戴良香,等. 干旱胁迫对花生根系生长发育和生理特性的影响[J]. 应用生态学报, 2013, 24(6).
[29]龙 毅, 孟凡栋, 王常顺,等. 高寒草甸主要植物地上地下生物量分布及退化对根冠比和根系表面积的影响[J]. 广西植物, 2015(4):532-538.
[30]胡建忠, 郑佳丽, 沈晶玉. 退耕地人工植物群落根系生态位及其分布特征[J]. 生态学报, 2005, 25(3):481-490.
[31]王国宏. 黄土高原自然植被演替过程中的植物特征与土壤元素动态[J]. 植物学报:英文版, 2002, 44(8):990-998.
[32]Wang C T, Long R J, Wang Q J, et al. Effects of altitude on plant-species diversity and productivity in an alpine meadow, Qinghai-Tibetan plateau[J]. Australian Journal of Botany, 2007, 55(2):110-117.
[33]李 鹏, 李占斌, 鲁克新. 黄土区草本植被根系与土壤垂直侵蚀产沙关系研究[J]. 植物生态学报, 2006(2):302-306.
[34]Oliveira R S, Bezerra L, Davidson E A, et al. Deep root function in soil water dynamics in cerrado savannas of central Brazil[J]. Functional Ecology, 2005, 19(4):574-581.
[35]汪诗平, 王艳芬, 李永宏,等. 不同放牧率对草原牧草再生性能和地上净初级生产力的影响[J]. 草地学报, 1998(4):275-281.
[36]李英年, 赵新全, 王勤学,等. 青海海北高寒草甸五种植被生物量及环境条件比较[J]. 山地学报, 2003, 21(3):257-264.
[37]王艳芬, 汪诗平. 不同放牧率对内蒙古典型草原地下生物量的影响[J]. 草地学报, 1999(3):198-203.
[38]王彦龙, 马玉寿, 施建军,等. 黄河源区高寒草甸不同植被生物量及土壤养分状况研究[J]. 草地学报, 2011, 19(1):1-6.
[39]周兴民. 中国嵩草草甸[M]. 北京:科学出版社, 2001.
[40]李英年, 鲍新奎, 曹广民. 祁连山海北高寒湿地40~80 cm土壤温度状况观测分析[J]. 冰川冻土, 2000(2):153-158.
[41]赵新全. 高寒草甸生态系统与全球变化[M]. 北京:科学出版社, 2009.
[42]戎郁萍, 韩建国, 王 培,等. 放牧强度对牧草再生性能的影响[J]. 草地学报, 2001(2):92-98.
[43]王长庭, 龙瑞军, 曹广民,等. 高寒草甸不同类型草地土壤养分与物种多样性——生产力关系[J]. 土壤通报, 2008(1):1-8.
[44]Wardle D. Is ‘sampling effect’ a problem for experiments investigating biodiversity-ecosystem function relationships?[J]. Oikos, 1999, 87(2):403-407.
[45]田玉强, 欧阳华, 宋明华,等. 青藏高原样带高寒生态系统土壤有机碳及氮素分布特征研究[C].青藏高原环境与变化研讨会论文摘要汇编,2005.
[46]王其兵, 李凌浩. 模拟气候变化对3种草原植物群落混合凋落物分解的影响[J]. 植物生态学报, 2000(6):674-679.
[47]Vogt K A, Grier C C, Vogt D J. Production, Turnover, and Nutrient Dynamics of Above-and Belowground Detritus of World Forests[J]. Advances in Ecological Research, 1986, 15(15):303-377.
[48]杨丽韫, 李文华. 长白山原始阔叶红松林细根分布及其周转的研究[J]. 北京林业大学学报, 2005, 27(2):1-5.
[49]侯扶江, 肖金玉, 南志标. 黄土高原退耕地的生态恢复[J]. 应用生态学报, 2002, 13(8):923-929.
[50]Wardle D A, Zackrisson O, Hörnberg G, et al. The influence of island area on ecosystem properties[J]. Science, 1997, 277(21):1296-1299.
[51]Loreau M, Naeem S, Inchausti P, et al. Biodiversity and ecoystem functioning:current knowledge and future research[J]. Science, 2001,294:804-808.
[52]Gross K L, Pregitzer K S, Burton A J. Spatial variation in nitrogen availability in three successional plant communities[J]. Journal of Ecology,1995, 83(3):357-367.
(责任编辑 陈 虹)
Change of Ratio of Root to Soil and Soil Nutrient Content at Different Grassland Types in Alpine Meadow
ZI Hong-biao1, ADE Lu-ji1, MA Li2, HU Lei3, CHEN Yan2, YANG You-fang2,WANG Chang-ting2*
(1.College of Tibetan Plateau Research, Southwest University for Nationalities, Sichuan Chengdu 610041, China; 2.College of Life and Technology, Southwest University for Nationalities, Sichuan Chengdu 610041, China; 3.School of Life Sciences, Nanjing University, Jiangsu Nanjing 210093, China)
In order to protect and reasonably utilize the ecosystem function of alpine meadow and to know the difference of the structure, function and ecological process, in this study, the ratio of root to soil, soil bulk density and soil nutrition at the different grassland types of alpine meadow communities in Haibei Research Station of Alpine Meadow Ecosystem were analyzed. The results showed that the ratio of root to soil significantly decreased with soil depth in different vegetation types of the alpine meadow, and the ratio of root to soil inKobresiatibeticaswamp meadow was the highest in three alpine meadows(P< 0.05); The soil bulk density significantly increased with soil depth increasing, in whichK.tibeticaswamp meadow was the lowest in three alpine meadows(P<0.05); Soil nutrient content significantly reduced with soil depth increasing, and the biggest nutrient contents occurred in theK.tibeticaswamp meadow. The soil nutrient content was significantly positively correlated with the ratio of root to soil, while negatively correlated with soil bulk density. Therefore, the ratio of root to soil and soil nutrient content in alpine meadow reflected the community types and stability at different grassland types, which could be used as a key indicator to estimate the sustainable utilization of the ecosystem function in alpine meadow.
Alpine meadow; Ratio of root to soil; Vegetation type; Soil nutrient content
1001-4829(2016)12-2916-06
10.16213/j.cnki.scjas.2016.12.026
2016-01-20
国家自然科学基金(31370542);中央高校基本科研业务费优秀科研及重大孵化项目(2014NZYTD01)
字洪标(1989-),男,彝族,云南永平人,在读硕士,主要从事恢复生态学,E-mail: zhbyn2010@163.com;*为通讯作者:E-mail: wangct@swun.edu.cn。
S156
A
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
幼儿100(2020年31期)2020-11-18
疯狂英语·初中版(2019年4期)2019-09-10
活力(2019年21期)2019-04-01
小太阳画报(2018年6期)2018-05-14
现代园艺(2017年21期)2018-01-03
河北林业科技(2016年5期)2016-11-08
绿色科技(2016年16期)2016-10-11
亚热带资源与环境学报(2015年1期)2015-01-22
