
红椿半同胞家系生长与早期选择
2016-12-21 03:09吴际友李志辉黄明军陈明皋廖德志吴其军
中南林业科技大学学报 2016年4期
吴际友 ,李 艳 ,李志辉 ,黄明军 ,陈明皋 ,程 勇 ,廖德志 ,刘 球 ,吴其军
(1.中国林科院湖南分院,湖南 长沙 410004;2. 湖南省优质用材工程技术研究中心,湖南 长沙 410004;3. 中南林业科技大学,湖南 长沙 410004;4. 湖南金淋林业有限公司,湖南 桃源 415701)
红椿半同胞家系生长与早期选择
吴际友1,2,李 艳3,李志辉3,黄明军1,2,陈明皋1,2,程 勇1,2,廖德志1,刘 球1,2,吴其军4
(1.中国林科院湖南分院,湖南 长沙 410004;2. 湖南省优质用材工程技术研究中心,湖南 长沙 410004;3. 中南林业科技大学,湖南 长沙 410004;4. 湖南金淋林业有限公司,湖南 桃源 415701)
以5年生红椿20个半同胞家系测定林为研究对象,研究了半同胞家系生长性状,并进行遗传力分析。结果表明:红椿半同胞家系的树高和胸径生长存在极显著差异,具有较高的遗传改良潜力;5年生红椿半同胞家系树高的遗传力为0.723 5,胸径的遗传力为0.730 6;通过Duncan法多重比较筛选出TC20、TC04、TC03、TC02、TC12等5个红椿半同胞家系为优良家系。
红椿,半同胞家系,遗传力
红椿Toona ciliataRoem别名红楝子,楝科香椿属高大乔木,国家Ⅱ级重点保护野生植物,是我国热带、亚热带地区的珍贵速生用材树种。其木材为上等家具用材,素有“中国桃花心木”之称[1-2]。红椿在我国主要分布在云南、广西、湖南、贵州及四川等省。由于环境变化、过度开发以及天然更新较慢,其数量不断减少[3]。红椿发展潜力很大,目前已成为我国南方重要的珍贵速生用材林造林树种和营造针阔混交林进行树种结构调整的首选树种之一[4]。近年来,我国科研人员对红椿地理分布、生态特性、天然更新、扦插繁殖、人工栽培、培育模式进行了研究报道[5-20],而对红椿遗传改良方面的研究报道不多。本论文以5年生的红椿半同胞家系测定林为研究对象,通过对测定林的生长性状测定及性状遗传力分析,筛选出了优良的红椿半同胞家系,为红椿产业发展提供了支撑。
1 材料与方法
1.1 试验材料
参试材料为中国林科院湖南分院研究团队在湖南省内选择的20株(编号为:TC01~TC20)红椿优树的种子播种育苗而成,2008年在湖南金淋林业有限公司育苗,2009年在汨罗科技园建立半同胞家系测定林。
1.2 试验地概况
试验地设于湖南汨罗林业科技示范园。科技示范园属亚热带湿润季风气候区,地理位置 为 113°06′E,28°49′N, 海 拔 90 m。 年 均 气温16.9℃,极端最高温39.7℃,极端最低温-13.4℃。年均降水量1 353 mm,无霜期263 d,年均日照时数714.9 h,年均相对湿度81%,土壤主要为第四纪网纹层红土。
1.3 试验方法
半同胞家系测定林均采用一年生实生苗造林,试验采用完全随机区组设计,20个半同胞家系按5株单行小区排列,重复3次。同一区组的立地条件一致。测定林造林密度为2 m×3 m,测定林边缘设置2~3行保护行。造林后前3年每年除草抚育2次,以后每年除草抚育1次。每年12月对测定林进行每木调查,测定胸径、树高、冠幅、枝下高等4个表型性状。
1.4 遗传力参数的计算[21]
表型变异系数(CV)/%=σp/x×100。式中σp为表型标准差,x为平均值。
遗传变异系数(GCV)/%=σg/x×100。式中σg为遗传型标准差,x为平均值。
遗传型方差σg2=1/r(Mf-Me)。式中σg2为遗传型方差,Mf为家系均方,Me为环境均方,r为组内重复数。
家系遗传力(hf2)=(Mf-Me)/Mf。式中Mf为家系均方(既包含家系的遗传变量又包含环境变量),Me为环境均方。
2 结果与分析
2.1 红椿半同胞家系测定林生长情况
从表1可以看出:5年生红椿半同胞家系测定林林分平均树高为5.8 m,其中最小值是TC9家系树高4.6 m,最大值是TC20家系树高7.2 m,两者相差2.6 m,其表型变异系数为13.8%。胸径平均值为6.9 cm,其中最小值是TC09家系胸径5.4 cm,最大值是TC12家系胸径8.8 cm,两者相差3.4 cm,其表型变异系数为13%。枝下高平均值为2.3 m,其中最小值是2.1 m,最大值是3.0 m,其表型变异系数为8.7%,冠幅的平均值为3.0,其中最大值为3.9 m,最小值为2.3 m,其表型变异系数为13.3%
2.2 5年生红椿半同胞家系测定林生长差异分析
对红椿半同胞家系测定林的生长调查数据进行方差分析,结果见表2。
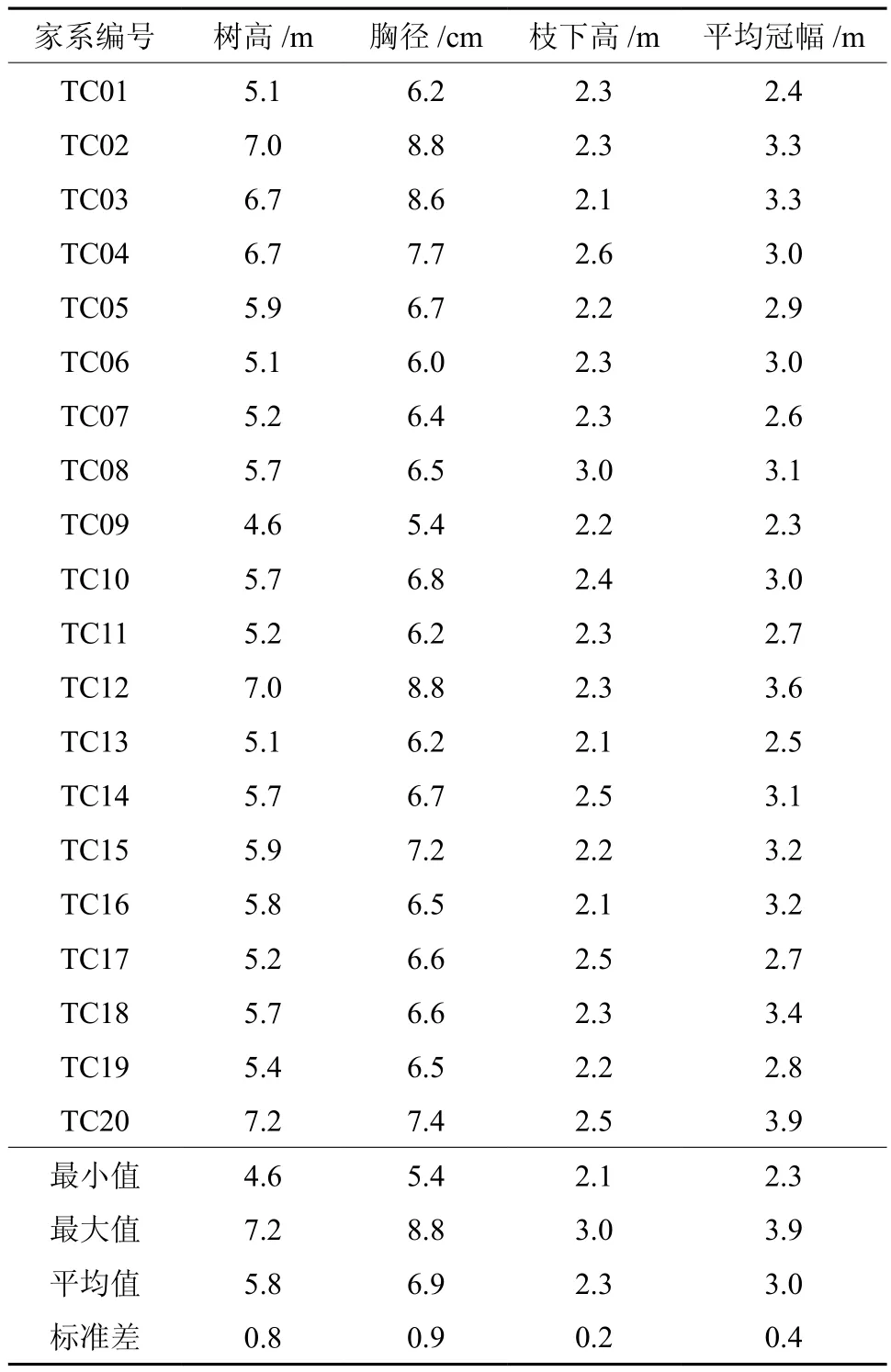
表1 5年生红椿半同胞家系测定林生长情况表(3重复平均)Table 1 Growth of half-sib families of 5-year-old Toonaciliata
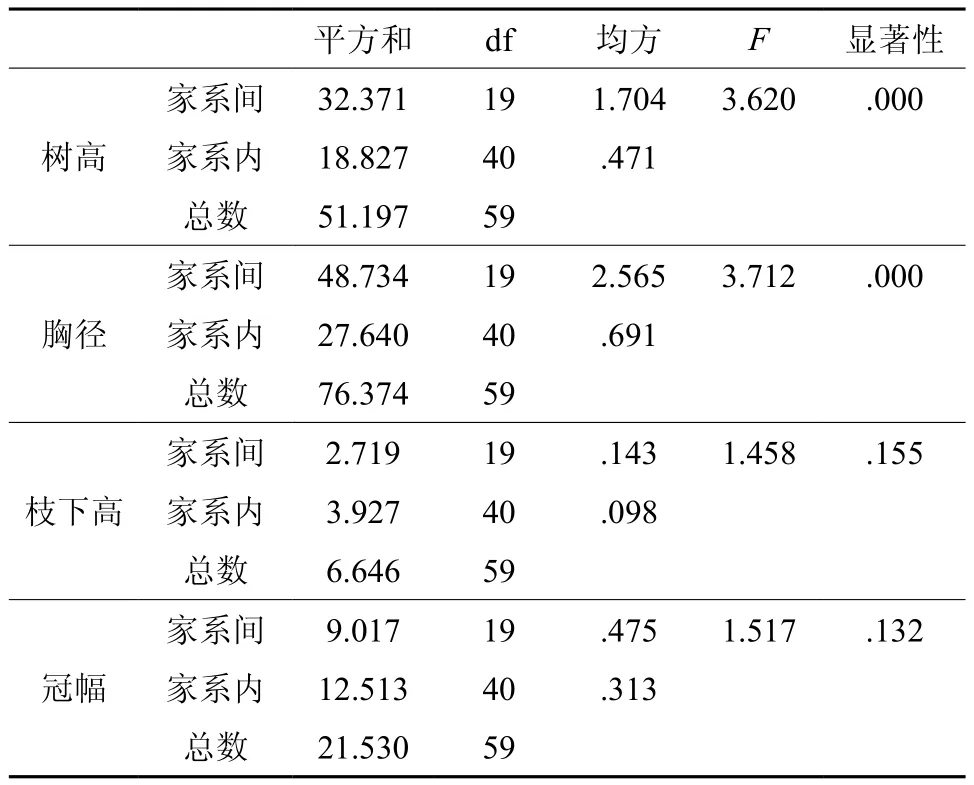
表2 5年生红椿半同胞家系测定林生长因子方差分析Table 2 Variance analysis of growth factor of Toona ciliate half-sib families
从表2可以看出,5年生红椿半同胞家系测定林家系间树高和胸径的差异均达到极显著水平,表明各家系测定林的生长性状存在丰富的变异,具有很大遗传改良潜力。枝下高和冠幅这2个生长性状没有显著差异。
2.3 红椿半同胞家系树高和胸径遗传力分析
因为红椿的枝下高和冠幅这2个生长性状与造林密度密切相关,造林密度越大,枝条自然整枝越强烈,枝下高就相对升高,冠幅长度变小,枝下高和冠幅这2个生长性状受环境影响较大,而不是遗传基因控制,因此不对其进行遗传力分析。

表3 红椿半同胞家系测定林遗传力分析Table 3 Heritability analysis of Toona ciliate half-sib families
经计算,红椿半同胞家系树高的遗传变异系数为11.03%,家系遗传力为0.7235,胸径的遗传变异系数为11.41%,家系遗传力为0.7306。结果表明20个红椿半同胞家系树高和胸径的生长性状差异主要由遗传因素决定,这为红椿优良家系选择提供了依据。
2.4 红椿半同胞家系选择
为了进一步选择出优良的红椿半同胞家系,分别对5年生红椿半同胞家系测定林的树高和胸径数据进行Duncan多重比较(邓肯法),分析结果见表4、表5。根据邓肯法的分组,树高生长表现最好的为 TC04、TC03、TC02、TC12、TC20五个红椿半同胞家系;胸径生长表现最好的为TC20、TC04、TC03、TC02、TC12五个红椿半同胞家系。
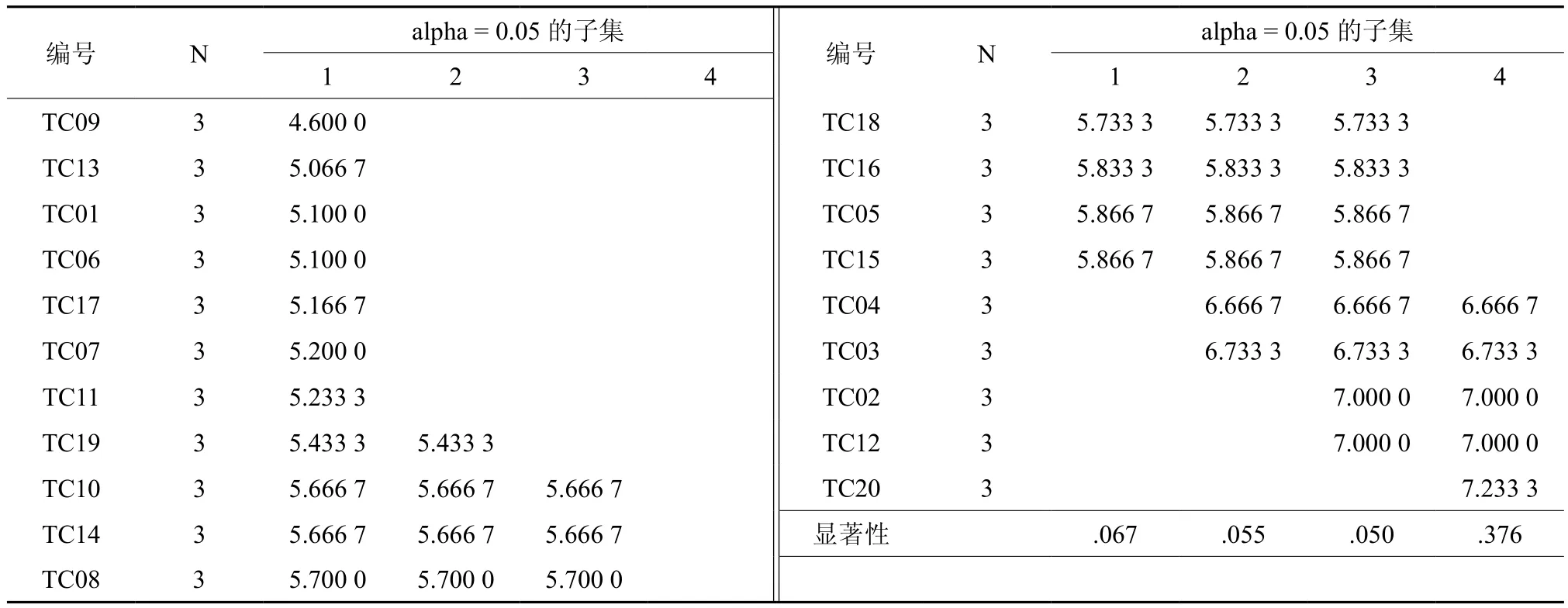
表4 5年生红椿半同胞家系测定林树高生长多重比较†Table 4 Multiple comparison of plant height of Toona ciliate half-sib families
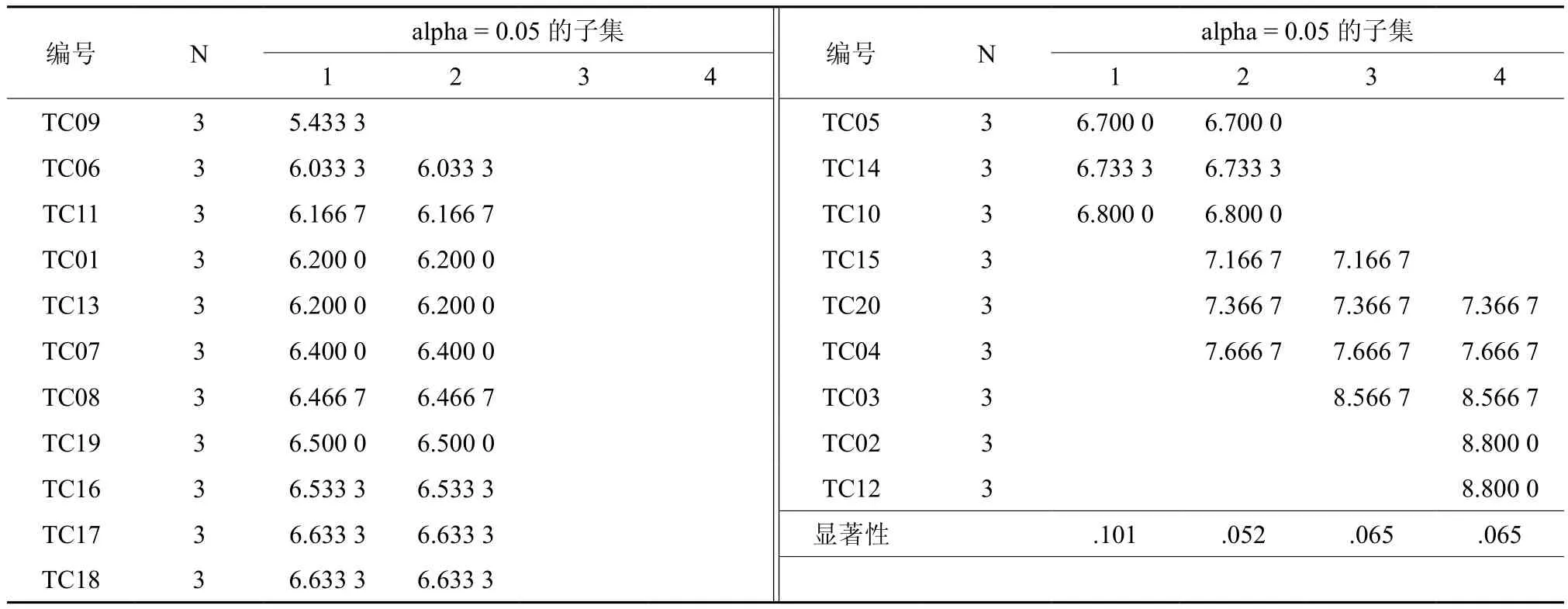
表5 5年生红椿半同胞家系测定林胸径生长多重比较†Table 5 Multiple comparison of DBH of Toona ciliate half-sib families
3 结 论
(1)5年生红椿半同胞家系测定林家系间树高和胸径的差异均达到极显著水平,表明各家系测定林的生长性状存在丰富的变异,具有很大的遗传改良的潜力。
(2)红椿半同胞家系树高和胸径的生长性状差异主要由遗传因素决定,红椿半同胞家系树高的遗传变异系数为11.03%,家系遗传力为0.7235,胸径的遗传变异系数为11.41%,家系遗传力为0.7306。
(3)以树高和胸径生长为指标,用Duncan多重比较(邓肯法)进行分析,筛选出树高和胸径生长表现最好的TC20、TC04、TC03、TC02、TC12等5个红椿半同胞家系。
[1]祁承经, 林亲众. 湖南树木志[M]. 长沙: 湖南科学技术出版社, 2000.
[2]廖德志, 吴际友. 两个优材更替树种[J]. 湖南林业, 2009, (7):24.
[3]刘福云, 鄢武先, 陈守庭, 等. 国内外珍贵树种用材林发展动态及其在四川省的发展现状[J]. 四川林业科技, 2009, 30(5):63-68.
[4]程冬生, 崔同林. 珍贵树种红椿的利用价值及培育技术[J].中国林副特产, 2010, (4): 39-40.
[5]张慧勤. 河南省珍贵用材树种现状、问题与发展对策[J]. 华东森林经理, 2008, 22(1): 15-17.
[6]刘 军, 陈益泰, 何贵平, 等. 毛红椿优树子代苗期性状遗传变异研究[J]. 江西农业大学学报, 2008, 30(1): 64-67.
[7]邹高顺. 珍贵速生树种红椿与毛红椿引种栽培研究[J]. 福建林学院学报, 1994, 14(3): 271-276.
[8]陈彩霞, 王瑞辉, 吴际友, 等. 持续干旱条件下红椿无性系幼苗的生理响应[J]. 中南林业科技大学学报, 2013,33(9):46-49.
[9]唐 强, 李志辉, 吴际友, 等. 毛红椿无性系扦插繁殖试验研究[J]. 中南林业科技大学学报, 2015, 35(1): 67-70.
[10]刘 军, 张海燕, 姜景民, 等. 毛红椿种实和苗期生长性状地理种源变异[J]. 南京林业大学学报, 2011, (3): 55-59.
[11]张 露, 郭联华, 杜天真, 等. 遮荫和土壤水分对毛红椿幼苗光合特性的影响[J]. 南京林业大学学报, 2006, (5): 63-66.
[12]刘 球, 陈彩霞, 吴际友, 等. 红椿无性系幼苗叶片抗氧化酶指标对干旱胁迫的响应[J]. 中南林业科技大学学报, 2013,33(11): 73-76.
[13]吴莉莉. 红椿树的生物学特性及人工栽培试验研究[J]. 安徽农学通报, 2006, 12(7): 168-169.
[14]吴际友, 李志辉, 刘 球, 等. 干旱胁迫对红椿无性系幼苗叶片相对含水量和叶绿素含量的影响[J]. 中国农学通报, 2013,29(4): 19-22.
[15]文卫华, 吴际友, 陈明皋, 等. 红椿优树子代苗期生长表现[J]. 中国农学通报, 2012, 28(34): 36-39.
[16]吴际友, 程 勇, 王旭军, 等. 红椿无性系嫩枝扦插繁殖试验[J]. 湖南林业科技, 2011, 38(4): 5-8.
[17]吴际友, 陈 瑞, 王旭军, 等. 毛红椿光合速率及生理生态因子的日变化规律[J]. 湖南林业科技, 2011, 38(2): 5-8.
[18]姜 芸, 吴际友, 姚贤清, 等. 湖南省珍贵乡土用材树种研究现状与发展对策[J]. 湖南林业科技, 2013, 40(3): 1-4.
[19]WU Ji-you, HUANG Ming-jun, CHEN Ming-gao. Cultivation Density of Toona ciliate[J]. Agricultural Science & Technology,2014, 15(10): 1730-1732.
[20]Wu Ji-you, Cheng Yong, WU Qi-jun. Study on Cutting Propagation of Toona ciliata Clones[J]. Agricultural Science &Technology, 2014, 15(11): 1844-1846.
[21]陈晓阳, 沈熙环. 林木育种学[M]. 北京: 高等教育出版社,2005.
Growth and early selection of half-sib families of Toona ciliata
WU Ji-you1,2, LI Yan3, LI Zhi-hui3, HUANG Ming-jun1,2, CHEN Ming-gao1,2, CHENG Yong1,2,
LIAO De-zhi1, LIU Qiu1,2, WU Qi-jun4
(1. Hunan Academy of Forestry, Changsha 410004, Hunan, China; 2. High Quality Timber Engineering Technology Research Center in Hunan Province, Changsha 410004, Hunan, China; 3. Central South University of Forestry and Technology, Changsha 410004, Hunan,China; 4. Jinlin of Hunan Forestry Co., LTD, Taoyuan 415701, Hunan, China)
The growth traits and heritability of 20 half-sib families test plantation of 5-year-oldToona ciliatewere studied. The results showed that: There were signi fi cant differences in tree height and DBH. In addition, there were high genetic improvement potential of tree height and DBH. Heritability for tree height and DBH of half-sib families was 0.723 5 and 0.730 6 respectively. Using Duncan to select the optimal families which were TC20, TC04, TC03, TC02, TC12.
Toona ciliate; half-sib families; heritability
S72
A
1673-923X(2016)04-0001-04
10.14067/j.cnki.1673-923x.2016.04.001
http: //qks.csuft.edu.cn
2014-12-22
湖南省林业科技计划项目“闽楠、红椿家具材良种选育与应用”(XLK201406)
吴际友,研究员,博士
李志辉,教授,博士生导师;E-mail:lzh1957@126.com
吴际友,李 艳,李志辉,等. 红椿半同胞家系生长与早期选择[J].中南林业科技大学学报,2016, 36(4): 1-4.
[本文编校:吴 彬]
猜你喜欢
广西林业科学(2022年6期)2023-01-16
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
养猪(2022年4期)2022-08-17
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
林业科技情报(2021年2期)2021-07-13
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
山东农业科学(2014年1期)2015-03-09
