
思茅松天然林林分直径大小多样性及环境解释1)
2016-12-19 08:55:59李超闾妍宇胥辉徐婷婷张博魏安超孙雪莲熊河先石晓琳欧光龙
东北林业大学学报 2016年11期
李超 闾妍宇 胥辉 徐婷婷 张博 魏安超 孙雪莲 熊河先 石晓琳 欧光龙
(西南地区生物多样性保育国家林业局重点实验室(西南林业大学),昆明,650224)
思茅松天然林林分直径大小多样性及环境解释1)
李超 闾妍宇 胥辉 徐婷婷 张博 魏安超 孙雪莲 熊河先 石晓琳 欧光龙
(西南地区生物多样性保育国家林业局重点实验室(西南林业大学),昆明,650224)
以云南省普洱市45块思茅松天然林样地为研究对象,选用Shannon指数(H)、Simpson指数(D)和断面积Gini系数(G)对思茅松林分总体、思茅松和其他树种的林分直径大小多样性进行量化,并采用CCA直接梯度排序法分析林分直径大小多样性随气候因子、地形因子、土壤因子和林分因子的变化规律。研究表明:(1)样地总体和思茅松的Shannon指数(H)和Simpson指数(D)的变化比较一致,其他树种的Shannon指数(Ho)、Simpson指数(Do)和断面积Gini系数(Go)变动幅度均比其他6个指数大。(2)气候、地形、土壤和林分因子直接排序的第一排序轴分别解释58.6%、92.0%、64.4%和76.4%的信息量,较好地反映了林分直径大小多样性随环境因子的变化规律。(3)从环境因子与排序轴的相关性上看,气候因子中除年降水外的8个降水因子相关性在0.15以上,而温度因子与林分直径大小多样性的相关性不高;地形因子中海拔(Alt)与第一排序轴的相关性最高(-0.467 5);土壤因子中全磷(TP)、全钾(TK)与第一轴排序相关性均高于0.30;林分因子中林分优势高(Ht)与第一排序轴呈现-0.418 0的负相关。(4)当较高气温和中等降水,海拔和坡度中等,全磷(TP)、全氮(TN)、有机质(OM)质量分数和pH值中等时,林分优势高和林分平均高最小;林分密度指数中等时,思茅松天然林具有最大的Ht、Hs、Dt、Ds值,而其他树种的林分直径多样性指数以及3个断面积Gini系数的规律性不强。
林分直径多样性;气候因子;地形因子;土壤因子;林分因子;CCA排序;思茅松
With 45 plots of Simao Pine (Pinuskesiyavar.langbianensis) natural forests in Pu’er City of Yunnan Province, China, Shannon index (H), Simpson index (D) and Gini coefficient (G) for overall stand, Simao pine and other trees in plots were selected to quantify the stand diameter size diversity. Then, canonical correlation analysis (CCA) was used to analyze the change law of stand diameter size diversity along with climate factors, topographical factors, soil factors and stand factors. The result showed that: (1)Shannon index (H) and Simpson index (D) of overall stand and Simao pine had more consistent changes, while theHo,DoandGoof other trees had great fluctuation than others which reflect that Simao pine had more abundant size classes. (2)The first ordination axis of the climate, topography, soil and stand factors reflected 58.6%, 92.0%, 64.4% and 76.4% for the variation, respectively, and it had better explanation on the variation of diameter size diversity along with the environment factors. (3)It was vividly depicted on the correlation between environment factors and ordination axes, the correlation between eight precipitation factors (except Annual Precipitation) and ordination axes was more than 0.15, and the correlation coefficient were not significant between temperature factors and ordination axes. The altitude (Alt) had the highest correlation with the first axis which was -0.467 5; the correlation between total phosphorus (TP), total potassium (TK), and the first axis was more than 0.30, while the stand dominant height (Ht) was the most relevant (-0.418) stand factor. (4)Simao pine natural forest had the largestHt,Hs,DtandDswhen the stand was under higher temperature and moderate precipitation, moderate altitude and slope, moderate total phosphorus (TP), total nitrogen (TN), organic matter (OM) content and pH value of soil, and least stand dominant height, stand average height and medium stand density index. But the change ofHo,Doand three Gini coefficients were with irregularity.
林分结构多样性是反映林分结构状况的重要指标,它在一定程度上决定了林分的稳定性,并直接关系到森林生态系统功能的发挥[1],一些学者提出了物种多样性、林木格局多样性和林木大小多样性3个指标[2]描述结构多样性。林木大小多样性主要体现在树高、胸径、年龄和冠幅等结构变异,由于胸径在林分中测定简单、方便、准确,并且与其他因子具有密切的相关性[3-4],因此,直径常被用来描述林木大小多样性。国内外大量有关林分大小多样性指标的研究[5],描述林木直径大小多样性的有Shannon指数、Simpson指数、Margalef指数、断面积Gini指数等8个指标[6]。
森林与环境存在相互影响、相互作用。林木的生长是气候、地形、土壤和林分等多种因素共同作用的结果,林分的密度、海拔高度、坡度、经纬度、土壤、当地水热条件等对林分结构具有一定程度的影响[7]。已经有研究表明立地条件对与林分结构特别是直径结构具有一定的影响,陈育峰[8]认为森林对气候的响应是非线性的,气候微小改变都可能对森林群落的结构产生影响;范叶青等[9]通过分析得出海拔、坡度、坡向和坡位等地形因子对毛竹林林分结构有显著影响,其中海拔、坡度比坡位和坡向的影响更明显;舒树淼等[10]研究认为,立地条件对林木大小多样性存在直接和间接影响,对胸径多样性的直接和间接影响系数均为0.23。
数量分类和排序是研究植物群落生态关系的重要的数量方法[11],有效的排序有助于表达植物群落与周围环境的关系,典范相关分析(CCA)自应用于生态学多元梯度分析后,由于能同时结合多个环境因子,包含的信息量大,结果直观、效果好受到了普遍重视[12]。目前大量排序的研究应用于植物群落多样性研究方面,对于林分结构(林分直径大小多样性)与周围环境的关系研究很少。
思茅松(Pinuskesiyavar.langbianensis)是卡西亚松(Pinuskesiya)的地理变种,属松科(Pinaceae)松属(Pinus)的暖热性树种,是云南省重要的用材树种之一[13],具有重要的经济价值、森林生态服务功能和碳汇效益[14-15]。目前有关思茅松的研究大都集中于思茅松的生物量、碳储量、苗木培育、产脂等方面;在思茅松林分结构方面,欧光龙等[16-17]研究了思茅松林分树高和胸径结构的变化以及两者偏度和峰度与环境因子的关系,但是对于思茅松林分直径大小多样性及其变化的环境影响因素研究较少。
本研究在云南省普洱市45块思茅松天然林样地调查的基础上,选择Shannon指数、Simpson指数和断面积Gini系数3个指标,分别描述样地总体、思茅松及其他树种的林分直径大小多样性;并结合气候、地形、土壤和林分因子,采用CCA排序方法进行直接梯度排序,分析思茅松天然林林分直径大小多样性的影响因素,揭示思茅松天然林林分直径大小多样性变化规律。
1 研究区概况
研究区位于云南省普洱市,地处北纬22°2′~24°50′、东经99°9′~102°19′,北回归线横穿中部,总面积45 385 km2,海拔317~3 307 m。全市由于受到亚热带季风气候的影响,气温15.3~20.2 ℃,年无霜期315 d以上,年降水量1 100~2 780 mm,年均相对湿度79%。全市森林覆盖率超过67%,是云南省重点林区和林业产业基地。思茅松是当地的主要用材和造林树种。
本研究涉及墨江哈尼族自治县(墨江县)通关镇(SiteⅠ)、普洱市思茅区云仙乡(SiteⅡ)和澜沧拉祜族自治县(澜沧县)糯福乡(SiteⅢ)(见图1)。
图1 研究区位置图
2 研究方法
2.1 数据调查与收集
本研究以墨江县鱼塘镇、澜沧县糯福乡以及思茅区云仙乡为研究区域,每个研究区域分别选取15个思茅松天然林典型样地,共计45个。林分均为成熟林,样地面积为20 m×30 m,样地为思茅松纯林,有些样地伴生有其他树种,其他树种为青冈(Cyclobalanopsisglauca)、余甘子(Phyllanthusemblica)、水锦树(Wendlandiauvariifolia)、红木荷(Schimawallichii)等。对样地内乔木,进行每木检尺,采集样地土壤样品,并记录样地经纬度信息、地形因子(海拔、坡度、坡向)等样地基本信息,从环境气候网站(http://www.worldclim.org)获取研究区气候数据。
2.2 林分直径大小多样性指数
选择Shannon指数、Simpson指数和胸高断面积Gin系数作为林分直径大小多样性指标[18-20],利用R语言分别计算样地总体、思茅松和其他树种的多样性指数。


式中:H为林分直径大小Shannon指数;D为Simpson指数;G为断面积Gini系数;pi为每个径阶林木株数ni占总株数n的百分比,按2cm标准整化径阶并分级,林木胸径数据共分为十级,其中<10cm和>40cm的分别为第一和第十级,10~40cm之间每两个偶数径阶为一级,统计各径阶的林木株数ni(i=1,….S);ai为样地内第i株树的胸高断面积。
2.3 环境因子数据处理及CCA排序分析
将得到的研究区气候数据进行整理,选用年均温(b1)、气温年较差(b2最热月均温-最冷月均温)、等温性(b3=b2/b7)、季节性气温变化(b4)、最热月最高温(b5)、最冷月最低温(b6)、气温年较差(b7=b5-b6)、最湿季均温(b8)、最干季均温(b9)、最热季均温(b10)、最冷季均温(b11)、年降水(b12)、降水最大月降水(b13)、降水最小月降水(b14)、季节降水变异系数(b15)、最湿季降水(b16)、最干季降水(b17)、最热季降水(b18)、最冷季降水(b19)19个气候因子进行分析。选用实地测定的样地海拔(Alt)、坡度(SLO)和坡向(ASPD)数据构建地形因子数据矩阵。基于各样地每木检尺的数据,计算得到林分平均年龄(Age)、郁闭度(YBD)、林分平均高(Hm)、林分优势高(Ht)、林分密度指数(SDI)和立地指数(SI)等,构建林分因子数据矩阵。测定样地土壤常规8项指标,构建含土壤pH值(pH)、土壤有机质(OM)、全氮(TN)、全磷(TP)、全钾(TK)、水解性氮(HN)、有效磷(YP)和速效钾(SK)8项土壤指标构建土壤因子数据矩阵。
采用CANOCO软件分别对样地的气候、地形、土壤和林分因子与林分直径大小多样性指数进行CCA直接梯度排序分析,分析林分直径多样性指数与各环境因子关系。
3 结果分析
3.1 多样性指数变化趋势分析
由图2可知,样地总体林分直径大小Shannon指数(Ht)取值为1~2;思茅松林分直径大小Shannon指数(Hs)取值为1~2.2,其他树种林分直径大小Shannon指数(Ho)取值范围0~2;样地总体Simpson指数(Dt)、样地内思茅松的Simpson指数(Ds)的取值分别为0.4~0.9,样地内其它树种的Simpson指数(Do)的取值为0~0.8;总的断面积Gini系数(Gt)、样地内思茅松的断面积Gini系数(Gs)的取值分别为0.25~0.8,样地内其他树种的断面积Gini系数(Go)的取值为0~0.8。Ht与Hs的变化趋势基本一致,仅在3号样地处出现异同,Ho变化幅度比较大,数值明显小于Ht与Hs,同时在4号和35号样地处出现0值,表明样地内,除思茅松外,其他树种的径级并不丰富,其中两个样地内除思茅松外其他树种只有一个径级(见图2(a))。Simpson指数的分布同Shannon指数的趋势基本一致,同样在4号和35号样地内Do处出现0值(见图2(b))。对于断面积Gini系数(见图2(c)),Gt比Gs、Go大,说明样地内林木整体的胸高断面积分化程度大于样地内思茅松的胸高断面积的分化程度;Go的变动幅度依然很大,说明各样地内除思茅松以外,其他树种的胸高断面积的分化程度并不相同,但是总的来说,其分化程度相对于各样地整体和思茅松的胸高断面积分化程度小,即径阶比较集中。
3.2 多样性指数CCA排序
从表1中可以看出,通过分别采用气候、地形、土壤和林分因子与林分直径大小多样性指数的CCA排序分析,各类环境因子的CCA排序结果较好,气候、地形、土壤和林分因子排序的第一排序轴分别解释了直径大小多样性变化信息58.6%、92.0%、64.4%和76.4%。前两个排序轴也分别累积解释其变化的81.1%、97.3%、88.5%和95.4%。可见排序的前两轴,尤其是第一轴较好的反映了林分直径大小多样性指数随各类环境因子的变化;且从排序解释的信息量看,地形因子解释了最高的直径大小多样性变化信息,林分因子次之,气候因子最小。

图2 样地林木大小多样性指数折线图

环境因子参数AX1AX2AX3气候CPVSD6.18.4 9.6CPVSECo58.681.192.1地形CPVSD16.617.618.1CPVSECo92.097.3100.0土壤CPVSD16.522.624.7CPVSECo64.488.596.7林分CPVSD22.327.828.7CPVSECo76.495.498.4
注:CPVSD多样性指数信息百分比,CPVSECo表示多样性指数与环境关系的累计解释量;AX1、AX2、AX3分别表示第一、二、三排序轴。
3.2.1 多样性指数与气候因子CCA排序
由表2可知,19个气候因子中,年降水量(b12)和最热季降水量(b18)与第一轴的相关性较高,其中前者为最大负相关(-0.062 3),后者为最大正相关(0.063 4);第二轴中,b13、b16、b17、b18、b19等降水因子的相关性均大于0.2。
根据前两轴绘制二维排序图(见图3(a)),第一轴从左至右,研究区等温性和年降水量逐渐降低,最冷月最低温逐渐变大;沿着第二轴从下到上,温度的季节变化、降水量最大月、降水量最小月、季节降水变异系数、最湿季降水、最干季降水和最冷最热季降水逐渐降低,最冷月最低温和等温性呈现增加的趋势。Ht、Dt、Hs、Ds与最冷月最低温和等温性具有一定的相关性,即两者较大并且降水因子最小的条件下,4个指数达到最大值,同时4个指数在排序图上位置很近,表明林分直径大小多样性与思茅松直径大小多样性变化趋势相似。Gt在最大月降水量和最湿季降水呈中等水平时达到最大值,而Gs在最大月降水和最湿季降水量最大时达到最大。Ho、Do和Go并没有体现出类似的规律性。
3.2.2 多样性指数与地形因子CCA排序
从表3可知,3个地形因子与CCA第一轴均呈现负相关,且海拔(Alt)为最大负相关(-0.467 5),而坡度与第二轴呈现最大的正相关(0.306 6),同时前两轴的解释量为97.3%(见表1),说明第一轴与第二轴很好的拟合了海拔、坡度与林分直径多样性指数的关系。根据前两轴绘制的二维排序图(见图3(b)),第一轴从左至右,3个因子均呈下降趋势,第二轴从下往上,坡向下降、坡度不断升高;Ht、Dt、Hs和Ds聚集在一起,表明样地直径大小多样性与思茅松的直径大小多样性变化趋势相似,它们均在相似的环境下取得最大值(即海拔、坡度偏小以及最小坡向的条件下);Gt和Gs受坡向影响较大。
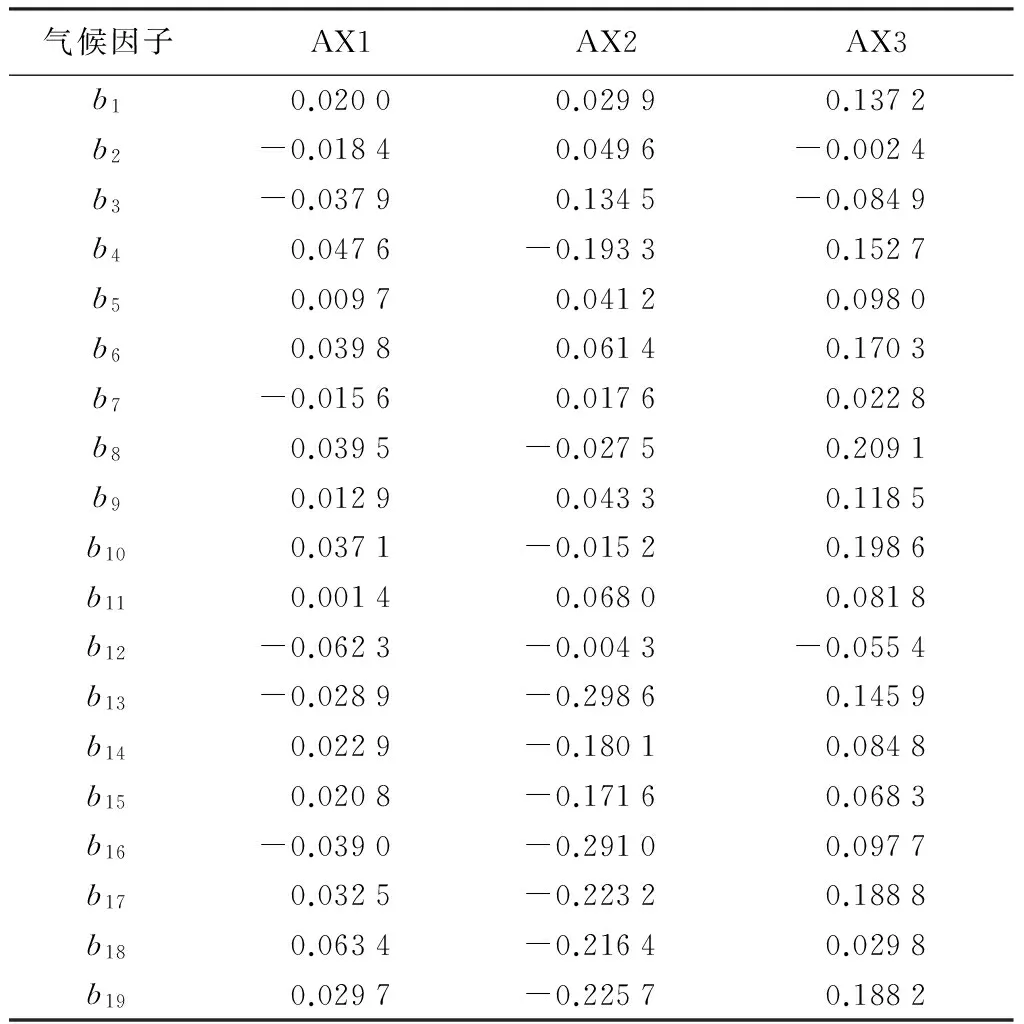
表2 CCA排序轴气候因子与排序轴关联系数
注:AX1、AX2、AX3分别表示第一、二、三排序轴。

表3 CCA排序轴地形因子与排序轴关联系数
注:AX1、AX2、AX3分别表示第一、二、三排序轴。
3.2.3 多样性指数与土壤因子CCA排序
从表4可知,与CCA第一轴存在最大相关性的土壤因子是全磷(TP),达到-0.330 1,其次是全钾(TK)为0.318 8,说明第一排序轴反映了林分直径大小多样性指数随土壤全磷和全钾的变化;与CCA第二轴存在最大相关性的土壤因子是全磷(TP),达到-0.400 4,此外,土壤速效钾和全氮相关系数也较高,均在0.30以上,且均呈负相关。可见,思茅松林分直径大小多样性指数主要受到全磷、全氮、速效钾的影响。
根据前两轴绘制二维排序图(见图3(c)),沿CCA第一轴从左至右,全磷的含量逐渐降低,全钾含量逐渐升高;沿着CCA第二轴从下到上,全磷、全氮和土壤有机质等土壤养分含量逐渐降低,土壤pH值逐渐增大。Ht、Dt、Hs和Ds聚集在一起,表明样地直径大小多样性与思茅松直径大小多样性变化趋势相似,它们均在相似的环境下取得最大值(即:中等条件的全磷、全氮、有机质、钾和最小pH值的土壤条件下);Ho和Do受全钾和速效钾的影响较大;3个断面积Gini系数都受土壤pH值的影响比较大。
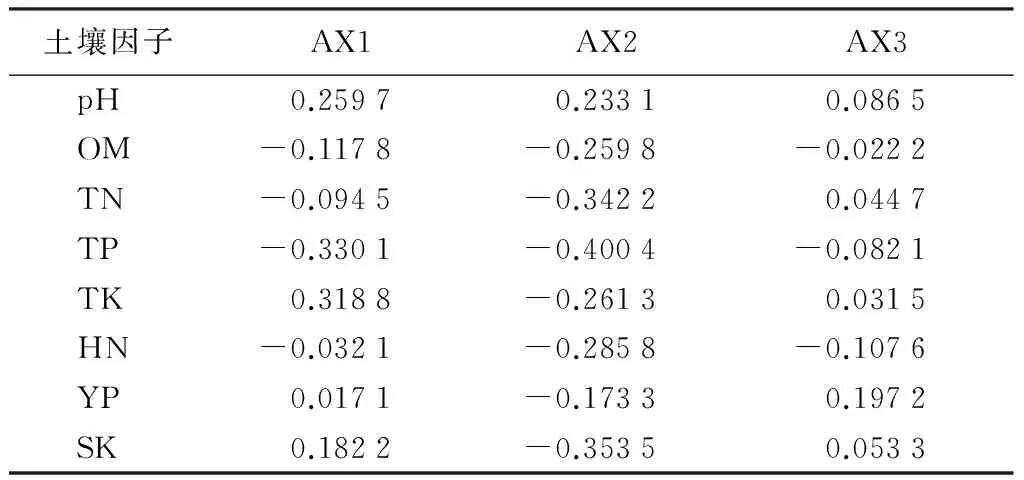
表4 CCA排序轴土壤因子与排序轴关联系数
注:AX1、AX2、AX3分别表示第一、二、三排序轴。
3.2.4 多样性指数与林分因子CCA排序
从表5可知,林分优势高与排序轴第一轴具有最大相关性,为-0.418 0,林分平均高次之,除林分密度指数外,其余林分因子均与第一轴均呈现负相关;林分平均高与第二轴呈现最大的相关性,为-0.439 1;而林分年龄与第三轴有最大的负相关。可见,对林木直径多样性指数影响比较大林分因子有林分平均高、林分优势高和林分年龄。

表5 CCA排序轴土壤因子与排序轴关联系数表
注:AX1、AX2、AX3分别表示第一、二、三排序轴。
根据前两轴绘制的二维排序图(见图3(d))可以看出,由第一轴从左至右,郁闭度、林分优势高和林分平均高等因子逐渐降低,林分密度指数逐渐增加;第二轴从下往上,随着林分密度指数、林分优势高、林分平均高等逐渐下降,郁闭度有上升的趋势。Ht、Dt、Hs和Ds聚集在一起,说明林分直径大小多样性与思茅松直径大小多样性变化趋势相似,它们在相似的林分条件下取得最大值,即林分优势高、林分平均高最小,林分密度指数中等的林分条件;3个断面积Gini系数并没有体现出类似的规律性。
4 结论与讨论
以Shannon指数、Simpson指数和断面积Gini系数,分别分析45个思茅松天然林样地的直径大小多样性,同时借用生态学中的梯度排序的方法分析气候、地形、土壤和林分因子对思茅松林分直径大小多样性的影响。
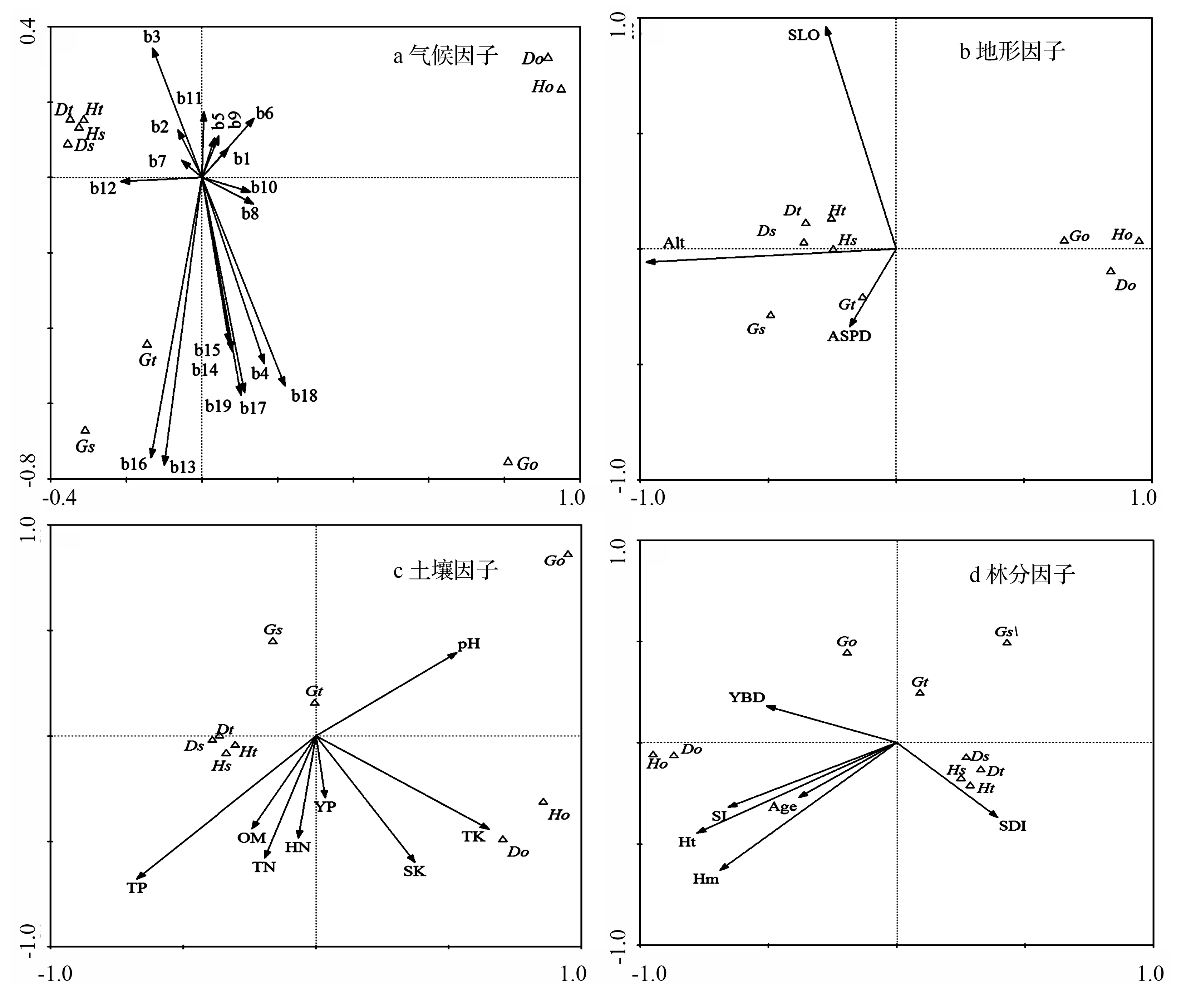
图3 环境因子CCA二维排序图
(1)采用3个与胸径有关的指数来描述思茅松林的林木直径大小多样性,3个指数可以较好地体现样地、思茅松和样地内其他树种的直径结构以及直径大小多样性。从Shannon指数上分析,说明样地内思茅松径阶级数比其他树种丰富,同时大部分样地内Hs>H,说明思茅松的径阶丰富程度基本上就是林分总的径阶丰富程度;从Simpson指数分析说明样地内其他树种处于比较集中的胸径范围内;从断面积Gini系数分析,G总体上比Gs和Go大,说明样地内思茅松和其他树种的直径分布比较均匀;而总体的直径分布悬殊比较大,样地内思茅松直径径阶比较丰富,且在一定程度上决定了样地整体的直径大小多样性。
(2)多样性指数CCA排序分析结果较好地反映了林分直径大小多样性随环境因子的变化规律,且4类因子中,地形因子最好地解释了林分直径结构变化,与其他因子的CCA排序结果比较,气候因子与第一轴的相关性并不突出,但也解释了58.6%的信息量,原因可能是3个研究点均处于一个地级市范围,气候表现一致,气温降水差异不大,反倒是地形因素的相关性较突出,表现出地形对林分直径大小多样性的影响,即在同一个气候区内,地形是影响植物群落重要的因子之一[21]。
(3)林分直径多样性与气候因子具有密切的关系。温度和降水对树木生长的影响在众多的气象因子中表现比较突出[22]。本研究中,描述林分直径多样性的指标与降水基本呈现比较显著的负相关关系,但是仅与几个温度因子呈现明显的相关性,说明CCA排序分析较好的拟合了林分直径多样性与降水因子之间的相关关系。断面积Gini系数与最大月和最湿季降水相关关系较大;Shannon和Simpson指数则与最热月均温和最冷月均温差、等温性和年降水等具有一定的相关性。
(4)地形是影响林木生长的重要因素,有研究表明,海拔比坡度和坡向对群落的结构特征影响大[23]。就本研究而言地形因素中海拔、坡度与思茅松天然林林分直径大小多样性存在显著的相关性,说明这两个因子是主要影响因素。
(5)土壤是森林植物生长发育的基础。从前述分析可以看出,对于思茅松天然林林分直径大小多样性影响较大的土壤因子主要有土壤pH值、土壤有机质以及磷和氮元素等。蒋云东等[24]的研究表明,土壤的化学性质对思茅松生长的影响较大的是速效P、pH值、有机质、全P、全N和全K等,本研究在土壤因子方面与其结论较一致。
(6)林分密度指数与Shannon和Simpson指数具有显著关系,主要是由于林分密度影响胸径的生长,随着林分密度的增加,林分平均胸径减小,直径分布的离散程度就越大[25];此外杨利华等[26]在研究林分密度对思茅松的生长量的影响时指出,思茅松不同密度对胸径具有显著影响,并且随林龄增大,密度效应更加显著;同时立地指数、郁闭度、林分优势高等,对思茅松林分直径大小多样性也有一定程度影响。
降水因素,海拔(Alt)、全磷(TP)、全钾(TK)和林分优势高(Ht)是气候、地形、土壤与林分因子中影响林分直径大小多样性的主要因素,Ht、Hs、Dt、Ds随环境因子的变化趋势一致,思茅松天然林具有最大的Ht、Hs、Dt、Ds值时,它们在相同的气候、地形、土壤和林分因子条件,而其他树种的林分直径多样性指数以及3个断面积Gini系数的规律性不强。
引入生态学中Shannon指数、Simpson指数和Gini系数作为评价思茅松天然林林分直径大小多样性的指标,并且结合生态学排序的方法研究了环境因子对思茅松天然林林分直径大小多样性的影响,这在思茅松的研究中较为少见。但本研究只考虑了与距离无关的多样性测度方法,没有考虑空间效应对直径大小多样性的影响,同时Shannon指数的分组缺乏统一的标准[5],本文以2 cm为径阶分组依据,并没有其它考虑,在分析林分直径大小多样性与不同环境因子的关系时,不同环境因子的相互作用没有考虑进来,以上几点有待进一步研究。
[1] 张连金,胡艳波,赵中华,等.北京九龙山侧柏人工林空间结构多样性[J].生态学杂志,2015,34(1):60-69.
[2] POMMERENING A. Approaches to quantifying forest structures[J]. Forestry,2002,75(3):305-324.
[3] 孟宪宇.测树学[M].3版.北京:中国林业出版社,1996:88-288.
[4] 李俊,佘济云,胡焕香,等.昌化江流域天然林直径结构研究[J].中南林业科技大学学报,2012,32(3):37-43.
[5] 雷相东,唐守正.林分结构多样性指标研究综述[J].林业科学,2002,38(3):140-146.
[6] LEXERØD N L, EID T. An evaluation of different diameter diversity indices based on criteria related to forest management planning[J]. Forest Ecology and Management,2006,222(1):17-28.
[7] 肖兴威.影响亚热带东部森林结构的因子分析[J].东北林业大学学报,2004,32(5):19-20.
[8] 陈育峰.气候-森林响应过程敏感性的初步研究:以四川西部紫果云杉群落为例[J].地理学报,1996(S1):58-65.
[9] 范叶青,周国模,施拥军,等.地形条件对毛竹林分结构和植被碳储量的影响[J].林业科学,2013,49(11):177-182.
[10] 舒树淼,赵洋毅,段旭,等.基于结构方程模型的云南松次生林林木多样性影响因子[J].东北林业大学学报,2015,43(10):63-67.
[11] 张金屯.数量生态学[M].北京:科学出版社,2004.
[12] 米湘成,张金屯,张峰,等.山西高原植被与气候的关系分析及植被数量区划的研究[J].植物生态学报,1996,20(6):549-560.
[13] 云南森林编写委员会.云南森林[M].昆明:云南科技出版社,1986.
[14] 温庆忠,赵元藩,陈晓鸣,等.中国思茅松林生态服务功能价值动态研究[J].林业科学研究,2010,23(5):671-677.
[15] 李江,孟梦,邱琼,等.思茅松中幼龄人工林生物量及生产力动态[J].东北林业大学学报,2010,38(8):36-38.
[16] 欧光龙,王俊峰,胥辉,等.思茅松天然林林分胸径与树高分布变化研究[J].广东农业科学,2013,40(21):54-57.
[17] 欧光龙,王俊峰,胥辉,等.思茅松天然林胸径与树高结构的变化[J].中南林业科技大学学报,2014,34(1):37-41.
[18] SHANNON C E. Communication theory of secrecy systems[J]. Bell System Technical Journal,1949,28(4):656-715.
[19] SIMPSON E H. Measurement of diversity[J]. Nature,1949,21:213-251.
[20] GINI C. Measurement of inequality of incomes[J]. The Economic Journal,1921,31:124-126.
[21] PALMER M W, DIXON P M. Small-scale environmental heterogeneity and the analysis of species distribution along gradients[J]. Journal of Vegetation Science,1990,1(1):57-65.
[22] 高洪娜,高瑞馨.气象因子对树木生长量影响研究综述[J].森林工程,2014,30(2):6-9.
[23] 朱彪,陈安平,刘增力,等.广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局[J].生物多样性,2004,12(1):44-52.
[24] 蒋云东,李思广,杨忠元,等.土壤化学性质对思茅松人工幼林生长的影响[J].东北林业大学学报,2006,34(1):25-27.
[25] 陈学群,朱配演,濑川幸三.不同密度30年生马尾松林分生产结构与现存量的研究[J].福建林业科技,1994(2):19-23.
[26] 杨利华,徐玉梅,杨德军,等.不同造林密度对思茅松中龄林生长量的影响[J].江苏林业科技,2013,40(6):43-46.
Stand Diameter Size Diversity and Their Environmental Explanation inPinuskesiyavar.langbianensisNatural Forests//
Li Chao, Lü Yanyu, Xu Hui, Xu Tingting, Zhang Bo, Wei Anchao, Sun Xuelian, Xiong Hexian, Shi Xiaolin, Ou Guanglong
(Key Laboratory of State Forestry Administration on Biodiversity Conservation in Southwest China, Southwest Forestry University, Kunming 650224, P. R. China)//Journal of Northeast Forestry University,2016,44(11):24-30.
Stand diameter diversity; Climate factors; Topographical factors; Soil factors; Stand factors; CCA ordination;Pinuskesiyavar.langbianensis
1)国家自然科学基金项目(31560209,31160157);西南林业大学博士科研启动基金项目(11146)。
李超,男,1991年2月生,西南地区生物多样性保育国家林业局重点实验室(西南林业大学),硕士研究生。E-mail:gislichao@163.com。
欧光龙,西南地区生物多样性保育国家林业局重点实验室(西南林业大学),副教授。E-mail:olg2007621@126.com。
2016年5月3日。
S715.3
责任编辑:王广建。
猜你喜欢
东北林业大学学报(2021年1期)2021-12-03 19:17:48
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
东北林业大学学报(2021年1期)2021-01-14 04:26:10
新农业(2020年18期)2021-01-07 02:17:08
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
山东林业科技(2017年1期)2017-06-29 07:53:58
上海农业学报(2016年5期)2016-02-10 06:53:24
