
干旱胁迫对3种豆科灌木生物量分配和生理特性的影响
2016-12-20 08:09李冬琴曾鹏程陈桂葵黎华寿
中南林业科技大学学报 2016年1期
李冬琴,曾鹏程,陈桂葵,黎华寿
(华南农业大学 农业部华南热带农业环境重点实验室,广东 广州 510642)
干旱胁迫对3种豆科灌木生物量分配和生理特性的影响
李冬琴,曾鹏程,陈桂葵,黎华寿
(华南农业大学 农业部华南热带农业环境重点实验室,广东 广州 510642)
通过盆栽实验,研究了干旱胁迫对南方常用绿化与护坡豆科灌木山毛豆Tephrosia candida、望江南Cassia occidentalis 和猪屎豆Crotalaria pallida 生物量分配、水分利用效率和生理特性的影响。结果表明:随着干旱胁迫程度的加剧,3种灌木的生物量下降,根冠比增加;干旱胁迫可提高3种灌木的水分利用效率,其中山毛豆和望江南在中度干旱下效率最高,猪屎豆在轻度干旱下最高;随着干旱胁迫程度的增加,3种灌木的脯氨酸(Pro)含量持续上升,望江南和猪屎豆的超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量也逐渐增加,但山毛豆的SOD活性和MDA含量则呈现出先增加后降低的趋势;干旱胁迫降低了3种灌木的叶绿素含量,并影响其叶绿素荧光动力学参数,其中变化幅度最大的是猪屎豆;3种灌木的抗旱性从强到弱依次为望江南>山毛豆>猪屎豆。
豆科灌木;干旱胁迫;抗旱能力;生物量分配;生理特性
干旱胁迫是影响植物生长、发育及分布的重要环境因子之一。长期的严重干旱不仅限制了植物的生长、发育,甚至会导致植物死亡[1-3]。近年来,随着高等级公路建设的迅速发展和人们对生态环境问题认识的提高,边坡生态恢复技术越来越受到重视。但是在植被恢复过程中,往往忽视了物种的生态适宜性、植物群落演替、生物多样性和景观价值等生态学规律,引入的植物在公路边坡的适应性差,3~5 a甚至更长的时间内都难以恢复[4],导致边坡土壤变得越来越干瘠、板结。因此,正确选择合理的植物是边坡生态防护和植被恢复工程中的重要环节。
豆科植物能与根瘤菌共生固氮,可补充土壤中的氮素,提高土壤肥力,改善区域生态环境[5],常认为共生固氮菌较寄主植物具有更强的抗旱能力[6]。另外,灌木较草本植物具有更发达的根系和更高的生物量,固土护坡及绿化效果好[7]。因此豆科灌木在干旱胁迫或者植被严重退化区域常常作为首选物种[5]。猪屎豆Crotalaria pallida、山毛豆Tephrosia candida和望江南Cassia occidentalis 均具有景观植物、饲料作物和绿肥作物的多种用途,近年在南方地区的园林绿化和边坡防护都推广应用。关于豆科灌木干旱胁迫的研究已有许多报道[8-14],但对猪屎豆、山毛豆和望江南的抗旱性特征及其种间差异的研究鲜有报道。因此,本试验主要研究人工模拟干旱胁迫对这3种豆科灌木生物量分配、水分利用效率和生理特性的影响,比较它们之间的抗旱差异,为边坡绿化植物的选择提供理论依据。
1 材料与方法
1.1 实验材料
供试植物材料为种子直接播种所得的当年生植株。其中猪屎豆和山毛豆种子购自广州华玲绿轩公司,望江南种子采集于华南农业大学校园。供试土壤为工程边坡土壤,自然风干,然后用5 mm筛过筛备用。土壤主要理化特性如下:pH值为 6.6,全氮 2.10 g·kg-1,全钾 7.60 g·kg-1,全磷0.29 g·kg-1,有机质 0.32 g·kg-1,石砾 0.291%,田间持水量为6.61%。
1.2 实验设计
土壤混合均匀后装入口径25 cm、高35 cm的塑料盆,每盆5.0 kg(干质量),并加入N、P、K有效养分各15%的挪威复合肥4 g。
选择籽粒饱满的种子,用自来水泡12 h后用3%双氧水消毒30 min,经蒸馏水漂洗多次后,埋于干净湿润的河沙至露出白芽。选择出芽一致的种子进行播种,每盆8粒,播种深度约2.0 cm,然后统一浇水管理。种子发芽一个半月后进行匀苗,每盆选留生长良好并较一致的植株2株,同时称取所有移出植物的干质量并计算其平均值,作为植株的初始生物量,随后进行干旱胁迫处理。
试验共设对照、轻度干旱、中度干旱和严重干旱4个水分梯度,使其保持田间持水量分别为100%、75%、50%和25%。每个梯度3个重复,共12盆。为了去除水分通过土壤表面蒸发的影响,同时准备了另外12盆装有同样的土壤,以测定每个水分处理下的土壤表面水分蒸发的含量。在处理过程中,定期移动植物在温棚中的位置,使每个植物在温棚内呈随机排列,并在植物之间保持一定距离,使冠层间通风透光。试验期间(共80 d),大棚内平均温度35℃/12℃(d/n),相对湿度45%~80%。中午最大光合有效辐射大约为1 500 μmol·s-1m-2。
试验期间,采用称重法保持各处理的土壤湿度。每2天称重浇水一次,每次浇水时间为下午16:00~18:00,使得处理过程中尽量保持土壤水分为田间持水量的100%、75%、50%、25%,干旱胁迫条件下的实际土壤质量含水量分别维持在(32.15±1.3)%、(24.11±0.5)%、(16.08±0.5)%、(8.04±0.4)%这个范围。
1.3 测定方法
生物量测定采用称重法;叶绿素含量参照李合生[15]方法,用分光光度计进行测定;超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBA)光还原法,活性单位以抑制NBA光还原的50%为一个酶活性单位[16];脯氨酸含量的测定用3%磺基水杨酸提取,用酸性茚三酮比色法[16]进行测定;丙二醛(MDA)含量的测定采用硫代巴比妥酸(TBA)比色法[16];叶绿素荧光动力学参数采用OS-30P型叶绿素荧光仪直接测,测定前叶片暗适应 20 min。
1.4 数据分析
采用Excel 和SPSS 13.0 等统计软件进行相关数据分析及统计。
2 结果与分析
2.1 干旱胁迫对3种灌木生物量的影响
不同程度干旱胁迫下,3种灌木各个处理间地上部生物量和根系生物量都存在显著差异,其地上部生物量的积累随着干旱胁迫的加剧呈下降趋势,与对照相比,在严重干旱胁迫时山毛豆、望江南和猪屎豆地上部生物量的积累量分别减少了87.98%、86.07%和96.68%。望江南和猪屎豆根系生物量的积累随着干旱胁迫的增加呈减小趋势,与对照相比分别减少了73.39%和93.22%。但山毛豆根系生物量的积累随着干旱胁迫的加剧呈现先增加后减少的趋势,在轻度胁迫时根系生物量最大,与对照相比增加了21.26%,但在严重干旱时减少了83.33%(见表1)。
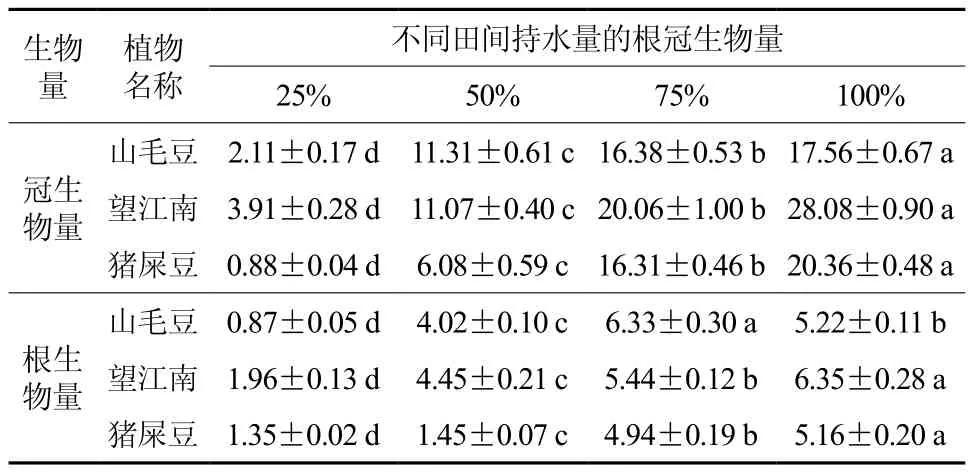
表1 干旱胁迫对3种灌木根冠生物量的影响 †Table 1 Effects of drought stress on aboveground biomass and root biomass of three kinds of shrubs g
2.2 干旱胁迫对3种灌木水分利用效率的影响
3种灌木的水分利用效率(WUE)随干旱胁迫加剧而呈增加趋势,田间持水量为25%、50%的水分利用效率显著大于75%、100%,其中山毛豆和望江南在50% 条件下最大,猪屎豆在75%条件下最大(见表2)。
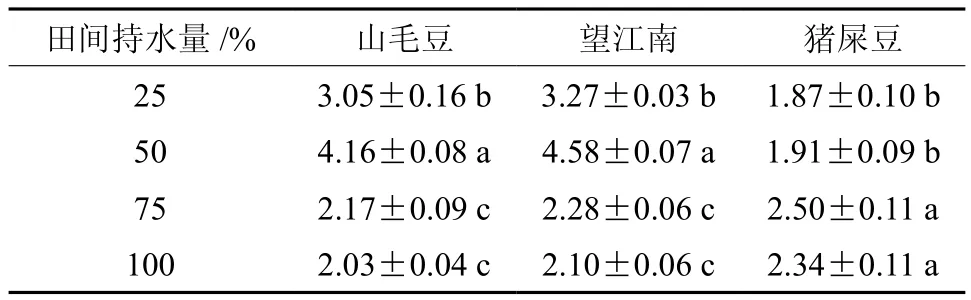
表2 干旱胁迫对3种灌木水分利用效率的影响Table 2 Effects of drought stress on water-use efficiency of three kinds of shrubs (g·kg-1)
2.3 干旱胁迫对3种灌木脯氨酸含量的影响
不同程度干旱胁迫下,3种灌木的脯氨酸(Pro)含量随胁迫程度的加重而逐渐增加,其中增加幅度最大的是望江南,在25%时显著大于对照,是对照的8.32倍,猪屎豆次之,增加幅度最小的是山毛豆,在25%时是对照的2.79倍(见表3)。
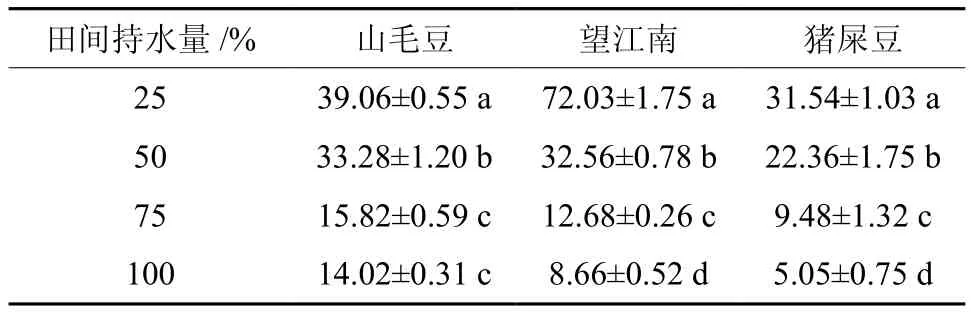
表3 干旱胁迫对3种灌木脯氨酸Pro含量的影响Table 3 Effects of drought stress on proline contents in three kinds of shrubs μg/g
2.4 干旱胁迫对3种灌木丙二醛含量的影响
在中度和重度干旱胁迫下, 3种植物体内的丙二醛(MDA)含量都显著增加。在田间持水量为75%时,望江南和猪屎豆的MDA与对照组无明显差异,但是山毛豆反而显著低于其对照组(见表4)。
2.5 干旱胁迫对3种灌木超氧化物歧化酶含量的影响
不同程度干旱胁迫下,望江南和猪屎豆体内产生的SOD含量随胁迫程度的加剧而逐渐增加,其中望江南的增幅最大,但在轻、中干旱胁迫下没有差异性;山毛豆产生的SOD呈先减少后增加的趋势,在严重干旱时达最高值,为341.14 U/mg(见表5)。

表5 干旱胁迫对3种灌木超氧化物歧化酶SOD含量的影响Table 5 Effects of drought stress on SOD contents in three kinds of shrubs U/mg
2.6 干旱胁迫对3种灌木叶绿荧光动力学参数的影响
随着干旱胁迫的加剧,山毛豆和望江南的基础荧光(Fo)先减少后上升,田间持水量为75%时,望江南的Fo显著低于其他处理,山毛豆各处理间无明显差异,而猪屎豆的F0随干旱胁迫的加剧逐渐上升,且各处理间差异显著。山毛豆的最大荧光(Fm)随干旱胁迫的加剧先上升后下降,且各处理间差异显著;猪屎豆的Fm在田间持水量75%条件下最大,其余处理间无明显差异;望江南的Fm在各处理间不显著。3种灌木的PSⅡ原初光能转化效率(Fv/Fm)随干旱胁迫加剧呈下降趋势,田间含水量为25%、75%的Fv/Fm显著低于75%、100%的,其中山毛豆和望江南在75%条件下最大(见表6)。
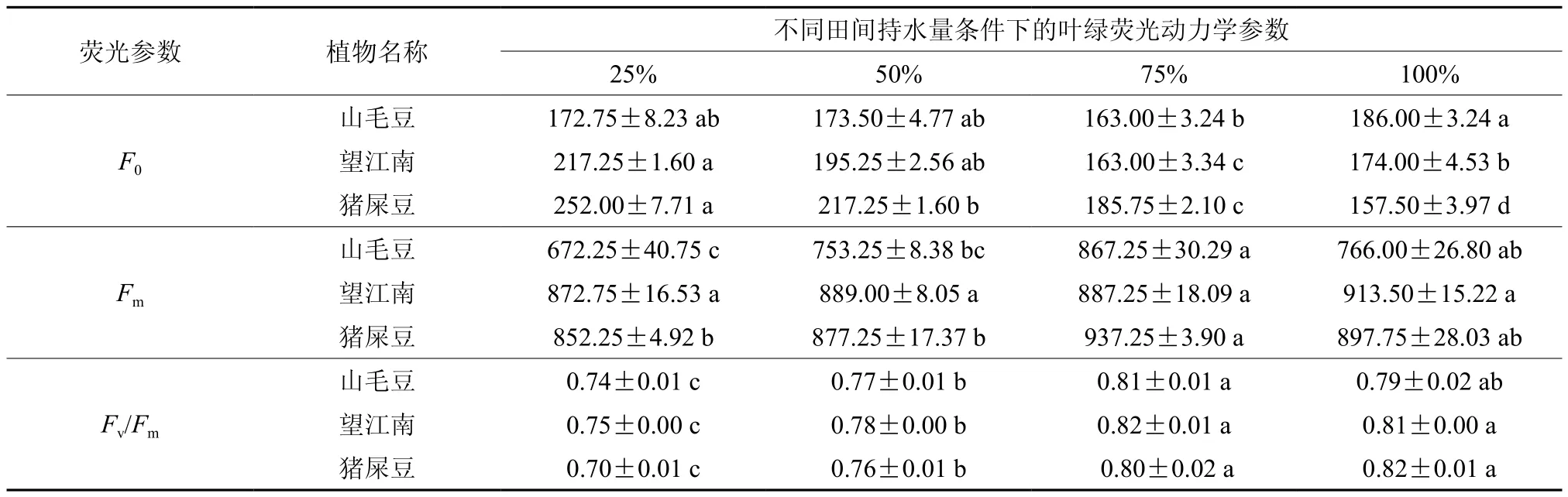
表6 干旱胁迫对3种灌木叶绿荧光动力学参数的影响Table 6 Effects of drought stress on chlorophyll fluorescence kinetics parameters in three kinds of shrubs
2.7 干旱胁迫对3种灌木叶绿素的影响
3种灌木的叶绿素含量在田间持水量为75%时与对照无明显差异。山毛豆的叶绿素α(Chlα)、叶绿素 b(Chlb)和总叶绿素 α+b(Chlα+b)的变化趋势相同,都是先增加后减少,在75%时,Chla、Chlb和Chla+b含量均达最高,且各处理间差异性显著;望江南的Chlα、Chlb和Chlα+b含量都是随干旱胁迫程度加剧而减少,在50%、25%时差异不显著,但显著低于100%、75%;猪屎豆的Chlα含量随干旱胁迫程度加剧而减少, Chlb和Chla+b含量则是先增加后减少,在75%时含量最高,50%、25%时差异显著(见表7)。
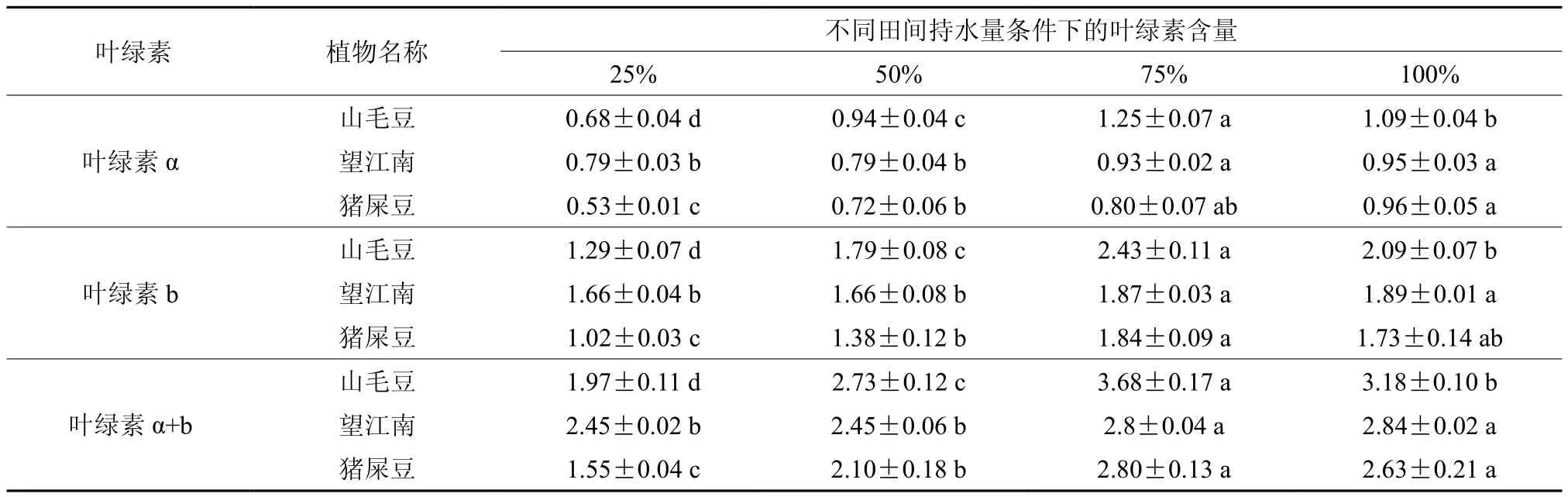
表7 干旱胁迫对3种灌木叶绿素含量的影响Table 7 Effects of drought stress on chlorophyll contents in three kinds of shrubs mg/g
3 讨 论
3.1 水分胁迫与生物量分配及水分利用效率的关系
不同环境条件下,生物量分配特性的变化可综合反映植物的适应能力。在干旱胁迫下,植物生物量通常都会减少,但根的减少幅度比地上部减少幅度相对较少[17-22],这说明植物可以通过提高根冠比来适应干旱胁迫。本试验研究也表明干旱胁迫可使3种豆科植物的根冠比增加,在对照、轻度、中度和严重干旱下,山毛豆的根冠比分别为0.30、0.39、0.36、0.41,望江南的分别为0.23、0.27、0.40、0.50,猪屎豆的分别为 0.25、0.30、0.24、1.53。其中在严重干旱时,3种灌木的根冠比最大,说明3种植物都可通过大幅降低地上部生物量来适应严重干旱。
水分利用效率用以表示植物产量与消耗水量的关系,也是评价植物生长适宜程度的综合生理生态指标。生物量和水分利用效率取决于土壤含水量的控制范围[23],一般而言,干旱胁迫(尤其是中等胁迫)可以提高水分利用效率[24-26],但有些物种水分利用效率降低或者不变[27-29]。当水分利用效率随着生物量与耗水量的减小而降低时,表明该植株具有浪费型水分利用对策[11]。本研究发现,望江南和山毛豆的水分利用效率随着干旱胁迫的加剧而上升,但在严重干旱时降低,而猪屎豆的水分利用效率从轻度胁迫时就开始下降;在生物量积累方面,望江南总生物量和水分利用效率远大于山毛豆和猪屎豆,其中,猪屎豆的生物量在干旱胁迫下下降幅度较大,望江南的生物量变幅较小,这说明猪屎豆可能具有浪费型水分利用对策,而望江南和山毛豆似乎具有节约型水分利用策略。
3.2 干旱胁迫与3种灌木生理特性的关系
脯氨酸(Pro)是植物体内最有效的一种亲和性渗透调节物质,具有很强的水溶性,它能与细胞内的一些化合物形成聚合物,具有一定的保水作用,从而抵御干旱胁迫。本研究中,随着干旱胁迫程度加剧,3种灌木脯氨酸含量持续增加,表明3种灌木都可以通过提高脯氨酸含量来维持自身的水分平衡,抵御干旱的影响。脯氨酸还具有抗氧化作用,能够清除一定的活性氧,并与丙二醛和膜透性呈显著相关[30],其含量的增加有利于提高植株的抗旱能力[31-32]。一般认为抗旱性强的植物渗透调节能力较强,且比抗旱性弱的植物脯氨酸累积量要多[33]。本研究发现,望江南增加幅度最大,山毛豆次之,猪屎豆最小,说明望江南的抗旱能力可能最强,猪屎豆最弱。
在干旱胁迫下,植物体内会产生大量活性氧,使膜脂过氧化而生成丙二醛。而SOD的主要功能是清除活性氧,防止活性氧对细胞膜进行伤害。有研究表明,干旱胁迫时植物体内的 SOD活性与抗旱性呈正相关关系[34-35],望江南和猪屎豆SOD活性有类似的结果,但山毛豆在轻度干旱时低于对照,表明山毛豆比较适合生长在轻度干旱的环境中。在严重干旱胁迫时,抗旱品种的花生叶片SOD活性增加程度明显大于敏感物种[36],且抗旱植物比不抗旱植物MDA 含量增加的幅度要小[33]。随着干旱胁迫程度加剧,望江南SOD活性增加程度明显大于山毛豆和猪屎豆,其中增幅最小的猪屎豆,在MDA含量增幅方面,猪屎豆最大,这进一步说明望江南的抗旱能力最强,而猪屎豆较弱。
基础荧光(Fo)表示PSⅡ反应中心全部开放即原初电子受体(QA)全部氧化时的荧光水平,常用于度量色素吸收的能量中以热和荧光形式散失的能量;最大荧光(Fm)是PSⅡ反应中心完全关闭时的荧光产量,反映了通过PSⅡ的电子传递情况;Fv/Fm比值代表PSⅡ原初光能转换效率。随着干旱胁迫的加剧,3种灌木Fo值整体呈上升趋势,Fm、Fv/Fm值呈下降趋势,这与许多研究结果类似,可作为植物抗旱性指标,参与抗旱品种的筛选[37-40],表明PSⅡ反应中心以热和荧光形式散失的能量增加,光合电子传递受阻和PSⅡ的原初光能转化效率降低。但山毛豆的Fo、Fm、Fv/Fm值变化趋势的拐点在田间含水量为75%时,即当干旱程度大于75%时,山毛豆Fo才开始上升,Fm、Fv/Fm才下降,说明该种植物最适宜在轻度水分胁迫环境中生长。望江南在中度、重度干旱胁迫下只有Fv/Fm显著低于轻度胁迫和对照,说明在中度、重度干旱胁迫下望江南PSⅡ的光能转化效率严重受损。猪屎豆在中、重干旱胁迫下Fo、Fm、Fv/Fm变化差异显著,且在3种植物中变化幅度最大,一般而言,耐旱植物叶绿素荧光参数值的变化幅度比不耐干旱植物小[33],所以猪屎豆的抗旱能力可能最弱。
叶绿素是绿色植物进行光合作用的主要物质,其含量在一定程度上能反映植物同化物质的能力。一般而言,植物在干旱胁迫下叶绿素含量的变化在一定程度上反映植物受害程度[41]。大多数研究表明,干旱胁迫降低了植物叶绿素含量[42-44],但抗旱性越强的植物降低幅度越小[45]。本实验的研究结果表明,在严重干旱情况下,叶绿素下降幅度最大的是猪屎豆,其次是山毛豆,降幅最小的是望江南。因此可以认为猪屎豆的抗旱性最弱,山毛豆的抗旱性中等,望江南的抗旱性最强。
4 结 论
植物的抗旱性受多种因素的共同作用,是一个复杂的调整机制,各个因子间有一定的关联。本实验中对3种豆科灌木生物量分配、水分利用效率和生理特性3个方面指标进行抗旱性评价,各指标所表达的植物抗旱性顺序基本一致,因此可以明确得出3种豆科灌木的抗旱能力从强到弱依次为:望江南>山毛豆>猪屎豆,望江南可作为南方地区边坡绿化和干旱地区景观绿化的首选豆科植物。
[1]Dias P C,Araujo W L,Moraes G A,et al. Morphological and physiological responses of two coffee progenies to soil water availability[J]. Journal of Plant Physiology,2007,164(12): 1639-1647.
[2]Guo W,Li B,Zhang X,et al. Architectural plasticity and growth responses of Hippophae rhamnoides and Caragana intermedia seedlings to simulated water stress[J]. Journal of Arid Environments,2007,69(3): 385-399.
[3]Rodiyati A,Arisoesilaningsih E,Isagi Y,et al. Responses of Cyperus brevifolius(Rottb.) Hassk. and Cyperus kyllingia Endl. to varying soil water availability[J]. Environmental and Experimental Botany,2005,53(3): 259-269.
[4]王仁德,钱金平. 生态脆弱区道路边坡生态恢复适用技术研究——以沙蔚铁路建设工程为例[J]. 水土保持研究,2007,14(3): 163-165.
[5]李芳兰. 三种豆科灌木对干旱胁迫的响应与适应[D].北京:中国科学院研究生院,2007.
[6]Zahran H H. Rhizobium-legume symbiosis and nitrogen fi xation under severe conditions and in an arid climate[J]. Microbiology and Molecular Biology Reviews,1999,63(4): 968-989.
[7]邓辅唐,喻正富,杨自全,等. 山毛豆、木豆、猪屎豆在高速公路边坡生态恢复工程中的应用[J].中国水土保持,2006,(4):21-23.
[8]Bargali K,Tewari A.Growth and water relation parameters in drought-stressed Coriaria nepalensis seedlings[J]. Journal of Arid Environments,2004,58(4): 505-512.
[9]Jurado E,García J F,Flores J,et al. Leguminous seedling establishment in Tamaulipan thornscrub of northeastern Mexico[J].Forest ecology and management,2006,221(1):133-139.
[10]Villagra P E,Cavagnaro J B. Water stress effects on the seedling growth of Prosopis argentina and Prosopis alpataco[J]. Journal of Arid Environments,2006,64(3): 390-400.
[11]李芳兰,包维楷,吴 宁. 白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J]. 生态学报, 2009,29(10): 5406-5416
[12]颜淑云,周志宇,邹丽娜,等. 干旱胁迫对紫穗槐幼苗生理生化特性的影响[J]. 干旱区研究,2011,28(1): 139-145.
[13]姜雪昊,穆立蔷,王晓春,等. 3 种护坡灌木对干旱胁迫的生理响应[J]. 草业科学,2013,30(05): 678-686.
[14]赵 洁,杜润峰,王龙飞,等. 达乌里胡枝子抗氧化防御系统对干旱及增强 UV-B 辐射的动态响应[J]. 草地学报,2013,21(2): 308-315.
[15]李合生. 植物生理生化实验原理与技术[M]. 北京: 高等教育出版社,2000.
[16]陈建勋,王晓峰. 植物生理学实验指导[M]. 广州: 华南理工大学出版社,2002.
[17]尉秋实,赵 明,李昌龙,等. 不同土壤水分胁迫下沙漠葳的生长及生物量的分配特征[J]. 生态学杂志,2006,25(1): 7-12.
[18]贺海波,李 彦. 干旱、 盐胁迫条件下两种盐生植物生物量分配对策的研究[J]. 干旱区研究,2008,25(2): 242-247.
[19]金不换,陈雅君,吴艳华,等. 早熟禾不同品种根系分布及生物量分配对干旱胁迫的响应[J]. 草地学报,2009,17(6):813-816.
[20]徐 飞,郭卫华,徐伟红,等. 刺槐幼苗形态、生物量分配和光合特性对水分胁迫的响应[J]. 北京林业大学学报,2010,(1):24-30.
[21]闫海霞,方路斌,黄大庄. 干旱胁迫对条墩桑生物量分配和光合特性的影响[J]. 应用生态学报,2012,22(12): 3365-3370.
[22]王晓冬,曹立萍,刘延迪. 干旱胁迫对真桦根系形态及生物量分配的影响[J]. 防护林科技,2011,(5): 20-22.
[23]Kang S,Zhang L,Liang Y,et al. Effects of limited irrigation on yield and water use ef fi ciency of winter wheat in the Loess Plateau of China[J]. Agricultural Water Management,2002,55(3): 203-216.
[24]Liu F,Andersen M N,Jacobsen S,et al. Stomatal control and water use ef fi ciency of soybean (Glycine max L. Merr.) during progressive soil drying[J]. Environmental and Experimental Botany,2005,54(1): 33-40.
[25]Maggio A,De Pascale S,Ruggiero C,et al. Physiological response of fi eld-grown cabbage to salinity and drought stress[J].European Journal of Agronomy,2005,23(1): 57-67.
[26]Bacelar E A,Moutinho-Pereira J M,Gonçalves B C,et al.Changes in growth,gas exchange,xylem hydraulic properties and water use ef fi ciency of three olive cultivars under contrasting water availability regimes[J]. Environmental and Experimental Botany,2007,60(2): 183-192.
[27]Hubick K T,Farquhar G D,Shorter R. Correlation between water-use ef fi ciency and carbon isotope discrimination in diverse peanut (Arachis) germplasm[J]. Functional Plant Biology,1986,13(6): 803-816.
[28]Anyia A O,Herzog H. Water-use efficiency,leaf area and leaf gas exchange of cowpeas under mid-season drought[J]. European Journal of Agronomy,2004,20(4): 327-339.
[29]Liu F,Stützel H. Biomass partitioning,specific leaf area,and water use efficiency of vegetable amaranth (Amaranthus spp.)in response to drought stress[J]. Scientia Horticulturae,2004,102(1): 15-27.
[30]蒋明义,郭绍川. 氧化胁迫下稻苗体内积累的脯氨酸的抗氧化作用[J]. 植物生理学报 ,1997,23(4):347-352.
[31]刘 涛,李 柱,安沙舟,等. 干旱胁迫对木地肤幼苗生理生化特性的影响[J]. 干旱区研究,2008,25(2): 231-235.
[32]姜 琴,应叶青,解楠楠,等. Ca/CaM 信使对毛竹抗旱生理的影响[J]. 江西农业大学学报,2012,34(4): 743-748.
[33]赵 琳,郎南军,温绍龙,等. 云南干热河谷4种植物抗旱机理的研究[J]. 西部林业科学,2006,35(2): 9-16.
[34]邵怡若,许建新,薛 立,等. 5 种绿化树种幼苗对干旱胁迫和复水的生理响应[J]. 生态科学,2013,(4): 420-428.
[35]Kholová J,Hash C T,Kočová M,et al. Does a terminal drought tolerance QTL contribute to differences in ROS scavenging enzymes and photosynthetic pigments in pearl millet exposed to drought[J]. Environmental and Experimental Botany,2011,71(1): 99-106.
[36]姜慧芳,任小平. 干旱胁迫对花生叶片 SOD 活性和蛋白质的影响[J]. 作物学报,2004,30(2): 169-174.
[37]蒲光兰,周兰英,胡学华,等. 干旱胁迫对金太阳杏叶绿素荧光动力学参数的影响[J]. 干旱地区农业研究,2005,23(3):44-48.
[38]胡学华,蒲光兰,肖千文,等. 水分胁迫下李树叶绿素荧光动力学特性研究[J]. 中国生态农业学报,2007,15(1):75-77.
[39]谭雪红,高艳鹏,郭小平,等. 五种高速公路边坡绿化植物的生理特性及抗旱性综合评价[J]. 生态学报,2012,32(16):5076-5086.
[40]Li G L,Wu H X,Sun Y Q,et al. Response of chlorophyll fl uorescence parameters to drought stress in sugar beet seedlings[J].Russian Journal of Plant Physiology,2013,60(3): 337-342.
[41]陈彩霞,王瑞辉,吴际友,等. 持续干旱条件下红椿无性系幼苗的生理响应[J]. 中南林业科技大学学报,2013,33(9):46-49.
[42]Gratani L,Varone L. Adaptive photosynthetic strategies of the Mediterranean maquis species according to their origin[J].Photosynthetica,2004,42(2): 551-558.
[43]惠竹梅,孙万金,张振文. 外源Ca2+对水分胁迫下酿酒葡萄黑比诺主要抗旱生理指标的影响 [J]. 西北农林科技大学学报:自然科学版,2007,35(9): 137-140.
[44]Bindi M,Bellesi S,Orlandini S,et al. In fl uence of water de fi cit stress on leaf area development and transpiration of Sangiovese grapevines grown in pots[J]. American Journal of Enology and Viticulture,2005,56(1): 68-72.
[45]聂华堂,陈竹生,计 玉. 水分胁迫下柑桔的生理变化与抗旱性的关系[J]. 中国农业科学,1991,24(4): 14-18.
Effects of drought stress on biomass distribution and physiological characteristics in three kinds of leguminous shrubs
LI Dong-qin,ZENG Peng-cheng,CHEN Gui-kui,LI Hua-shou
(Key Lab. of Agro-Environment in Tropical Zone of South China Supported by China Ministry of Agriculture,South China Agricultural University,Guangzhou 510642,Guangdong,China)
The potted experiments were conducted to investigate the biomass allocation,water-use efficiency and physiological responses of Tephrosia candida,Cassia occidentalis and Crotalaria pallida. The results show that the biomass of three kinds of shrubs decreased and root-shoot ratio increased with the drought degree aggravating; Drought stress could improve the water-use ef fi ciency of three kinds of shrubs,and the highest water use ef fi ciency in T. candida and C. ccidentalis appeared when they were subjected to moderate drought stress,but in C. pallida was at low drought stress; When the drought stress increased,the proline contents of the three shrub species increased gradually,the superoxide dismutase (SOD) activity and malondialdehyde (MDA) content of C. ccidentalis and C. pallida increased,but the SOD activity and MDA content of T. Candida increased fi rst and then reduced; Drought stress decreased chlorophyll contents of three kinds of shrubs,and their chlorophyll fl uorescence kinetics parameters were affected,of them C. pallida had the maximum changing range. The order of three species in drought resistance was: C. ccidentalis > T. candida > C. pallida.
leguminous shrubs; drought stress; drought resistance; biomass distribution; physiological characteristics
S718.43;Q142
A
1673-923X(2016)01-0033-07
10.14067/j.cnki.1673-923x.2016.01.006
2014-05-11
国家重点基础研究发展计划(973计划)项目(2011CB100400);广东省科技计划项目(0203-E12147)
李冬琴,硕士研究生 通讯作者:黎华寿,教授,博士,博士生导师;E-mail:lihuashou@scau.edu.cn
李冬琴,曾鹏程,陈桂葵,等. 干旱胁迫对3种豆科灌木生物量分配和生理特性的影[J].中南林业科技大学学报,2016,36(1): 33-39.
[本文编校:谢荣秀]
猜你喜欢
作物研究(2022年4期)2022-09-21
心声歌刊(2021年1期)2021-07-22
中国糖料(2021年3期)2021-07-13
种子(2020年11期)2020-12-05
语文周报·教研版(2020年4期)2020-03-23
时代邮刊(2019年18期)2019-12-17
学生天地·小学低年级版(2019年4期)2019-05-05
中老年健康(2018年3期)2018-04-11
散文诗(2017年18期)2018-01-31
小天使·五年级语数英综合(2016年1期)2016-02-23
