
环状RNA的生物学功能及其在肿瘤发生中的作用
2016-12-12 05:13谢依李田文莫小燕郭俊明
生物工程学报 2016年11期
谢依,李田文,莫小燕,郭俊明
宁波大学医学院生物化学与分子生物学研究所 浙江省病理生理学技术研究重点实验室,浙江 宁波 315211
环状RNA的生物学功能及其在肿瘤发生中的作用
谢依,李田文,莫小燕,郭俊明
宁波大学医学院生物化学与分子生物学研究所 浙江省病理生理学技术研究重点实验室,浙江 宁波 315211
谢依, 李田文, 莫小燕, 等. 环状RNA的生物学功能及其在肿瘤发生中的作用. 生物工程学报, 2016, 32(11): 1507-1518.
Xie Y, Li TW, Mo XY, et al. Circular RNAs and their roles in tumorigenesis. Chin J Biotech, 2016, 32(11): 1507-1518.
近年来,随着RNA研究技术的进步,研究者们在多种生物中发现了数量众多的环状RNA,且发现它们具有重要的生物学功能。环状RNA来源于内含子或外显子,可以充当微小RNA海绵,还能与蛋白质相结合,从而参与基因表达调控并影响蛋白质的功能,此外,个别环状RNA甚至能编码蛋白质。更重要的是,环状RNA在肿瘤 (如:胃癌、肝癌、结直肠癌、乳腺癌、宫颈癌和卵巢癌等) 的发生和发展过程中起着重要的调控作用。因此,环状RNA有希望成为肿瘤诊断的标志物和治疗的新靶点。
环状RNA,微小RNA海绵,肿瘤,发生
环状RNA (Circular RNA, circRNA) 是广泛存在于转录组中的一类闭合环状的RNA分子。参与生物体多种生命活动的调控。circRNA不易被核酸酶降解,比线性RNA更稳定,可能是一种较为理想的疾病诊断标志物。
随着转录组研究的深入和RNA测序技术的发展,科研工作者在生物体内检测到大量的circRNA。它们与蛋白质相互作用可以影响细胞周期进程、细胞衰老和凋亡等基本生命活动。另外,circRNA还与胃癌、结直肠癌、乳腺癌、胆管癌、胰腺癌和卵巢癌等肿瘤的发生和发展有着直接或间接的联系。本文针对国内外circRNA的最新研究进展,对其种类、生物合成、特征、功能及其与肿瘤的关系作一综述。
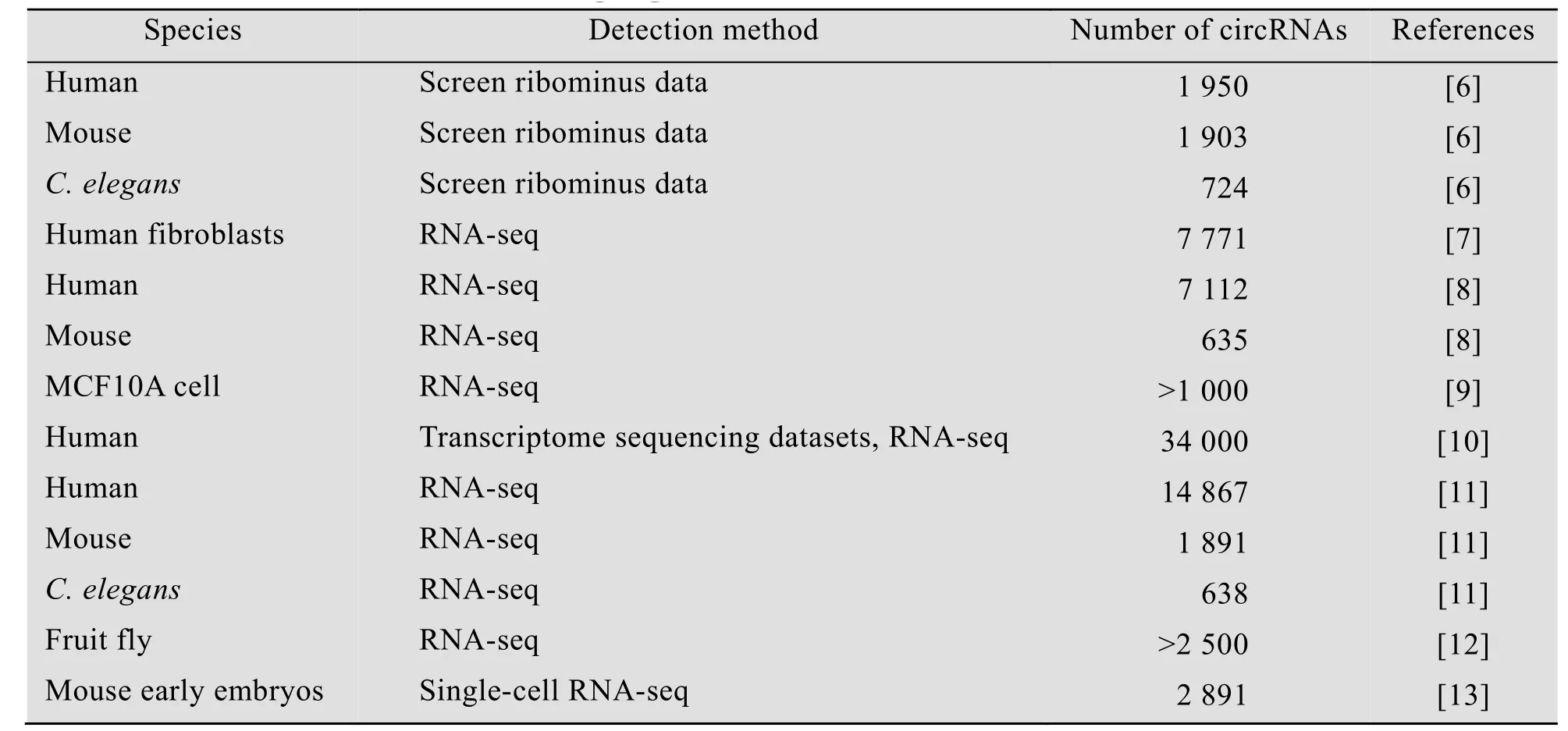
表1 环状RNA在生物体的分布情况Table 1 Distribution of circRNAs among organisms
1 环状RNA的分类
circRNA是一类长期被忽视的、却具有重要生物学功能的RNA分子。早在1976年,第一个circRNA就在病毒中被发现[1]。随后,研究者们在不同种类的生物体中发现了越来越多的circRNA,包括古生菌circRNA[2-3]、小鼠circRNA[4]以及人类circRNA[5]等。
通过RNA测序、单细胞RNA测序等技术,科研人员已经在多种生物体中发现了数目可观的circRNA[6-13](表1)。
目前发现的circRNA根据其来源主要分为3类:内含子序列形成的circRNA (Circular intronic RNA, ciRNA)[14]、外显子序列形成的circRNA(Exonic circular RNA, ecircRNA)[6-7,15-16]以及内含子序列和外显子序列共同形成的circRNA (Exon-intron circular RNA, EIciRNA)[17]。人类和小鼠的绝大部分circRNA来自编码基因[7,18]。

图1 环状RNA的生成形式Fig. 1 Models of circRNA formation.
2 环状RNA的生物合成
circRNA在单细胞生物中主要来自前核糖体RNA基因转录本的自我剪接[19],但是在古生菌中则由蛋白质编码基因转录生成[3]。动物中circRNA主要来源自剪接体中同一外显子5'端和下游3'序列环化而成[18]。3种circRNA的生成方式各不相同 (图1)。
在ecircRNA的形成过程中,最常见的方式是索尾剪接。大致分为两步:首先,待环化外显子分叉点上游的2'-羟基攻击其下游5'剪接位点,形成一个Y型结构的中间产物;其次,待环化外显子第一步新形成的3'-羟基末端攻击上游3'剪接位点,最后释放一个环状RNA[20]。而在形成ecircRNA时,保留了内含子序列则会产生EIciRNA。
索尾剪接时,circRNA的形成依赖侧翼的互补序列,包括重复的或者非重复元件,序列的完美配对有利于circRNA的高效形成[15]。另外,剪切位点之间应该彼此靠近才能促进索尾剪接。这一步尽管机理简单,却具有高度选择性,这些重复序列能够彻底改变circRNA的产生效率。研究发现,越长的外显子似乎环化效率越高,此外,3'端的加工是环化所必需的,这表明circRNA的生成可能在转录后阶段[21]。
但在ciRNA的形成过程中,关键序列(比如某些内含子序列)是环化正常进行所必需的[14]。有研究者发现,ciRNA的形成过程中存在一些共有的保守序列,如:5'剪切位点附近存在一段7 nt左右富含GU碱基的结构,分支点附近存在一段11 nt左右富含C碱基的结构[14]。实际上,这些长度确定的关键RNA元件增加了ciRNA的形成效率,而其他顺式作用元件可能进一步促进了该circRNA的生成[14]。
有趣的是,Zhang等[15]研究发现,同一基因位点可以生成多种circRNA,比如,人类CAMSAP1基因至少可以产生7种不同的ecircRNA,他们把这种现象叫作可变环化 (Alternative circularization, AC)。理论上,这一特点将使circRNA在调控基因表达方面更加灵活和方便。
3 环状RNA的生物学特征
多数circRNA来自外显子的索尾剪接,并且在细胞质中稳定存在[6-7]。细胞质中的circRNA大多具有48 h以上的半衰期[7],而mRNA的半衰期平均只有10 h[22]。circRNA的稳定性可能是由于它们缺乏类似mRNA 的5'帽子和3'尾结构,所以可以避免脱腺苷化、脱帽反应以及与mRNA相关的正常降解反应[23]。一些circRNA在不同组织和不同发育阶段特异性表达[6]。circRNA的保守核苷酸序列丰度明显增加,表明circRNA可与其他RNA或者微小RNA (MicroRNA, miRNA) 竞争性结合RNA结合蛋白 (RNA binding protein, RBP)[6]。因为竞争性内源RNA (Competing endogenous RNA, ceRNA)通过自身的miRNA应答元件 (MicroRNA response element, MRE) 竞争性结合同一miRNA[24],与其他具有MRE的ceRNA (假基因转录本、lncRNA和mRNA等) 相比,circRNA因其特殊结构而具有的稳定性使其成为一种优势ceRNA,对ceRNA网络的平衡起着重要作用。
大部分人类circRNA仅在少数几种细胞类型中表达且表达水平很低,它们的细胞特异性没有相应的mRNA强。尽管多数circRNA含有蛋白质编码序列,但是核糖体分析却无法证明它们能指导蛋白质的合成。有人发现635种小鼠circRNA中有20%与人类circRNA是直系同源,但这些同源circRNA的序列保守程度没有其邻近的同源线性外显子高[8]。
细胞质中来自索尾剪接的circRNA的主要作用之一是作为miRNA海绵[25],而在细胞核中含量丰富的ciRNA的功能不同于细胞质circRNA[14]。一系列实验表明,ciRNA与某些基因的调控相关;在细胞核中含量丰富的ciRNA可能参与基因表达的顺式调控[14]。ciRNA在人类细胞中广泛表达,但进化上的保守性较低[14]。另外,研究者还发现一些ciRNA具有物种特异性[14]。因而可利用不同组织和物种特异性的相同或相似性去鉴别其他的ciRNA。
4 环状RNA的生物学功能
生物大分子的生物学功能与其结构和特性有着密切联系。目前circRNA功能的相关研究表明,它们主要在miRNA海绵、转录调控、结合蛋白质和翻译等4个方面发挥作用 (图2)。
4.1 miRNA海绵
充当miRNA海绵是目前circRNA研究中最热点的方向之一,同时也可能是其较为普遍的作用。
在可充当miRNA海绵的circRNA中,最典型是ciRS-7 (Circular RNA sponge for miR-7)。它含有70多个保守的miRNA结合位点,并且很可能与依赖miR-7的阿格蛋白 (Argonaute, AGO) 有关系。该circRNA强烈抑制miR-7的活性,从而增加miR-7靶标的水平[25]。
另一个作为miRNA海绵的circRNA是雄性性别决定基因 (Sex-determining region Y, Sry)的睾丸特异性转录本,它含有miR-138的16个保守结合位点[25]。
新发现的心脏相关环状RNA (Heart-related circRNA, HRCR)作为miR-223的海绵,可以抑制miR-223的活性,导致miR-223的下游基因表达量增加,最终减缓心肌肥厚和心力衰竭的进程[26]。
miRNA海绵是circRNA作为ceRNA的主要作用。在生物体内,ceRNA网络复杂而精细,任何微小的紊乱都会改变基因的表达,导致肿瘤的发生,而circRNA作为其中的重要成员,对维护ceRNA网络的平衡必不可少;另外,研究circRNA的miRNA海绵作用或许可以解释某些肿瘤发生的分子机理。
4.2 调控转录
小鼠形成素 (Formin) 对肢体发育至关重要,其基因fmn可转录形成一个circRNA。在索尾剪接过程中涉及到fmn基因编码序列上游的一个剪接受体,若敲除小鼠fmn基因中的这个剪接受体,将检测不到该circRNA,小鼠肢体正常发育,但有不完全渗透肾发育不全的表型[27]。这说明该circRNA的剪接受体参与了小鼠基因的转录与翻译,若缺失它,则会引起某些疾病。
结合生物化学、细胞功能检测和生物信息学分析,研究者们发现小脑变性相关蛋白1反义转录物 (Antisense to the cerebellar degenerationrelated protein 1 transcript, CDR1as) 这一circRNA具有下调miR-7的功能,而miR-7在线虫与人类中均具有完整的序列保守性[28]。这也表明circRNA是一个重要的转录后调控者[6]。
另外,Li等[17]研究发现,一些EIciRNA通过与U1核内小分子RNA (Small nuclear RNA, snRNA) 的相互作用调控亲本基因的表达,EIciRNA先与U1小分子核糖核蛋白体 (Small nuclear ribonucleoprotein, snRNP) 形成EIciRNAU1 snRNP复合物,该复合物再与RNA聚合酶Ⅱ转录复合物相互作用促进基因的表达。这种RNA-RNA相互作用调控转录的模式既丰富了circRNA的功能,也为研究circRNA提供了一个新方向。

图2 环状RNA的主要功能Fig. 2 Main functions of circRNA.
4.3 与RNA结合蛋白相互作用
circRNA能稳定地与AGO蛋白和RNA聚合酶Ⅱ结合。另外,它们还可以与多种RBP结合充当RBP的脚手架[29]。circRNA也可能作为目标序列元件,利用其互补序列同时结合RBP、RNA或者DNA[6]。
Ashwal-Fluss等[30]发现,剪接因子Muscleblind (MBL/MBNL1) 第2个外显子形成的circRNA (circMBL) 能结合RBP。当细胞内含有过量的MBL蛋白时,会促进circMBL的生成从而减少该蛋白的mRNA产量,而circMBL则会与过量的MBL蛋白相结合,使MBL蛋白的含量趋于正常。
最近,研究者发现叉形头盒O3环状RNA (CircRNA-forkhead box O3, circ-Foxo3) 在成年小鼠和年长的心脏病患者的心脏组织中过量表达;进一步研究表明,circ-Foxo3能与缺氧诱导因子 (Hypoxia inducible factor 1α, HIF1α) 和局部粘着斑激酶 (Focal adhesion kinase, FAK) 相结合[31]。因为HIF1α和FAK具有抑制细胞衰老的作用,当细胞质中circ-Foxo3的表达量增加时,这些蛋白质被捕获,它们的功能无法发挥,会导致细胞衰老的进程加快。由此可见,circ-Foxo3的异常高表达可以促进细胞衰老,而沉默该circRNA可以减缓细胞衰老。另外,circ-Foxo3还能与细胞周期蛋白依赖性激酶2 (Cyclindependent kinase 2, CDK2) 和细胞周期蛋白依赖性激酶抑制剂P21形成三元复合物抑制CDK2的活性,从而将细胞阻滞在G1/S期,影响细胞周期的进展,抑制细胞增殖[32]。
上述circ-Foxo3的研究中,该circRNA均通过与功能蛋白质相结合使后者失活,间接发挥调控作用。这说明,单个circRNA的功能并不单一,可能会涉及到多种蛋白质。
4.4 环状RNA翻译成多肽
在哺乳动物细胞内,已检测到可编码蛋白质的外源性circRNA,如:来源于丁型肝炎病毒(Hepatitis D virus, HDV)的RNA[33]。HDV是乙型肝炎病毒 (Hepatitis B virus, HBV) 的卫星病毒。HDV的circRNA可表达产生致病性病毒蛋白[29]。
如果circRNA含有内部核糖体进入位点,则有可能指导蛋白质合成,真核生物核糖体40S小亚基能够进入这些circRNA的内部核糖体进入位点,从而指导蛋白质合成[34]。实验证明,如果circRNA含有的碱基是3的倍数,则该编码蛋白质的circRNA会沿着阅读框循环阅读,从而翻译出一段重复的多肽序列[34]。但是到目前为止,尚无较多证据证明真核生物内源circRNA可以指导蛋白质合成,也没有发现由circRNA翻译生成的多肽。
但是,许多circRNA含有起始密码子,有些甚至含有线性转录本中典型的AUG启动子,所以探索内源性circRNA作为转录本依然是一个活跃的研究方向[7]。
以上研究表明,HDV是目前仅有的能编码蛋白质的circRNA。尽管多数circRNA来自编码区,甚至含有起始密码子,但它们仍属于非编码RNA。
5 环状RNA与肿瘤
近年来,越来越多的研究表明,circRNA通过直接或者间接作用,在肿瘤发生和发展过程中扮演着重要角色。比如,有研究发现,结直肠癌组织细胞中总circRNA丰度与癌症转移程度存在联系[35];本课题组首次发现,Has_circ_002059与胃癌患者的多种临床病理因素密切相关,有希望成为胃癌诊断的新型标志物[36]。为此,深入了解circRNA在肿瘤发生和发展中的作用,将有利于揭示肿瘤的发生机制,寻找肿瘤诊断和治疗的新方法。
5.1 环状RNA通过调控miR-7影响肿瘤的发生和发展
miRNA是生物体内普遍存在的一种调控性非编码RNA。它们通常在转录后水平通过降解或者抑制目标mRNA来调控基因表达[37]。miRNA可起致癌或抑癌作用。一些circRNA含有miRNA的结合位点,可以调控miRNA的生物学功能,从而间接参与癌症的发生和发展过程。在非编码RNA的调控网络中,circRNA和miRNA的结合与竞争关系为我们理解癌症发生和发展过程提供了新思路。
如上所述,ciRS-7能结合miR-7,从而抑制miR-7发挥其生物学功能[25]。例如:miR-7可以调控人类肿瘤细胞中表皮生长因子受体(Epidermal growth factor receptor, EGFR) 的表达,它通过结合EGFR mRNA 3'非翻译区(Untranslated region, UTR) 降低EGFR的mRNA和蛋白质的水平[38]。miR-7抑制EGFR信号通路还通过钝化蛋白激酶B (PKB/Akt) 和细胞外信号调节激酶1/2 (Erk1/2)[39-41]。因此,circRNA通过调控miR-7表达来影响肿瘤细胞生长——作为miRNA海绵,过表达ciRS-7可抑制miR-7的表达从而增加EGFR的表达量。EGFR过表达与多种肿瘤(包括肿瘤干细胞、胃癌、肝细胞癌和结直肠癌等) 发展、对化疗和放疗的抵抗以及不良预后有关[41-42]。
在胃癌细胞中,miR-7可抑制胰岛素生长因子受体 (Insulin-like growth factor 1 receptor, IGF1R) 的表达。IGF1R表达的减少增加了E钙黏蛋白的表达量,部分逆转了上皮-间质转化,结果使癌细胞转移和增殖受阻,延缓了癌细胞的生长与侵袭的过程[43]。因此,ciRS-7使miR-7下调的结果是IGF1R原癌基因抑制解除,诱导癌细胞迁移和增殖。
在肝癌细胞中,若上调miR-7的表达,可能会使细胞周期停滞在G1/S期,从而抑制癌细胞增殖[44]。因为miR-7能结合到细胞周期素E1 (Cyclin E1, CCNE1) mRNA的3'UTR上,直接沉默其表达;还作为CDK2的一个调控亚基,在G1期到S期过渡时起作用[44]。肝癌细胞中miR-7的其他靶标——磷脂酰肌醇-3激酶催化亚基δ (Phosphoinositide 3-kinase catalytic subunit delta, PIK3CD)、哺乳动物雷帕霉素靶蛋白 (Mammalian target of rapamycin, mTOR) 和P70s6k,均被miR-7的过表达所抑制[41]。这表明,miR-7通过阻止细胞周期中的G0/G1阶段以及干扰PI3K/Akt/mTOR信号通路来影响肝癌细胞的细胞周期。
结直肠癌中,miR-7具有肿瘤抑制的作用。它可以抑制癌基因yin yang1的表达[45]。该致癌基因通过抑制P53,调控下游的P15效应器、Caspase级联反应和原癌基因c-jun来起作用;并且通过活化β-链蛋白、生存素和纤维母细胞生长因子-4来激活Wnt信号通路[45-46]。因此,miR-7能够抑制结直肠癌细胞增殖,诱导细胞凋亡。
除了消化道肿瘤外,miR-7与其他癌症也有关系。miR-7通过抑制多种肿瘤相关信号通路抑制Her-2调控的乳腺癌,并且部分逆转曲妥珠单抗耐性[47-49]。此外,miR-7可增加非小细胞肺癌细胞对紫杉醇敏感性[50]。一项宫颈癌的研究发现,miR-7在转录和翻译水平通过以黏着斑激酶为靶标抑制宫颈癌的转移和侵袭[51]。
由上可知,miR-7调控多种与肿瘤密切相关的信号通路,而ciRS-7可以通过结合miR-7而间接影响肿瘤的发生和发展过程。
5.2 环状RNA通过调控miR-138影响肿瘤的发生和发展
除了ciRS-7外,还有其他一些circRNA对肿瘤的发生和发展有着重要影响。
sry是雄性性别决定基因所表达的睾丸特异性环状转录产物[8,52]。作为一种miRNA海绵,sry通过结合到miR-138上的16个保守位点来调控其生物学功能。
在胆管癌细胞中,sry抑制miR-138的表达,导致Ras同源基因家族的成员C (Ras homolog gene family member C, RhoC) 的mRNA及其编码的蛋白质表达量增加,结果使RhoC/p-ERK/ MMP-2/MMP-9通路上调,从而促进胆管癌细胞的增殖、迁移以及侵袭[53]。在结直肠癌中,miR-138直接以碱性螺旋-环-螺旋家族成员Twist2作为靶标,阻碍其功能,最终抑制了结直肠癌细胞的迁移和侵袭[54]。在卵巢癌中,miR-138通过阻止增殖细胞核抗原 (Proliferating cell nuclear antigen, PCNA) 和B细胞淋巴瘤-2通路来抑制癌细胞的生长;而抑制miR-138时,LIM激酶1 (Limk1)/cofilin信号通路被激活,将导致卵巢癌细胞的转移[55-57]。小细胞肺癌细胞中,miR-138以H2A组蛋白家族成员X (H2A histone family member X, H2AX) 作为靶标,参与调控DNA损伤应答;它的上调增加了肺癌细胞的辐射敏感度[58-59]。
miR-138不仅在实体瘤中,也在慢性淋巴白血病 (Chronic lymphocytic leukemia, CLL) 中起重要作用。Berg等[60]研究表明,在CLL中,miR-138的显著下调与抑制CD95调控的细胞凋亡有关。
circRNA作为一种重新引起研究者们重视的调控分子,目前关于其功能的研究多数都致力于发现其与下游靶标的相互作用,比如ciRS-7和 sry分别与它们下游靶标miR-7和miR-138的相互作用。对其他circRNA作为miRNA海绵的鉴定也一直在进行中,比如最近发现的环状RNAHRCR可作为miR-223的海绵[26]。由于成千上万的功能未知的circRNA陆续在不同物种中被鉴定出来,所以我们有理由相信将会发现越来越多的circRNA与相应miRNA之间的相互作用,以及它们参与更多种类癌症的调控过程。
5.3 环状RNA在肿瘤组织中的异常表达
从circRNA被发现至今,其与肿瘤的关系一直是研究的热点。研究者们希望从circRNA中寻找肿瘤的发病机理,以及诊断和治疗肿瘤的新方法。
研究发现,与癌旁组织相比,cir-ITCH的表达水平在食管鳞癌组织中明显下调[35],并且该circRNA也在结直肠癌组织中明显下调[61],Has_circ_002059、Has_circ_0001649和Has_circ_ 104912的表达水平分别在胃癌组织[36]、肝癌组织[62]和喉鳞癌组织中[63]下调;而Has_circ_ 100855的表达水平在喉鳞癌组织中明显上调[63]。
这些circRNA在癌组织中异常表达的现象,说明了circRNA与肿瘤的发生和发展相关。
6 展望
随着对circRNA研究的逐渐深入,我们对其了解也更加广泛。虽然circRNA在几十年前就被发现了,但最近几年才受到重视,关于它的重大发现也越来越多。circRNA具有结构特殊性、生物学性质稳定性等特点,这为其成为新型肿瘤诊断生物标志物奠定了基础,并已有研究结果支持这方面的应用,如:本课题组发现Has_circ_002059具有作为胃癌新型标志物的可能性[36];Lukiw等[64]研究表明,患有阿兹海默症的患者体内的CDR1as表达水平减少。另一方面,在寻找肿瘤诊断标志物方面,因采集体液 (唾液、血液等) 创伤小、易于获得等特点,一直是研究者们最理想的选择。Li等[65]发现,外泌体中的circRNA (Exosome circRNA, exo-circRNA)含量丰富且性质稳定,而血清中多种exo-circRNA的表达量在癌症患者和健康人中具有明显差异,可据此将患者和正常人区分开。
circRNA虽然参与基因的表达调控、肿瘤的发生和发展,但目前发现大部分circRNA是通过调控miRNA间接实现其功能。circRNA与miRNA之间既存在竞争关系又存在协同关系,它们之间的平衡对生物体至关重要。我们可以以circRNA为靶标,来调控miRNA,进而调控一系列生命活动。所以,在临床上,有可能针对circRNA用于某些肿瘤的靶向治疗。这些都使circRNA具有很大的研究价值和应用前景。
REFERENCES
[1] Sanger HL, Klotz G, Riesner D, et al. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc Natl Acad Sci USA, 1976, 73(11): 3852-3856.
[2] Kjems J, Garrett RA. Novel splicing mechanism for the ribosomal RNA intron in the archaebacterium desulfurococcus mobilis. Cell, 1988, 54(5): 693-703.
[3] Danan M, Schwartz S, Edelheit S, et al. Transcriptome-wide discovery of circular RNAs in Archaea. Nucleic Acids Res, 2012, 40(7): 3131-3142.
[4] Capel B, Swain A, Nicolis S, et al. Circular transcripts of the testis-determining gene Sry in adult mouse testis. Cell, 1993, 73(5): 1019-1030.
[5] Burd CE, Jeck WR, Liu Y, et al. Expression of linear and novel circular forms of an INK4/ARF-associated non-coding RNA correlates with atherosclerosis risk. PLoS Genet, 2010, 6(12): e1001233.
[6] Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature, 2013, 495(7441): 333-338.
[7] Jeck WR, Sorrentino JA, Wang K, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA, 2013, 19(2): 141-157.
[8] Guo JU, Agarwal V, Guo HL, et al. Expanded identification and characterization of mammalian circular RNAs. Genome Biol, 2014, 15(7): 409.
[9] Enuka Y, Lauriola M, Feldman ME, et al. Circular RNAs are long-lived and display only minimal early alterations in response to a growth factor. Nucleic Acids Res, 2016, 44(3): 1370-1383.
[10] Liu YC, Li JR, Sun CH, et al. CircNet: a database of circular RNAs derived from transcriptome sequencing data. Nucleic Acids Res, 2016, 44(D1): D209-D215.
[11] Zheng LL, Li JH, Wu J, et al. DeepBase v2.0: identification, expression, evolution and function of small RNAs, LncRNAs and circular RNAs from deep-sequencing data. Nucleic Acids Res, 2016, 44(D1): D196-D202.
[12] Westholm JO, Miura P, Olson S, et al. Genome-wide analysis of Drosophila circular RNAs reveals their structural and sequence properties and age-dependent neural accumulation. Cell Rep, 2014, 9(5): 1966-1980.
[13] Fan XY, Zhang XN, Wu XL, et al. Single-cell RNA-seq transcriptome analysis of linear and circular RNAs in mouse preimplantation embryos. Genome Biol, 2015, 16(1): 148.
[14] Zhang Y, Zhang XO, Chen T, et al. Circular intronic long noncoding RNAs. Mol Cell, 2013, 51(6): 792-806.
[15] Zhang XO, Wang HB, Zhang Y, et al. Complementary sequence-mediated exon circularization. Cell, 2014, 159(1): 134-147.
[16] Salzman J, Chen RE, Olsen MN, et al. Cell-type specific features of circular RNA expression. PLoS Genet, 2013, 9(9): e1003777.
[17] Li ZY, Huang C, Bao C, et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol, 2015, 22(3): 256-264.
[18] Salzman J, Gawad C, Wang PL, et al. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE, 2012, 7(2): e30733.
[19] Grabowski PJ, Zaug AJ, Cech TR. The intervening sequence of the ribosomal RNA precursor is converted to a circular RNA in isolated nuclei of Tetrahymena. Cell, 1981, 23(2): 467-476.
[20] Starke S, Jost I, Rossbach O, et al. Exon circularization requires canonical splice signals. Cell Rep, 2015, 10(1): 103-111.
[21] Liang DM, Wilusz JE. Short intronic repeat sequences facilitate circular RNA production. Genes Dev, 2014, 28(20): 2233-2247.
[22] Schwanhäusser B, Busse D, Li N, et al. Global quantification of mammalian gene expression control. Nature, 2011, 473(7347): 337-342.
[23] Huntzinger E, Izaurralde E. Gene silencing by microRNAs: contributions of translational repression and mRNA decay. Nat Rev Genet, 2011, 12(2): 99-110.
[24] Salmena L, Poliseno L, Tay Y, et al. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell, 2011, 146(3): 353-358.
[25] Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges. Nature, 2013, 495(7441): 384-388.
[26] Wang K, Long B, Liu F, et al. A circular RNA protects the heart from pathological hypertrophy and heart failure by targeting miR-223. Eur Heart J, 2016, 37(33): 2602-2611.
[27] Chao CW, Chan DC, Kuo A, et al. The mouse formin (fmn) gene: abundant circular RNA transcripts and gene-targeted deletion analysis. Mol Med, 1998, 4(9): 614-628.
[28] Hansen TB, Wiklund ED, Bramsen JB, et al. MiRNA-dependent gene silencing involving Ago2-mediated cleavage of a circular antisense RNA. EMBO J, 2011, 30(21): 4414-4422.
[29] Jeck WR, Sharpless NE. Detecting and characterizing circular RNAs. Nat Biotechnol, 2014, 32(5): 453-461.
[30] Ashwal-Fluss R, Meyer M, Pamudurti NR, et al. CircRNA biogenesis competes with pre-mRNA splicing. Mol Cell, 2014, 56(1): 55-66.
[31] Du WW, Yang WN, Chen Y, et al. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur Heart J, 2016, doi: 10.1093/eurheartj/ehw001.
[32] Du WW, Yang WN, Liu E, et al. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res, 2016, 44(6):2846-2858.
[33] Kos A, Dijkema R, Arnberg AC, et al. The hepatitis delta (δ) virus possesses a circular RNA. Nature, 1986, 323(6088): 558-560.
[34] Chen CY, Sarnow P. Initiation of protein synthesis by the eukaryotic translational apparatus on circular RNAs. Science, 1995, 268(5209): 415-417.
[35] Bachmayr-Heyda A, Reiner AT, Auer K, et al. Correlation of circular RNA abundance with proliferation-exemplified with colorectal and ovarian cancer, idiopathic lung fibrosis, and normal human tissues. Sci Rep, 2015, 5: 8057.
[36] Li PF, Chen SC, Chen HL, et al. Using circular RNA as a novel type of biomarker in the screening of gastric cancer. Clin Chim Acta, 2015, 444: 132-136.
[37] Chen CZ. MicroRNAs as oncogenes and tumor suppressors. N Engl J Med, 2005, 353(17): 1768-1771.
[38] Chen YJ, Chien PH, Chen WS, et al. Hepatitis B virus-encoded X protein downregulates EGFR expression via inducing microRNA-7 in hepatocellularcarcinoma cells. Evid Based Complement Alternat Med, 2013, 2013: 682380.
[39] Zhou XC, Hu Y, Dai L, et al. MicroRNA-7 inhibits tumor metastasis and reverses epithelial-mesenchymal transition through AKT/ERK1/2 inactivation by targeting EGFR in epithelial ovarian cancer. PLoS ONE, 2014, 9(5): e96718.
[40] Webster RJ, Giles KM, Price KJ, et al. Regulation of epidermal growth factor receptor signaling in human cancer cells by microRNA-7. J Biol Chem, 2009, 284(9): 5731-5741.
[41] Fang YX, Xue JL, Shen Q, et al. MicroRNA-7 inhibits tumor growth and metastasis by targeting the phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma. Hepatology, 2012, 55(6): 1852-1862.
[42] Janbabai G, Oladi Z, Farazmandfar T, et al. The prognostic impact of EGFR, ErbB2 and MET gene amplification in human gastric carcinomas as measured by quantitative real-time PCR. J Cancer Res Clin Oncol, 2015, 141(11): 1945-1952.
[43] Zhao X, Dou W, He L, et al. MicroRNA-7 functions as an anti-metastatic microRNA in gastric cancer by targeting insulin-like growth factor-1 receptor. Oncogene, 2013, 32(11): 1363-1372.
[44] Zhang X, Hu SJ, Zhang X, et al. MicroRNA-7 arrests cell cycle in G1 phase by directly targeting CCNE1 in human hepatocellular carcinoma cells. Biochem Biophys Res Commun, 2014, 443(3): 1078-1084.
[45] Chinnappan D, Xiao DM, Ratnasari A, et al. Transcription factor YY1 expression in human gastrointestinal cancer cells. Int J Oncol, 2009, 34(5): 1417-1423.
[46] Zhang N, Li X, Wu CW, et al. MicroRNA-7 is a novel inhibitor of YY1 contributing to colorectal tumorigenesis. Oncogene, 2013, 32(42): 5078-5088.
[47] Huynh FC, Jones FE. MicroRNA-7 inhibits multiple oncogenic pathways to suppress HER2Δ16 mediated breast tumorigenesis and reverse trastuzumab resistance. PLoS ONE, 2014, 9(12): e114419.
[48] Babae N, Bourajjaj M, Liu YJ, et al. Systemic miRNA-7 delivery inhibits tumor angiogenesis and growth in murine xenograft glioblastoma. Oncotarget, 2014, 5(16): 6687-6700.
[49] Liu ZL, Jiang ZM, Huang JY, et al. MiR-7 inhibits glioblastoma growth by simultaneously interfering with the PI3K/ATK and Raf/MEK/ERK pathways. Int J Oncol, 2014, 44(5): 1571-1580.
[50] Liu RH, Liu XM, Zheng YJ, et al. MicroRNA-7 sensitizes non-small cell lung cancer cells to paclitaxel. Oncol Lett, 2014, 8(5): 2193-2200.
[51] Hao ZF, Yang JS, Wang CH, et al. MicroRNA-7 inhibits metastasis and invasion through targeting focal adhesion kinase in cervical cancer. Int J Clin Exp Med, 2015, 8(1): 480-487.
[52] Granados-Riveron JT, Aquino-Jarquin G. Does the linear Sry transcript function as a ceRNA for miR-138? The sense of antisense. F1000Res, 2014, 3: 90.
[53] Wang Q, Tang HH, Yin SS, et al. Downregulation of microRNA-138 enhances the proliferation, migration and invasion of cholangiocarcinoma cells through the upregulation of RhoC/p-ERK/MMP-2/MMP-9. Oncol Rep, 2013, 29(5): 2046-2052.
[54] Long LM, Huang GQ, Zhu HY, et al. Down-regulation of miR-138 promotes colorectal cancer metastasis via directly targeting TWIST2. J Transl Med, 2013, 11(1): 275.
[55] Ayaz L, Çayan F, Balci Ş, et al. Circulating microRNA expression profiles in ovarian cancer. J Obstet Gynaecol, 2014, 34(7): 620-624.
[56] Chen PX, Zeng MJ, Zhao Y, et al. Upregulation of Limk1 caused by microRNA-138 loss aggravates the metastasis of ovarian cancer by activation of Limk1/cofilin signaling. Oncol Rep, 2014, 32(5): 2070-2076.
[57] Yeh YM, Chuang CM, Chao KC, et al. MicroRNA-138 suppresses ovarian cancer cell invasion and metastasis by targeting SOX4 andHIF-1α. Int J Cancer, 2013, 133(4): 867-878.
[58] Yang H, Tang Y, Guo W, et al. Up-regulation of microRNA-138 induce radiosensitization in lung cancer cells. Tumor Biol, 2014, 35(7): 6557-6565.
[59] Yang H, Luo JW, Liu ZG, et al. MicroRNA-138 regulates DNA damage response in small cell lung cancer cells by directly targeting H2AX. Cancer Invest, 2015, 33(4): 126-136.
[60] Berg V, Rusch M, Vartak N, et al. MiRs-138 and -424 control palmitoylation-dependent CD95-mediated cell death by targeting acyl protein thioesterases 1 and 2 in CLL. Blood, 2015, 125(19): 2948-2957.
[61] Huang GL, Zhu H, Shi YX, et al. Cir-ITCH plays an inhibitory role in colorectal cancer by regulating the Wnt/β-catenin pathway. PLoS ONE, 2015, 10(6): e0131225.
[62] Qin ML, Liu G, Huo XS, et al. Hsa_circ_0001649: a circular RNA and potential novel biomarker for hepatocellular carcinoma. Cancer Biomark, 2016, 16(1): 161-169.
[63] Xuan LJ, Qu LM, Zhou H, et al. Circular RNA: a novel biomarker for progressive laryngeal cancer. Am J Transl Res, 2016, 8(2): 932-939.
[64] Lukiw WJ. Circular RNA (circRNA) in Alzheimer's disease (AD). Front Genet, 2013, 4: 307.
[65] Li Y, Zheng QP, Bao CY, et al. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res, 2015, 25(8): 981-984.
(本文责编 郝丽芳)
Circular RNAs and their roles in tumorigenesis
Yi Xie, Tianwen Li, Xiaoyan Mo, and Junming Guo
Zhejiang Key Laboratory of Pathophysiology, Department of Biochemistry and Molecular Biology, School of Medicine, Ningbo University, Ningbo 315211, Zhejiang, China
Recently, with the development of RNA research techniques, a wide variety of circular RNAs (circRNAs) have been discovered and some of them are confirmed to have crucial biological functions. CircRNAs arise from exons (i.e. exonic circRNAs) or introns (i.e. intronic circRNAs). Acting as microRNA sponges or combining with proteins, circRNAs participate in the regulation of gene expression and influence the activity of some proteins. In addition, some circRNAs even encode proteins. More importantly, several circRNAs play a key role in the occurrence and progression of some tumors, including stomach, liver, colon, breast, cervical, and ovarian cancers. Therefore, circRNAs may be a novel type ofdiagnostic marker and therapeutic target of cancers.
March 29, 2016; Accepted: September 1, 2016
Junming Guo. Tel/Fax: +86-574-87600758; E-mail: guojunming@nbu.edu.cn
circular RNA, microRNA sponge, tumor, occurrence
Supported by: Applied Research Project on Nonprofit Technology of Zhejiang Province (No. 2016C33177), Zhejiang Provincial High-Education Teaching Reform Project (No. jg2015047).
浙江省公益技术应用研究计划项目 (No. 2016C33177),浙江省高等教育教学改革项目 (No. jg2015047) 资助。
网络出版时间:2016-09-23 网络出版地址:http://www.cnki.net/kcms/detail/11.1998.Q.20160923.1114.001.html
猜你喜欢
现代实用医学(2022年10期)2022-12-08
钛工业进展(2022年2期)2022-05-13
少儿科学周刊·儿童版(2021年19期)2021-12-10
少儿科学周刊·儿童版(2021年19期)2021-12-10
中国民间疗法(2021年9期)2021-07-22
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
小学生必读(中年级版)(2018年5期)2018-08-15
作文教学研究(2016年1期)2016-07-05
医学研究杂志(2015年8期)2015-06-22
