
青秀山风景区优势种大叶栎种内与种间的竞争关系
2016-12-20 08:09张忠华李玉芳
中南林业科技大学学报 2016年1期
张忠华 ,胡 刚 ,秦 川 ,何 炎 ,李玉芳
(广西师范学院 a.环境与生命科学学院;b.北部湾环境演变与资源利用教育部重点实验室,广西 南宁 530001)
青秀山风景区优势种大叶栎种内与种间的竞争关系
张忠华a,b,胡 刚a,b,秦 川a,何 炎a,李玉芳a
(广西师范学院 a.环境与生命科学学院;b.北部湾环境演变与资源利用教育部重点实验室,广西 南宁 530001)
树木之间的竞争关系是塑造森林结构和动态的关键因素。采用Hegyi单木竞争指数,定量分析了青秀山次生林优势种大叶栎Castanopsis fi ssa种内与种间的竞争关系。结果表明:1)大叶栎种内竞争强度随着胸径的增大而逐渐减小,这与种内的密度制约性的自疏效应有关。2)大叶栎与乔木上层建群种马尾松Pinus massoniana的竞争最为强烈,其次为大叶栎、稠木Lithocarpus glabra、阴香Cinnamomum burmannii等树种。大叶栎种间竞争较种内竞争更为剧烈,竞争的结果可能会导致马尾松种群逐渐退出群落。3)大叶栎种内以及与整个林分的竞争强度与对象木胸径之间近似服从幂函数关系,而大叶栎与伴生树种的竞争强度与其胸径间近似服从对数函数关系,竞争强度和对象木胸径间呈显著的负相关关系。大叶栎种内和种间的竞争强度随大叶栎树龄的增大而减小,当其胸径超过30 cm后,种内和种间竞争强度的变化很小。
大叶栎;优势种;种内和种间竞争;竞争指数;青秀山风景区
物种竞争是同种或异种之间为争夺资源(光、养分、水分和生长空间等)而产生的一种直接或间接抑制对方的现象,竞争可能会导致一方取得优势而另一方受抑制甚至逐渐衰亡[1-2]。资源的获取和利用能力影响到植物种群的生存适合度,当邻体植物争夺有限资源时,会导致植物种群存活率和生长率的降低[3-4]。因此,在资源有限情况下,除了气候、土壤等环境因素外,植物种群的大小和群落的结构往往取决于种内和种间的生物调控因素(如,竞争、互惠关系等)[5]。植物种内和种间竞争是自然界的普遍现象,这种竞争关系是塑造森林林分结构和动态的关键驱动因素。因此,树木种内与种间的竞争关系及其影响因素一直是生态学研究的重要内容[1-5]。竞争在植物种群和群落的结构以及植被演进的动态过程中扮演着重要角色,研究植物的竞争关系有助于掌握森林结构和动态的演变趋势,揭示群落的发育与演替进程,同时对森林的可持续管理、林分的改造以及退化生态系统的恢复与重建具有重要指导意义。
次生林是由于人为破坏性干扰或异常自然干扰使原始林固有的林分结构、物种组成或基本功能发生了显著变化,经过天然更新或人工诱导天然更新恢复形成的林分[6]。次生林是我国森林资源的主体,是林产品的生产基地,在维护生态平衡、改善生态环境以及保护生物多样性等方面发挥重要作用。南亚热带次生林是我国华南地区的主要森林类型,其群落的建群种往往是由先锋树种组成,在物种组成、群落结构、生产力、林分环境以及相应的生态服务功能等与原始林有诸多差异,它的存在对改善生态环境和维系区域陆地生态平衡方面具有重要意义[7]。大叶栎Castanopsis fi ssa为壳斗科栎属常绿阔叶乔木树种,主要分布于我国广西、广东、福建、江西、湖南、贵州南部及云南东南部等地的山地疏林和阳坡等生境类型中[8]。大叶栎是广西的乡土树种,也是森林干扰后形成次生林的先锋树种,其自然更新能力强、生长快,分布广泛,是植被生态恢复与重建以及水土保持的优良树种,具有较大的开发利用潜力[9]。然而,迄今对大叶栎次生林的林分结构和动态等方面研究却鲜有报道。
在植物种内和种间竞争方面,国内外学者开展了丰富的研究工作。例如,Castagneri[3]和Das[4]等运用竞争指数分别探讨了意大利亚高山针叶林和美国加利福利亚针叶林内树木的竞争关系,发现竞争作用显著提高了森林树种的死亡率。我国学者采用Hegyi单木竞争指数对不同森林类型中的优势种或珍稀濒危树种,如黄果厚壳桂Cryptocarya concinna[10]、 青 冈 栎 Cyclobalanopsis glauca[11]、宝华玉兰Magnolia zenii[12]和太白红杉Larix chinensis[13]等树种进行了种内和种间竞争关系的研究,表明自疏和它疏过程在树木竞争中扮演着重要角色。但迄今为止,对大叶栎种内和种间竞争关系研究未见有文献报道。广西南宁的青秀山风景区分布有以大叶栎为优势种的典型南亚热带次生林,本文以青秀山大叶栎群落为研究对象,采用Hegyi单木竞争指数,分析大叶栎种内及其与伴生树种之间的竞争关系,旨在为南亚热带次生林的林分结构与动态以及植被恢复与重建研究提供理论参考,同时也为林业经营与科学管理提供依据。
1 研究地概况
青秀山风景区位于广西南宁市的东南郊(22°46′ N,108°33′ E),总面积约 22 km2,海拔83~289 m。该山地处亚热带的南缘,属湿润亚热带季风气候,阳光充足,雨量充沛,气候温和,夏长冬短,年平均气温为21.6 ℃,冬季最冷的1月平均气温为12.8 ℃,夏季最热的7、8月平均气温28.2℃,年均降水量达1 304.2 mm,平均相对湿度为79%。土壤主要是由砂岩和砂页岩发育而成的赤红壤,母岩主要是花岗岩,土壤类型属红粘土。青秀山拥有维管束植物达174科752属1 431种[14]。由于人为干扰与破坏,青秀山的原生植被已丧失殆尽,现除园林植被外,残存有以马尾松Pinus massoniana、大叶栎、稠木Lithocarpus glabra等为优势树种的常绿针阔混交次生林。该森林类型的郁闭度较高,植物种类较为丰富,形成了具有代表性的典型南亚热带森林景观。
大叶栎群落的垂直分层明显,乔木层通常可分为两个亚层,第一亚层以马尾松为主,高度16 m左右,个体分布较少,郁闭度为0.3~0.6,冠层不连续;第二亚层主要由大叶栎、稠木、阴香Cinnamomum burmannii、香叶树Lindera communis、木姜子Litsea pungens等组成,高度为8~12 m,冠层较连续;灌木层盖度为20%~50%,平均高度1.5 m,主要种类有九节Psychotria rubra、米碎花Eurya chinensis、三桠苦Melicope pteleifolia、栀子Gardenia jasminoides、鸭脚木Schef fl era heptaphylla、盐肤木Rhus chinensis等。此外,大叶栎、稠木、阴香等乔木树种的幼苗和幼树也较为常见;草本植物的种类较少,密度较低,盖度5%~30%,平均高度0.15 m,其主要种类有半边旗Pteris semipinnata、铁芒萁Dicranopteris linearis、山菅兰Dianella ensifolia、山麦冬Liriope spicata、薹草属Carex spp.等。层间植物种类稀少。
2 研究方法
2.1 野外调查
在青秀山风景区人为干扰相对较少的次生林内,随机选取胸径(离地1.3 m处)>5 cm的大叶栎为对象木,测定每株对象木的胸径、树高、冠幅、生长状态等指标。同时,以每株大叶栎对象木为中心,测定半径6 m样圆内所有乔木树种(竞争木)的种名、胸径、与对象木的间距、树高、冠幅、生长状态等指标。此外,记录林分的乔木层、灌木层和草本层的种类组成以及坡度、坡向和地被物厚度等生境特征。
2.2 数据处理
树木竞争模型种类较多,其中Hegyi单木竞争指数应用较为广泛且预测效果较好[10-13]。Hegyi单木竞争指数在形式上反映的是林木个体生长与生存空间的关系,实质反映了林木对环境质量的需求与现实生境下林木对环境资源占有量之间的关系[10]。该竞争指数的计测方法[15]如下:
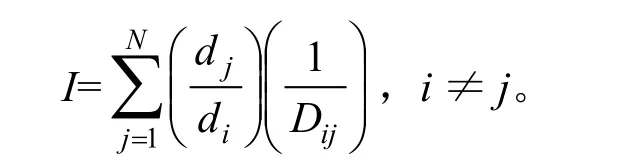
式中:I为竞争指数,其值越大,表明树种之间的竞争越剧烈;di为对象木i的胸径;dj为竞争木j的胸径;Dij为对象木i与竞争木j之间的距离;N为竞争木株数。
3 结果与分析
3.1 对象木与竞争木特征
在青秀山风景区,共随机调查大叶栎对象木30株,最小胸径6.8 cm,最大胸径54.1 cm,平均胸径22.9 cm。将大叶栎对象木按径级分组,其胸径集中分布在5~35 m之间,占对象木总数的86.7%,表明林分中大叶栎的幼树和小树较多,种群的更新良好(见表1)。由表2可知,总共测得竞争木18种949株,其中大叶栎366株,占竞争木总数的38.6%,其次为稠木、马尾松、香叶树和木姜子,分别占总竞争木个体数的 29.5%、11.4%、5.1%和4.5%。每株对象木在半径6 m、面积113.04 m2的样圆内平均拥有竞争木6.67种31.6株。
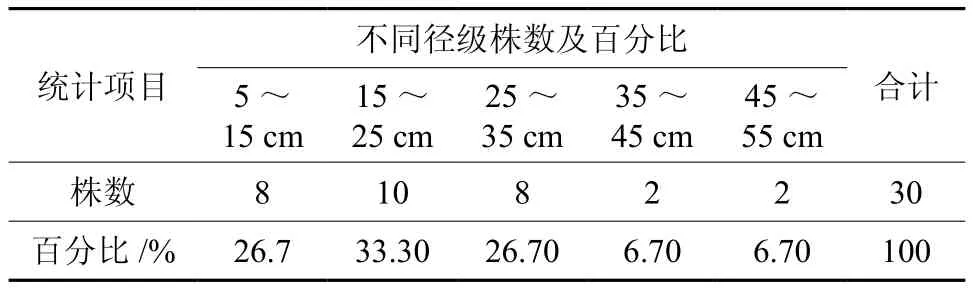
表1 对象木的胸径分布Table 1 DBH distribution of subject trees of C. fissa
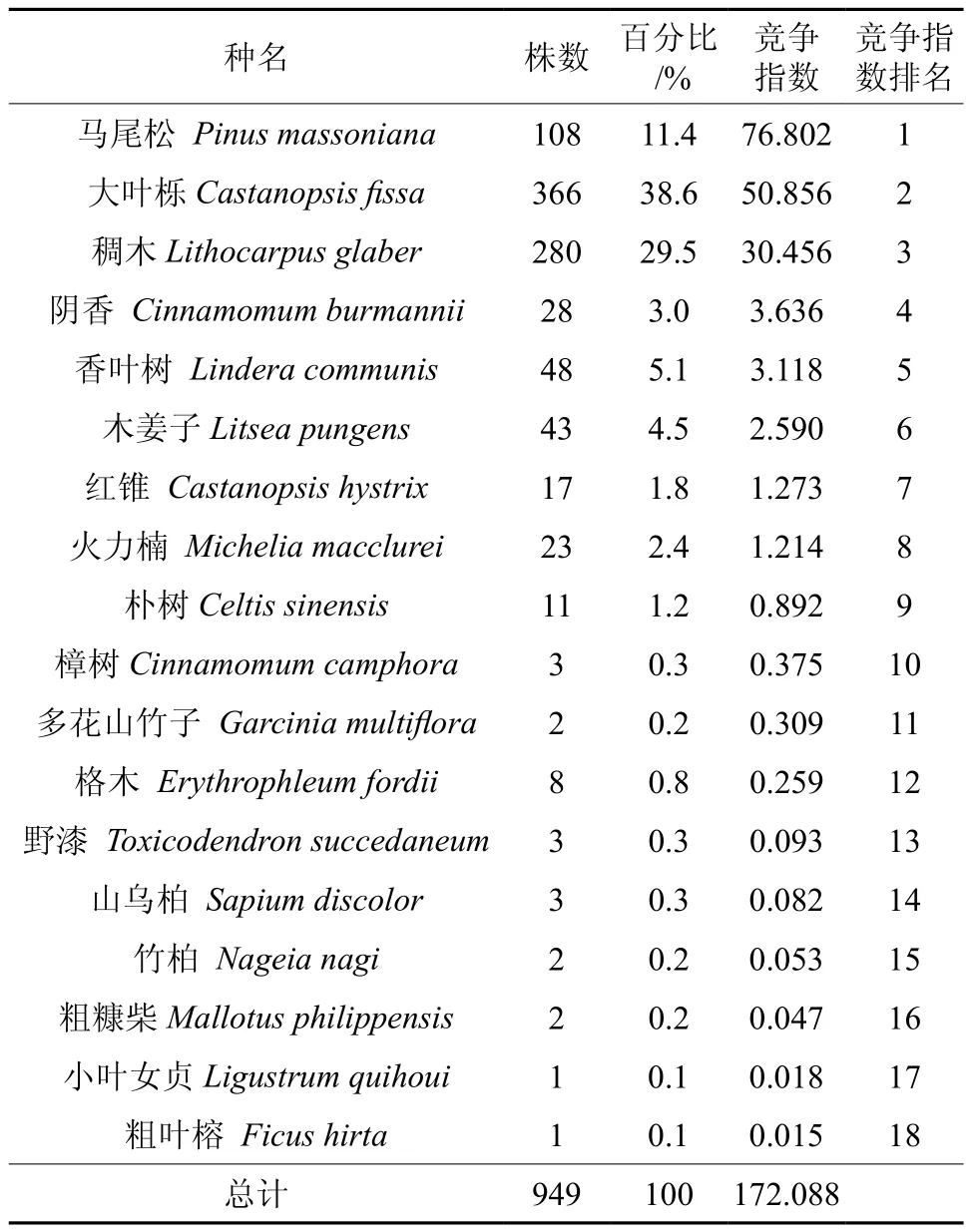
表2 竞争木的种类组成和竞争指数Table 2 Composition of competitive trees and competitive strength between C. fissa and competitive trees
3.2 大叶栎种内与种间的竞争关系
由表3可看出,随着对象木胸径的增大,种内竞争强度逐渐趋于减小。青秀山的大叶栎种群更新良好,幼树和小树较多,但周围的同种竞争木对其产生剧烈竞争。因此,幼树和小树阶段,大叶栎的种内竞争最强,这主要是密度制约效应而产生自疏效应的结果[16]。伴随树龄的增加,自疏导致种群密度逐渐降低,植株间的距离逐步加大,由于自疏调节作用使得个体间对水热、土壤养分、空间等资源的竞争性利用逐渐减弱,因而其种内竞争强度随个体的增大而趋于减小。
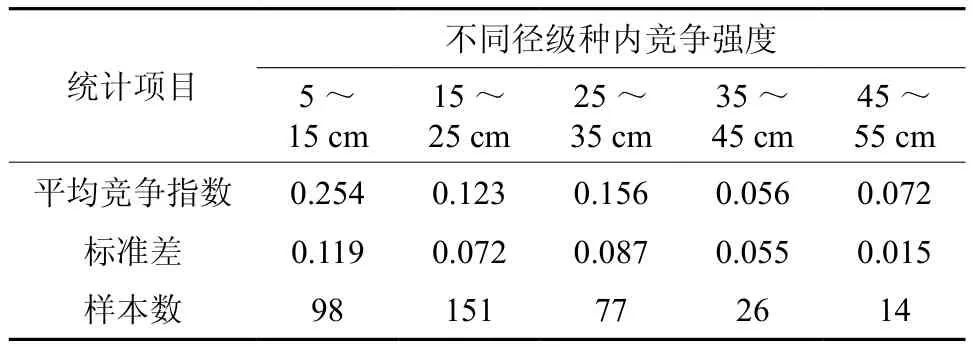
表3 大叶栎种内竞争强度Table 3 Intraspecific competition intensity of C. fissa
在大叶栎生长过程中,除了与同种发生竞争性的自疏效应外,还与异种植物争夺水分、养分和空间等资源,从而发生它疏效应。从表2 可看出,不同竞争木对大叶栎的竞争强度存在差异,其中马尾松与大叶栎的单木竞争指数值最大(76.802),其次为大叶栎(50.856)和稠木(30.456)。其余树种,如阴香、香叶树、木姜子以及后续的12种树种的竞争指数相差不大,表明其与大叶栎的竞争强度相对较弱。粗叶榕是众多竞争林木中与大叶栎竞争强度最小的树种(0.015),所有伴生树种的竞争指数总和为172.088。大叶栎种内和种间的竞争强度顺序依次为:马尾松>大叶栎>稠木>阴香>香叶树>木姜子>红锥>火力楠(其余竞争指数较小的10种略)。
3.3 竞争强度与对象木胸径的关系
树木竞争能力的强弱往往与个体大小密切相关[17]。将大叶栎对象木所受到的竞争强度与其胸径进行回归分析(见表4,图1),表明整个林分以及大叶栎种内对象木的竞争强度与对象木胸径之间的关系近似服从幂函数关系(I=AD-B,其中I为竞争指数,D为对象木胸径,A、B为模型参数),而伴生树种和对象木间的竞争强度与对象木胸径近似服从对数函数关系(I=A+BlnD)。经显著性检验,相关系数(r)均达到显著水平。这一结果表明,大叶栎种内、种间以及整个林分的竞争强度随着对象木树龄的增大而减小,当大叶栎胸径超过30 cm后,大叶栎种内以及与伴生树种之间的竞争强度变化很小。大叶栎的树龄越大,其周围生长的其它树木(竞争木)越少,致使竞争强度减弱。
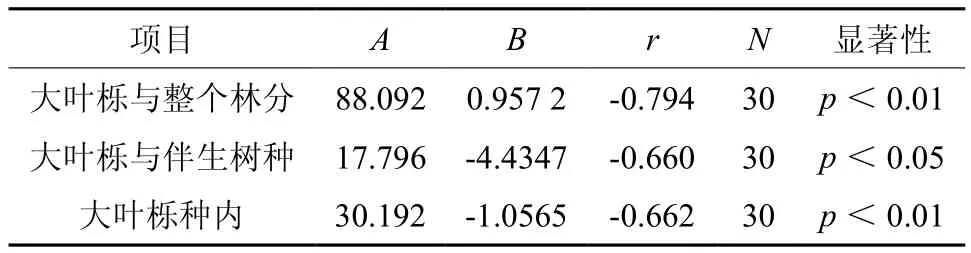
表4 竞争强度与对象木的胸径模型参数Table 4 Model parameters of competition intensity and DBH of subject trees
4 讨 论
本研究运用Hegyi单木竞争指数较好地量化了青秀山南亚热带次生林优势种大叶栎的种内和种间竞争强度。物种的竞争能力主要决定于该种的生物和生态习性以及物种的生态位情况[18-19]。生物和生态习性越接近的树种,往往占据相似的资源生态位,种间对有限资源的竞争可能会越剧烈,这种剧烈竞争的结果会导致部分种群降低其生存的空间[20]。张池等[10]采用Hegyi单木竞争指数对广东鼎湖山南亚热带成熟林中的黄果厚壳桂Cryptocarya concinna种内和种间竞争强度进行了分析,表明黄果厚壳桂种间竞争作用较种内竞争强,其竞争强度随林木径级的增大而逐渐减小。本研究同样表明,大叶栎与乔木上层树种马尾松的竞争最为强烈,其次为大叶栎种内、稠木、阴香等树种,表明这些树种与大叶栎之间对共同资源的竞争较为激烈,林分的它疏作用要大于自疏作用。此外,大叶栎幼树受到的竞争强度较大,在竞争中易遭到淘汰,而伴随大叶栎的生长,竞争木与大叶栎的竞争强度逐渐降低。
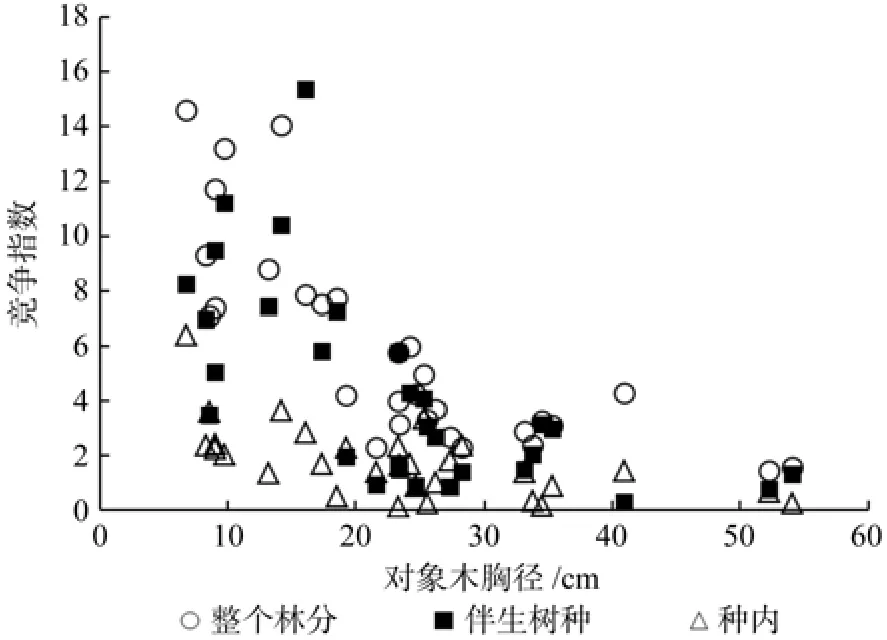
图1 大叶栎个体胸径与整个林分、伴生树种及种内竞争指数的关系Fig. 1 Relationships between DBH of objective tree and competition index of entire stand,associated and intraspeci fi c trees in C. fi ssacommunity
建群种或优势种在植物群落中占有主导地位,其竞争能力相较于其他伴生树种往往更强[11,13]。在大叶栎次生林中,马尾松为乔木上层优势种之一,大叶栎处于乔木中层,为林内少数几个优势常绿阔叶树种之一。在次生演替过程中,受到林下环境改变(如,光环境的改变)等增补制约因素的影响,马尾松几乎无幼苗和幼树,因此,其种群更新较差。在种间竞争作用下,特别是与大叶栎之间的剧烈竞争,加速了马尾松种群的衰退进程。然而,大叶栎的幼苗、幼树和小树个体数较多,其优势地位逐渐更为突出,属增长型种群。因而,在次生演替的现阶段,大叶栎与马尾松的种间竞争要比大叶栎种内竞争更为剧烈,伴随次生林的正向演替,可以预见,大叶栎将会逐步取代马尾松成为森林的建群种。大叶栎的种子主要依靠重力作用进行传播,因而大叶栎母树周围的幼苗和幼树较多。一般情况下,以母株为中心的繁殖方式使幼树分布较为集中,种群单位密度较大,但随着个体的生长,其对环境资源,如光、水、营养元素和空间等的需求也随之加剧,导致大叶栎种内相互作用增强而造成竞争性密度制约效应[21]。因此,除了与马尾松间的剧烈竞争外,大叶栎种内竞争也较为明显。
竞争指数与对象木胸径存在显著的负相关关系,即胸径大的个体,其所受的竞争强度小,反之则大[17,20]。对于小径级的大叶栎个体来说,因为个体小,处于林分的中下层,受邻近竞争木的影响,其所获取的生存空间和光照等资源有限,死亡率较高。随着林分的发育,在竞争过程中树木密度逐渐降低(自疏或它疏过程),存活下来的大叶栎以及其他树种均占据了相对合适的生态位。大叶栎的个体越大,其附近一定范围内生长的其它树木相对减少,林木因密度制约性的自疏或它疏作用而加大距离,因此对资源的竞争强度降低,树木分布渐渐趋于均匀化。因而,大叶栎对象木与同种或异种之间的竞争强度反而逐渐减弱,当大叶栎的胸径超过30 cm后,种内和种间的竞争强度变化不大。成熟的大叶栎母树与周围树种竞争强度的降低有利于幼树或小树获得足够的生存资源,竞争强度的动态变化是植物种群的一种适应机制或生存策略。
处于演替过程中的次生林相较于顶极阶段的原始林,林木之间的竞争格局与过程往往更为复杂和剧烈[7]。随着演替的进展,树木种内和种间竞争是一个动态的过程,竞争强度不断激化。森林群落中优势种的更替过程主要通过竞争实现,各种群的竞争强度越大,受竞争影响越大,竞争作用是次生林群落恢复演替的驱动力之一[3,22]。传统的竞争排斥理论认为,如果不出现生态位分化,完全相同的两个竞争物种不能稳定共存于同一生境中[23]。青秀山的次生林为马尾松、大叶栎和稠木等优势种组成的针阔混交林,大叶栎和马尾松均是造林的先锋树种,但马尾松不耐蔽荫,成林后往往更新不良,在大叶栎的竞争作用下,它疏作用致使马尾松的优势地位逐步被大叶栎取代,可能会演替为以大叶栎为建群种的常绿阔叶林。然而,在次生演替中,物种组成及其动态变化是受到生物因素(如物种生物学特性以及种间相互作用)以及非生物因素(如植被历史和气候特征等)的共同影响[24-26]。因此,在考虑种内和种间的竞争关系基础上,需要进一步考量其他生物与非生物因素,才能更为清晰地揭示南亚热带次生林的树木的种内和种间关系以及演替动态。树木竞争在森林生态系统中是一种十分常见而又复杂的现象,对竞争规律的研究对于了解群落结构与功能,预测群落的发展动态,进而辅以必要的人工管理措施以促进森林生态系统功能的完善具有重要的理论和现实意义。
[1]李 博,陈家宽,沃金森. 植物竞争研究进展[J]. 植物学通报,1998,15 (4):18-29.
[2]CASTAGNERI D,VACCHIANO G,LINGUA E,et al. Analysis of intraspeci fi c competition in two subalpine Norway spruce(Picea abies (L.) Karst.) stands in Paneveggio (Trento,Italy)[J].Forest Ecology and Management,2008,255:651-659.
[3]GUARIGUATA M R,OSTERTAG R. Neotropical secondary forest succession: changes in structural and functional characteristics[J]. Forest Ecology and Management,2001,148:185-206.
[4]DASA A,BATTLESB J,STEPHENSONC N L,et al. The contribution of competition to tree mortality in old-growth coniferous forests[J]. Forest Ecology and Management,2011,261: 1203-1213.
[5]CHARLES D,CANHAM M J,PAPAIK M U,et al.Neighborhood analyses of canopy tree competition along environmental gradients in New England forests[J]. Ecological Applications,2006,16:540-554.
[6]朱教君,刘世荣. 次生林概念与生态干扰度[J]. 生态学杂志,2007,26(7):1085-1093.
[7]康 冰,刘世荣,温远光,等. 广西大青山南亚热带次生林演替过程的种群动态[J].植物生态学报,2006,30(6):931-940.
[8]中国科学院中国植物志编辑委员会. 中国植物志(第22卷)[M].北京: 科学出版社,1998: 21-22.
[9]梁瑞龙,侯远瑞,周全连. 大叶栎生长调查研究[J]. 广西林业科学,2006,35(1): 8-11.
[10]张 池,黄忠良,李 炯,等. 黄果厚壳桂种内与种间竞争的数量关系[J]. 应用生态学报,2006,17(1): 22-26.
[11]胡 刚,梁士楚,张忠华,等. 桂林岩溶石山青冈栎种内与种间竞争的数量关系[J]. 西北林学院学报,2007,22(5): 32-36.
[12]蒋国梅,孙 国,张光富,等. 濒危植物宝华玉兰种内与种间竞争[J]. 生态学杂志,2010,29(2): 201-206.
[13]段仁燕,王孝安. 太白红杉种内和种间竞争研究[J]. 植物生态学报,2005,29(2):242-250.
[14]济 东,庞 洁,和太平,等. 南宁市青秀山风景旅游区植物区系研究[J]. 安徽农业科学,2010,38(19): 10422-10425.
[15]HEGYI F. A simulation model for managing jack-pine stands[C]//FRIES J. Growth models for tree and stand simulation. Stockholm:Royal College of Forestry,1974: 74-90.
[16]李先琨,苏宗明,欧祖兰,等. 元宝山冷杉群落种内与种间竞争的数量关系[J]. 植物资源与环境学报,2002,11(1):20-24.
[17]SIMARD S W,ZIMONICK B J. Neighborhood size effects on mortality,growth and crown morphology of paper birch[J].Forest Ecology and Management,2005,214: 251-265.
[18]邹春静,徐文铎. 沙地云杉种内、种间竞争的研究[J]. 植物生态学报,1998,22 (3):269-274.
[19]廖秋林,沈守云,卜国华,等. 多花木兰人工生态恢复植物群落种群竞争及其生态位研究[J]. 中国农学通报,2012,28(31):71-74.
[20]刘 彤,李云灵,周志强,等. 天然东北红豆杉Taxus cuspidata种内和种间竞争[J]. 生态学报,2007,27(3):924-929.
[21]SIMARD S W,SACHS D L. Assessment of interspeci fi c competition using relative height and distance indices in an age sequence of serial interior cedar-hemlock forests in British Columbia[J]. Canadian Journal of Forest Research,2004,34:1228-1240.
[22]OLIVER C D,LARSON B C. Forest Stand Dynamics[M]. New York: McGraw Hill,1996.
[23]HARDIN G. The competitive exclusion principle[J]. Science,1960,131: 1292-1297.
[24]徐远杰,林敦梅,米湘成,等. 古田山不同干扰程度森林的群落恢复动态[J]. 生物多样性,2014,22(3): 358-365.
[25]李志洪,胡淑仪,李 伟,等. 粤东北马尾松-木荷混交林种内种间竞争强度分析[J].中南林业科技大学学报,2013,33(8):91-95.
[26]李建军,李际平,刘素青,等. 基于Hegyi改进模型的红树林空间结构竞争分析[J]. 中南林业科技大学学报,2010,30(12):23-27.
Intraspeci fi c and interspeci fi c competition of dominant tree Castanopsis fi ssa in Qingxiushan Scenic Spot,Guangxi,southern China
ZHANG Zhong-huaa,b,HU Ganga,b,QIN Chuana,HE Yana,LI Yu-fanga
(a. School of Environment and Life Sciences; b. Key Lab. of Beibu Gulf Environment Change and Resources Utilization Co-constructed by China Education Ministry and Guangxi Province,Guangxi Teachers Education University,Nanning 530001,Guangxi,China)
Competitive relation between forest trees is a key factor in shaping forest structure and dynamics evolution. By using Hegyi single-tree competition index model,the intraspecific and interspecific competition intensity of Castanopsis fissa in Qingxiushan Scenic Spot Guangxi,southern China were quantitatively analyzed. The results show that (1) the intensity of intraspeci fi c competition in C. fissa stand decreased gradually with increasing tree diameter class,this was related to self-thinning effect of intraspecific density-conditionality. (2) The intense completion occurred between C. fissa and Pinus massoniana,followed by tree species C.fi ssa,Lithocarpus glabra,Cinnamomum burmannii,et al; this resulted that the interspeci fi c competition was more intensive than the intraspeci fi c competition in C. fi ssa,and the P. massoniana population may gradually decline in community. (3) The relationships of the DBH (diameter at breast height) of objective trees and the competition intensity between competitive trees and objective trees in the whole forest and C. fi ssa population nearly conformed to power function relations,while that between other competitive trees and the objective C. fi ssa tree conformed to logarithm function; the DBH of the objective tree was signi fi cantly negative correlation with competition intensity. The results show that the larger the DBH of the objective trees,the smaller the competition intensity of the C. fi ssa population,and when the DBH of C. fi ssa reached 30 cm,the change in competition intensity was very small.
Castanopsis fi ssa; dominant species; intraspeci fi c and interspeci fi c competition; competition index; Qingxiushan scenic spot
S718.5;Q948
A
1673-923X(2016)01-0067-05
10.14067/j.cnki.1673-923x.2016.01.012
2014-06-20
国家自然科学基金项目(31300351);广西自然科学基金项目(2013GXNSFBA019085);广西教育厅项目(201204LX235);广西高校大学生创新创业训练计划立项项目
张忠华,副教授,博士,硕士生导师 通讯作者:胡 刚,博士,硕士生导师;E-mail:ahhugang@gmail.com
张忠华,胡 刚,秦 川,等. 青秀山风景区优势种大叶栎种内与种间的竞争关系[J].中南林业科技大学学报,2016,36(1): 67-71,85.
[本文编校:谢荣秀]
猜你喜欢
现代农村科技(2022年5期)2022-11-18
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
宝藏(2019年6期)2019-07-04
天然产物研究与开发(2018年10期)2018-11-06
蔬菜(2018年5期)2018-05-17
现代农业研究(2017年11期)2018-01-12
中国农业文摘-农业工程(2017年2期)2017-01-12
农业与技术(2016年6期)2016-05-14
杂草学报(2015年2期)2016-01-04
中医研究(2014年12期)2014-03-11
