
残膜对番茄苗期和开花坐果期生长的影响*
2016-12-10 03:21邹小阳牛文全刘晶晶官雅辉梁博惠郭丽丽张明智
中国生态农业学报(中英文) 2016年12期
邹小阳 牛文全** 刘晶晶 李 元 官雅辉 梁博惠 郭丽丽 张明智
残膜对番茄苗期和开花坐果期生长的影响*
邹小阳1牛文全1**刘晶晶2李 元3官雅辉1梁博惠2郭丽丽2张明智4
(1. 中国科学院水利部水土保持研究所 杨凌 712100; 2. 西北农林科技大学水利与建筑工程学院 杨凌 712100; 3. 西北农林科技大学水土保持研究所 杨凌 712100; 4. 河南省水利科学研究院 郑州 450002)
残膜污染已成为危害农田生态系统的重要因子, 不仅阻碍土壤水分和养分运移, 而且影响作物生长。为探明残膜对不同生育期番茄生长的影响规律, 采用小区试验方法, 设置0 kg∙hm-2、80 kg∙hm-2、160 kg∙hm-2、320 kg∙hm-2、640 kg∙hm-2和1 280 kg∙hm-2共6种不同残膜量水平, 研究残膜量对番茄苗期和开花坐果期的根系特征、地上部生长和干物质积累的影响, 并采用Logistic生长模型定量分析残膜对番茄养分积累始盛期、高峰期和盛末期的影响。结果表明, 残膜阻碍了番茄苗期和开花坐果期根系的生长, 根体积、根长密度和根干质量密度均随残膜量增加而降低; 随着残膜量增加, 苗期和开花坐果期的株高和茎粗均呈减小趋势, 且株高和茎粗的增长速率逐渐降低。番茄养分积累的始盛期和高峰期随残膜量增加而提前, 番茄追肥的最佳时期也应提前。在番茄苗期和开花坐果期, 根系、茎秆、花和幼果的干物质量均随残膜量增加而减少, 而叶片的干物质量呈递增趋势。残膜对番茄苗期根系、地上部生长和干物质积累的阻碍作用高于开花坐果期。由此可见, 残膜对番茄苗期的危害强于开花坐果期, 且干物质积累的始盛期和高峰期均随残膜量增加而提前, 加强番茄苗期水肥管理和提前水肥施加时间是减轻残膜危害的有利措施。
番茄 根系干物质积累 生长指标 残膜
地膜具有增温保墒、抑制土壤盐渍化、防治虫草害和改良土壤等效果[1-2], 是重要的农业生产资料。地膜覆盖技术已推广应用到棉、粮、瓜、果、茶等40余种农作物, 带来了农业生产力跨越式发展和生产方式的改变[3]。然而, 地膜在土壤中可残留200~400 a[4], 残膜量随覆膜年限的延长而增加。随着中国地膜使用量和覆膜面积的持续增长, 残膜对农业生态系统的危害逐年加剧, 残膜污染也成为限制农业可持续发展的瓶颈[5]。据调查, 土壤中残膜以小膜(<4 cm2)为主, 且具有分层性, 超过2/3的残膜分布在0~10 cm表土层[6]。残膜不仅破坏土壤孔隙连续性, 降低土壤过水断面积, 阻碍水分和养分运移[7-10], 而且阻碍作物根系生长, 使根系呈鸡爪型和丛生型等畸形, 影响作物生长发育[11]。
土壤中残留地膜, 必然会阻碍作物的生长发育。解红娥等[12]发现随残膜量增加, 小麦(L.)产量降低0.8%~22.1%, 玉米(L.)籽粒产量降低2.1%~27.5%, 棉花(Buch.)产量降低1%~7.5%。董合干等[13]研究发现, 当残膜量达1 000 kg∙hm-2, 棉花产量降低13.5%~18.1%, 成苗率下降13.0%~21.1%, 生物量、根系表面积和根冠比均明显降低。高玉山等[14]研究发现, 残膜会引起玉米出苗率降低、出苗时间推迟、根系伸长受阻, 进而影响植株地上部分的生长发育。辛静静等[15]基于大田试验, 发现玉米出苗率随残膜量增加而降低, 残膜对玉米的影响程度在苗期和拔节期最为明显。上述研究多集中于残膜对作物成熟期产量、株高和茎粗等方面的影响, 而关于残膜对作物不同生育期生长的影响研究较少。
在作物不同生育期内, 作物根系、地上部分和干物质积累对残膜的响应不同。根据不同生育期作物对残膜的响应差异, 探明作物需加强田间管理的时间段, 可有效减弱残膜对作物的危害。目前通过分析作物不同生育期对某一因素的响应差异, 从而制定适宜的田间管理措施的研究较多。如韩凡香等[16]研究了玉米秸秆带状覆盖对马铃薯(L.)生育前期和生育中后期生长的影响, 发现该措施可显著促进旱地马铃薯的生长。崔宁博等[17]研究了不同生育期水分亏缺对梨枣(Mill.)品质的影响, 发现果实成熟期中度亏水有利于改善梨枣品质。而关于残膜对作物不同生育期生长的影响研究较少。因此, 本文设置不同残膜量水平, 分析不同残膜量对番茄(Miller.)苗期和开花坐果期根系特征、地上部分生长和干物质积累的影响, 揭示残膜对作物不同生育期生长的影响差异, 为残膜污染区制定适宜的田间管理提供理论依据。
1 材料与方法
1.1 试验地点与材料
试验于2015年10月2日至2016年1月22日在陕西省杨凌西北农林科技大学中国旱区节水农业研究院的日光温室(108°04¢E, 34°20¢N, 海拔521 m)内进行。试验区属暖温带半干旱半湿润气候, 年均辐射总量475.8 kJ·m-2, 年均日照2 163.8 h, 无霜期221 d, 年均温13 ℃, 年均蒸发量约1 400 mm。试验温室结构为房脊型, 南北走向(长13.5 m×宽6.5 m×脊高3.8 m)。温室顶部用透光率较高的防水无滴膜覆盖, 番茄生育期内温室平均气温和湿度分别为21.82℃和50.72%。试验土壤为砂质黏壤土, 0~50 cm土层内, 黏粒(<0.001 mm)15.3%, 粉砂粒(0.001~0.05 mm) 62.1%, 砂粒(0.05~2 mm)22.6%。土壤pH为7.23, 电导率为0.24 dS·m-1。土壤初始质量含水率为20.16% (烘干法), 容重为1.36 g·cm-3(环刀法)。土壤全氮含量、碱解氮含量、有机质含量、全钾含量和速效磷含量分别为0.88 g·kg-1、32.51 mg·kg-1、7.67 g·kg-1、22.3 g·kg-1和12.87 mg·kg-1, 土壤肥力中等。
供试番茄品种为‘海地’, 2015年9月12日通过穴盘育苗, 20 d秧龄时(2015年10月2日)选取生长势一致的幼苗移栽。试验小区(图1)总面积为75.6 m2(6.0 m×12.6 m), 通过土垄(高10 cm, 长1 260 cm, 上宽10 cm, 下宽20 cm)将小区等分为6个试验区(1.0 m×12.6 m), 每个小区内再均分为3个微小区(1.0 m×4.2 m), 作为3个重复。并在土垄两侧埋设深50 cm的防水帆布, 消除各小区间水肥相互渗透和根系缠绕的影响。种植株距为60 cm, 每个微小区种植6株生长势一致的幼苗, 各处理的番茄总数均为18株。移栽前施用200 kg·hm-2氮肥(N含量46.4%), 150 kg·hm-2磷肥(P含量12%)和300 kg·hm-2钾肥(KCl含量24%)作为基肥。并在移栽前2 d灌溉足量的保苗水。番茄苗期和开花坐果期施用的肥料为丰得利水溶肥(总氮20%, 总磷20%, 总钾20%, 微量元素≥0.5%)。供试的微薄白色地膜厚度为0.008 mm, 拉伸负荷≤2.7 N, 断裂标称应变≥300 N, 直角撕裂负荷≥1.4 N, 总灰分≤0.5%, 拉伸负荷较低, 以模拟田间易破碎残膜。
1.2 试验设计
土壤中大部分残膜为小膜(<4 cm2)[3], 为消除残膜形状大小对作物根系生长的影响, 本试验统一使用1 cm×2 cm的长方形残膜。已有研究发现[3,11,13,18], 在不进行残膜清理的条件下, 残膜量M(kg∙hm-2)与覆膜年限(a)之间存在线性关系:M=5.546+47.840 (2=0.871)。根据该方程, 预测长期覆膜耕作对作物生长发育的影响, 设定6个残膜梯度, 分别为0 kg∙hm-2、80 kg∙hm-2(覆膜6 a左右)、160 kg∙hm-2(覆膜20 a左右)、320 kg∙hm-2(覆膜49 a左右)、640 kg∙hm-2(覆膜107 a左右)和1 280 kg∙hm-2(覆膜222 a左右), 用T0、T1、T2、T3、T4和T5表示。各处理田间布设及重复见图1。
图中Ⅰ、Ⅱ和Ⅲ分别代表各处理的3个微小区。T0、T1、T2、T3、T4和T5示残膜量分别为0 kg∙hm-2、80 kg∙hm-2(覆膜6 a左右)、160 kg∙hm-2(覆膜20 a左右)、320 kg∙hm-2(覆膜49 a左右)、640 kg∙hm-2(覆膜107 a左右)和1 280 kg∙hm-2(覆膜222 a左右)处理。In this picture, the symbols of Ⅰ,Ⅱand Ⅲ represent three micro plots of each treatment, respectively. T0, T1, T2, T3, T4 and T5 are treatments of residual plastic film amounts of 0 kg∙hm-2, 80 kg∙hm-2, 160 kg∙hm-2, 320 kg∙hm-2, 640 kg∙hm-2and 1 280 kg∙hm-2, which respectively represent plastic film mulching for 0 year, 6 years, 20 years, 49 years, 107 years and 222 years.
各小区所需基肥和残膜通过YJB4型移动式搅拌机(江阴市永宏化工机械有限公司)进行充分混合,转速控制为100 r·min-1, 搅拌30 min。搅拌后, 将肥料和残膜混合物均匀地撒在各小区。番茄生育期按照联合国粮农组织推行的FAO-56标准划分为苗期(2015年10月2日—11月20日)和开花坐果期(2015年11月20日—2016年1月22日)。在生长期内, 每株番茄间隔4 d灌溉2 L水, 每隔8 d在灌溉水中添加4 g水溶肥, 均灌溉在植株基部。在番茄生长期内及时除草, 防治病虫害, 其他田间管理措施与当地习惯一致。苗期末和开花坐果期末在各微小区选取3株长势一致的番茄植株, 各处理共取9株, 进行根系特征和干物质积累量分析。
1.3 测定指标与方法
1.3.1 土壤质量含水率测定
每次灌水2 d后, 按0~10 cm、10~20 cm、20~30 cm和30~40 cm分层用土钻取土样, 各处理的6个土样采样点均分布于植株中间(图1), 土样装入铝盒中称鲜土质量, 在105 ℃下烘干, 按照公式计算土壤质量含水率: 土壤质量含水率(%)=(鲜土质量-干土质量)/干土质量×100。
1.3.2 根系采样及测定方法
在番茄苗期末和开花坐果期末, 采用沟壕法取出植株根样, 取样时间分别为2015年11月20日和2016年1月22日。以植株为中心, 在0~40 cm土层挖取0.064 m3(长40 cm, 宽40 cm, 深40 cm)土样, 低压水冲洗根样, 剔除枯枝落叶等杂质。用滤纸吸干根系表面水分, 按0~10 cm、10~20 cm、20~30 cm和30~40 cm分层, 用EPSON Perfection V700 Photo型扫描仪扫描各土层根样, 分辨率设为300 dpi, 扫描图像通过WinRHIZO软件分析得到根长和根体积等参数。
1.3.3 株高和茎粗测定
移栽前用卷尺和数显游标卡尺测量番茄株高和茎粗, 定植后, 每隔7 d测量株高和茎粗。株高用卷尺从植株靠近地表处起测量, 茎粗用数显游标卡尺在主茎基部测量。
1.3.4 各器官干物质积累量测定
在番茄苗期末和开花坐果期末, 将样品植株分离为根、茎、叶、花和幼果, 在105 ℃下杀青30 min后, 于80 ℃下烘干至恒重, 用电子称(精度0.01 g)称量各器官干重。
1.3.5 根干质量密度和根长密度的计算
根干质量密度(m)和根长密度(l)计算公式分别为:

(2)
式中:m为0~40 cm土层的根干质量密度(g·m-3);l为0~40 cm土层的根长密度(m·m-3);为0~40 cm土层的根系干重(g);为0~40 cm土层体积(cm3), 本文为0.064 m3;为0~40 cm土层根系长度(cm)。
1.4 Logistic生长模型
生长模型广泛应用于描述和预测生物个体生长发育及经济特性, 多数作物的生长符合Logistic生长模型, 该模型的生长曲线呈“S”型, 适用于描述各类作物的生长发育过程[19]。该模型的表达式为[20]:

式中:为番茄各项生长指标的估计值,为番茄各指标可能达到的最大值,为番茄植株生长初期的状态参数,为番茄的增长速率系数, e为自然对数的底数,为番茄植株的发育时间(d)。
根据Logistic方程解析原理, 对公式(3)求一阶导数, 求得番茄的生长速率函数R(t):

当=ln(/),=/2时, 番茄的生长速率R达到最大值MGR:
MGR=´/4 (5)
对公式(4)再求二阶导数可得公式(6):

令公式(6)等于0, 得到番茄生长速率函数的两个拐点1和2。求得1=(ln1.317)/和2=(ln+ 1.317)/。
当番茄生长速率R达到最大值时, 番茄的发育时间记为max, 即max=ln/。max、1和2为Logistic生长模型的3个关键时间点, 分别为番茄养分积累的始盛期、高峰期和盛末期[20], 即番茄在0—1为养分渐增期,1—2为养分陡增期,2—番茄成熟期为养分缓增期, 番茄在max时期养分利用效率最高, 生长速率最快。因此, 番茄在1和max时间点加强水肥管理, 可实现番茄的高产稳产。
1.5 数据处理与分析方法
采用Microsoft Excel 2010软件进行绘图, Origin Pro 9.0进行函数拟合, Curve Expert 1.3 软件进行Logistic生长模型拟合, AutoCAD 2009软件绘制试验区示意图, SPSS 19.0软件进行方差分析和多重比较(LSD法)。
2 结果与分析
2.1 残膜对番茄根系生长的影响
2.1.1 根体积
不同残膜量对番茄苗期和开花坐果期根体积的影响结果表明(图2), 在番茄苗期, 残膜量≤320 kg·hm-2处理之间无显著差异(=0.73,>0.05), 残膜量≥640 kg·hm-2处理的根体积显著低于其他处理(=4.94,<0.05)。含残膜处理的苗期根体积较T0降低了0.35%、0.99%、1.92%、3.20%和4.17%。开花坐果期的根体积随残膜量增加而降低, 含残膜处理较对照处理减少了0.03%、0.17%、0.25%、0.69%和0.97%。开花坐果期的根系体积降低率明显低于苗期(=5.56,<0.05), 表明残膜对苗期根系生长的阻碍作用高于开花坐果期。
不同小写字母表示番茄苗期各处理间差异显著(<0.05), 不同大写字母表示番茄开花坐果期各处理间差异显著(<0.05);下同。In this picture, different lowercase letters indicate significant differences at 0.05 level among treatments at seedling stage, different capital letters indicate significant differences among different treatments at blooming and fruit-setting stage. The same below.
2.1.2 根长密度
由图3可知,含残膜处理的番茄苗期根长密度较对照处理分别降低2.59%、6.94%、9.16%、15.85%和24.67%,含残膜处理开花坐果期的根长密度比对照处理分别降低1.82%、5.29%、8.21%、13.85%和17.57%, 番茄苗期根长密度的降低率高于开花坐果期, 表明残膜对番茄苗期根系长度增长的阻碍作用强于开花坐果期。由图3还可知, 在番茄苗期, 残膜量≥320 kg·hm-2处理的根长密度明显低于其他处理(<0.05); 在番茄开花坐果期, 残膜量≥160 kg·hm-2处理的根长密度随残膜量增加呈显著下降趋势(=29.20,<0.05)。苗期和开花坐果期的根长密度均随残膜量增加而降低, 其原因是表土层残膜会阻滞土壤水分入渗, 使大部分水分通过大气蒸发转化为无效水, 降低了土壤有效水含量, 限制番茄根系的生长。
2.1.3 根干质量密度
在番茄苗期, 残膜量≤160 kg·hm-2的处理之间根干质量密度差异不显著(=1.69,>0.05); 残膜量为1 280 kg·hm-2时, 根干质量密度明显低于其他处理(=13.11,<0.05)。在番茄开花坐果期残膜量≥640 kg·hm-2处理显著降低(=31.12,<0.05)。苗期和开花坐果期的根干质量密度均随残膜量增加而降低, 含残膜处理的苗期根干质量密度较对照处理分别降低2.29%、8.67%、17.44%、23.92%和31.59%; 含残膜处理的开花坐果期根干质量密度较对照处理降低1.72%、6.52%、13.14%、17.75%和24.17%; 含残膜处理苗期的根干质量密度降低率与开花坐果期的差值分别为0.57%、2.15%、4.3%、6.17%和7.42%, 差值均为正值, 且随残膜量增加而增大(图4)。表明残膜阻碍番茄苗期根系生物量积累的强度高于开花坐果期, 且阻滞作用随残膜量增加而增强。
2.2 残膜对番茄地上部分生长的影响
2.2.1 残膜对株高和茎粗的影响
不同残膜水平对番茄株高和茎粗的影响见图5。由图5a可知, 在苗期末, 含残膜处理的株高较T0分别降低了1.47%、6.62%、8.75%、15.84%和24.11%, 株高的降低率随残膜增多而增大。在开花坐果期末, 含残膜处理的株高为T0的0.94倍、0.93倍、0.75倍、0.67倍和0.59倍, 番茄开花坐果期株高均随残膜量增加而降低。由图5b可知, 在番茄苗期末, 含残膜处理的茎粗较T0分别降低了5.73%、16.41%、38.55%、45.04%和57.63%; 开花坐果期末, 含残膜处理的茎粗较T0分别减少了1.97%、6.90%、13.14%、23.11%和30.45%, 番茄苗期和开花坐果期的株高和茎粗均随残膜量增加而降低。
株高增长速率与残膜量呈负相关关系(图5c), 含残膜处理的苗期株高增长速率较T0分别降低了0.99%、10.60%、14.24%、23.51%和34.77%, 残膜量≥640 kg·hm-2处理的株高增长速率明显降低(<0.05); 开花坐果期的株高增长速率较T0分别降低了6.28%、7.01%、43.96%、48.55%和59.18%, T5处理的株高增长速率明显低于其他处理(=11.25,<0.05), 番茄苗期和开花坐果期株高的增长速率均随残膜量增加而降低。在番茄开花坐果期, 苗期和开花坐果期的茎粗增长速率最大值分别为0.328 mm·week-1和0.443 mm·week-1,最小值分别为0.139 mm·week-1和0.258 mm·week-1, 苗期和开花坐果期的茎粗增长速率均随残膜量增加而降低(图5d)。试验结果表明, 残膜对作物地上部分生长的阻碍作用随残膜量增加而增强。
番茄株高(cm)和茎粗(mm)通过Logistic生长模型进行拟合, 拟合结果如表1所示。

表1 不同残膜量处理间番茄株高和茎粗的Logistic生长模型拟合参数
由表1可知, 各处理的决定系数2均大于0.87, 番茄苗期和开花坐果期的株高和茎粗均符合Logistic生长模型。参数值随残膜量增加而减小, 说明番茄株高可能达到的最大值随残膜增多呈减小趋势。参数值随残膜增加而降低, 表明残膜对番茄苗期植株生长产生阻碍作用。参数为番茄的增长速率系数, 各处理的值随残膜量增加逐渐减小, 且最大生长速率MGR也呈减小趋势, 表明番茄的生长势随残膜量增加而减弱, 残膜对番茄生长的阻碍作用随残膜增多而增强。
max、1和2分别为养分积累始盛期、高峰期和盛末期的时间节点。从番茄株高的角度考虑, 含残膜处理的max值较T0分别减少了1.29 d、2.91 d、5.94 d、8.27 d和12.35 d, 说明番茄养分积累的始盛期随残膜量增加而提前。含残膜处理的1较T0分别降低了4.67%、16.18%、29.81%、39.63%和57.46%(表1), 番茄的养分高峰期随残膜量增加而提前。2呈先减后增的趋势, 当残膜量≥160 kg×hm-2时,2值随残膜量增加而递减(表1)。对于番茄茎粗,max和1值均随残膜量增加而减小, 表明养分积累始盛期和高峰期均随残膜量增加而提前。对番茄加强水肥管理的最佳时期是max和1, 由上述结果可知, 随着残膜量增加, 番茄养分积累的始盛期和高峰期逐渐提前, 因此番茄追肥的最佳时期也应提前。
2.2.2 残膜对干物质积累的影响
由图6a可知, 苗期干物质积累量随残膜量增加而降低, 含残膜处理的总干物质积累量较T0分别降低了1.48%、5.37%、10.74%、18.26%和27.11%。残膜量≤160kg·hm-2处理之间根系、茎秆和叶片的干物质量均无显著差异(>0.05); 残膜量≥320 kg·hm-2时, 番茄茎秆的干物质量随残膜量增加显著降低(=19.87,<0.05), 而叶片干物质量降低趋势不明显(=1.45,>0.05)。番茄苗期根系和茎的干物质量均随残膜量增加而递减, 而叶片的干物质量逐渐递增。由图6b可知, 开花坐果期的干物质积累量与残膜量呈负相关关系, 残膜量≥160kg·hm-2处理的干物质量明显低于其他处理(=66.145,<0.05)。含残膜处理的总干物质积累量较T0降低了1.24%、4.04%、7.75%、11.53%和18.37%, 开花坐果期总干物质积累量的降低率低于苗期, 说明残膜对番茄苗期干物质积累的阻碍作用强于开花坐果期。番茄开花坐果期根系、茎秆、花和幼果的干物质量均随残膜量增加而减少, 而叶片的干物质量呈递增趋势。
不同字母表示各处理间根、茎、叶、花和幼果的干物质量差异显著(<0.05)。In this picture, different letters indicate significant differences in dry matter accumulation of root, stem, leaf, flower and young fruit among different treatments at 0.05 level.
3 讨论
3.1 残膜对番茄根系生长的影响
残膜作为土壤的外源物质, 会堵塞土壤水分运移通道, 降低土壤孔隙度和透气性, 阻碍水分下渗和水平扩散, 引起土壤板结[8]; 且残膜是一种柔韧性较强的物质, 易缠绕在作物根系周围, 阻碍土壤与大气之间的气体交换, 恶化土壤水热状况, 减弱微生物分解作用和土壤矿化作用, 影响作物根系的养分吸收和呼吸作用[21-22]。
本研究发现番茄的根体积、根长密度和根干质量密度均随残膜量增加而减小。这是因为在番茄苗期, 0~10 cm土层质量含水率随残膜量增加而增大(图7), 表土层水分含量越高, 土壤通气状况越差, 使土壤中CO2逐渐累积, 阻碍根系的有氧呼吸, 原有的有机同化物大量被消耗, 阻碍了根系生长发育[23]。张静等[24]研究发现, 稻田土壤表层的O2浓度随表土含水率的增加而减少, 使CO2浓度逐渐上升, 阻碍水稻根系的生长。Cook等[25]研究发现, 土壤含水率对土壤孔隙度和土壤通透性影响很大, 较高含水率使土壤中CO2逐渐积聚, 抑制好氧微生物生理活动和根系的呼吸作用, 威胁作物生长。可见, 残膜主要通过改变土壤水分状况、根系呼吸作用和土壤水热条件来影响作物根系的生长发育。
土壤水分与根系及地上部分生长发育紧密相关, 根据图7所测的土壤质量含水率数据, 对不同土层质量含水率与番茄地上和地下各项生长指标进行相关分析(表2)。0~10 cm土层质量含水率与番茄的生长指标呈极显著负相关(<0.01), 表明在含残膜的条件下, 0~10 cm土层质量含水率越高越不利于番茄的生长。本研究发现, 随着残膜量增加, 0~10 cm土层质量含水率逐渐上升(图7), 而番茄根体积、根长密度和根干质量密度均呈递减趋势。其原因是残膜大部分分布在表土层, 阻碍了水分向深土层入渗, 使水分集中分布于0~10 cm; 在强烈的大气蒸发条件下, 灌溉水转化为无效水, 使番茄易产生生理干旱, 阻碍了番茄根系的生长。Kuster等[26]与王靖等[27]研究发现, 表土层水分大量无效蒸发, 对植物的根系生长产生不利影响, 导致根长、根表面积和根体积等减小。
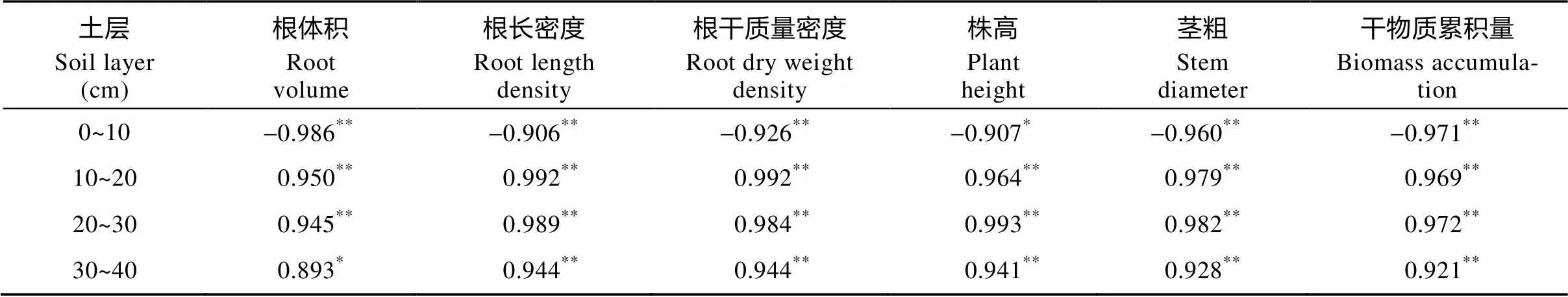
表2 各土层质量含水率与番茄植株各项生长指标的相关系数
*和**分别表示在0.05和0.01水平上显著相关。 * and ** represent significant differences at the 0.05, 0.01 probability levels, respectively.
根系的生长状况还与根区土壤含水率有关。番茄根系主要分布于10~40 cm 土层, 10~40 cm土层含水率与番茄的各项生长指标均呈显著正相关(<0.05)(表2), 10~40 cm土层含水率对番茄根系生长起明显的促进作用。而含残膜处理的10~40 cm土层平均质量含水率较T0分别降低了0.10%、0.26%、0.42%、0.60%和0.73%, 10~40 cm土层含水率随残膜量增加而降低, 表明番茄根区土壤含水率随残膜增多而减少。番茄根区土壤含水率对根系的生长起决定性作用, 番茄根系通过调整根系结构和空间分布来应对根区质量含水率的变化。薛丽华等[28]研究发现, 0~80 cm土层质量含水率随春季灌水量的减少而降低, 小麦根干质量密度、根长密度和根体积密度均显著减少。丁红等[29]研究0~40 cm土层水分亏缺对花生中后期根系生长的影响, 发现花生根系的根长密度、根表面积和根体积均随土壤含水率的降低而减少。本研究还发现残膜对番茄苗期根体积、根长密度和根干质量密度的阻碍作用强于开花坐果期, 因为苗期是植株生长发育过程中对土壤水分最敏感的时期, 且苗期根系的穿透能力较弱, 开花坐果期根系的吸水能力和穿透能力均强于苗期。苗期根系主要分布于0~10 cm土层, 而该土层含水率随残膜量增加而减少, 阻碍了根系的生长发育。马旭凤等[30]研究发现, 表土层水分含量降低, 使作物根系的中柱面积和导管直径减小, 导管壁变得不规整, 导致根系长度、根直径和根生物量降低。残膜不仅影响土壤水分的分布, 还会堵塞土壤孔隙, 增大作物根系穿透土壤下扎的阻力。残膜堵塞土壤孔隙的面积随残膜量增加而增大[9], 使作物根系的生存空间减少, 根系生长阻力增加; 而开花坐果期的根系活力和生长能力均优于苗期, 使残膜对苗期根系生长的阻碍作用强于开花坐果期。因此, 在番茄苗期应加强水肥管理, 减弱残膜对苗期番茄生长发育的危害。
3.2 残膜对番茄地上部分生长和干物质积累的影响
地上部分生长状况与根系生长紧密相关, 合理的根系结构是作物高产优质的生理基础。本研究发现番茄苗期和开花坐果期的株高、茎粗和干物质积累量均随残膜量增加而递减, 这是因为根系体积、根长密度和根干质量密度均随残膜量增加而减小, 减弱根系吸收水分和养分的能力, 降低水分向地上部分的传输, 使叶片相对含水量和叶绿素含量减少, 减弱光合作用强度[31], 进而影响地上部分生长和干物质量的积累。刘晚苟等[32]研究发现, 土壤中掺加固体物质会增大根系穿透土壤的阻力, 使作物根系平均直径逐渐增大, 横向导水阻力增大; 并加速根系内外皮层木质化和栓质化, 降低了根系的导水能力, 引起轻度的水分亏缺, 影响作物地上部分的生长发育。
植株地上部生长和干物质积累还与土壤水分含量相关,各处理0~40cm土层平均含水率分别为(21.859±0.878)%、(21.787±0.794)%、(21.676±0.680)%、(21.570±0.583)%、(21.494±0.477)%和(21.470±0.477)% (图7), 土壤含水率随残膜量增加而减小, 使含残膜处理的番茄形成水分亏缺。且水分亏缺程度随残膜量增加而增大, 番茄叶片的总气孔密度和关闭气孔数逐渐增加, 气孔的开张度逐渐变小,导致CO2亏缺, 减弱了番茄的光合作用, 使番茄干物质积累量减少[33]。残膜引起的水分亏缺还会导致根系脱落酸(ABA)产生累积效应, 影响叶片气孔开度, 降低根系吸水能力, 抑制番茄生长及同化物的合成与积累[34]。光合作用产物是干物质形成的基础, 水分亏缺会降低叶片叶绿素含量, 减少叶片总超氧化物歧化酶和过氧化氢酶的含量, 缩短番茄叶片和茎等器官的功能期, 减弱光合作用, 阻碍了光合作用产物的积累和转运, 引起干物质积累量的减少[35]。在含残膜的条件下, 根区土壤质量含水率随残膜量增加而递减, 加速了叶片淀粉的水解过程, 使糖类物质逐渐累积, 光合产物的运输过程减缓, 引起番茄呼吸作用和光合作用之间碳源供应的失衡[36], 导致番茄株高、茎粗和干物质积累量随残膜量增加而减少。
本研究还发现叶片的干物质量随残膜增多而增加。这是因为土壤质量含水率随残膜量增加而减少, 而番茄生长对水分亏缺较为敏感, 残膜会促使根系合成ABA, 并转运至番茄地上部分调控植物的生长和物质分配, 实现在水分亏缺条件下的最优生长, 减少生长冗余并调节叶片生长与茎秆生长的平衡[37], 提高番茄叶片面积, 使番茄叶片干物质量呈增加趋势。同时本试验采用周期性灌水方式, 番茄在生长期内经历多次复水过程, 使番茄的生长补偿效应更为显著, 促使番茄叶片质量和叶面积的增大, 干物质逐渐向叶片分配, 使叶片干物质量随残膜增多而增加[38]。王利彬等[39]研究发现, 作物经过多次复水过程, 植株内的丙二醛和脯氨酸等物质逐渐下降, 叶面积和叶片生物量出现短暂的快速增长过程。
本研究通过小区试验研究了残膜对番茄苗期和开花坐果期的根系特征、生长状况和干物质积累量的影响, 对残膜污染区制定合理的田间管理措施具有重要指导意义。但残膜长期残留在土壤中并参与土壤各项生物地球化学循环, 今后应加强残膜对不同作物生长发育的长期影响研究。
4 结论
1)番茄根体积、根长密度和根干质量密度均随着残膜量增加而减小, 残膜对番茄苗期根系生长的阻碍作用强于开花坐果期, 应加强番茄苗期的水肥管理。
2)苗期和开花坐果期的株高和茎粗均随着残膜量增加而减小, 且株高和茎粗的增长速率呈递减趋势, 残膜对番茄苗期地上部生长的阻碍作用强于开花坐果期。Logistic生长模型对残膜量≤1 280 kg×hm-2处理的番茄生长过程模拟效果较好, 番茄养分积累的始盛期和高峰期随残膜量增加而提前, 番茄追肥的最佳时期也应提前。
3)苗期和开花坐果期的总干物质量均随残膜量增加而降低。在番茄苗期和开花坐果期, 根系、茎秆、花和幼果的干物质量均随残膜量增加而减少, 而叶片的干物质量呈递增趋势。残膜阻碍番茄苗期干物质积累的作用强度高于开花坐果期, 为减轻残膜对番茄的危害, 需加强番茄苗期的水肥管理。
References
[1] 龚雪文, 李仙岳, 史海滨, 等. 番茄、玉米套种膜下滴灌条件下农田地温变化特征[J]. 生态学报, 2015, 35(2): 489–496 Gong X W, Li X Y, Shi H B, et al. The interplanting between tomato and maize of soil temperature dynamics under mulched drip irrigation[J]. Acta Ecologica Sinica,2015, 35(2): 489–496
[2] 杨书运, 江昌俊. 稻草和地膜覆盖对冬季茶园保温增温作用的研究[J]. 中国生态农业学报, 2010, 18(2): 327–333 Yang S Y, Jiang C J. Effect of straw and plastic film mulching on warming and insulation of tea plantation in winter[J]. Chinese Journal of Eco-Agriculture, 2010, 18(2): 327–333
[3] 严昌荣, 梅旭荣, 何文清, 等. 农用地膜残留污染的现状与防治[J]. 农业工程学报, 2006, 22(11): 269–272 Yan C R, Mei X R, He W Q, et al. Present situation of residue pollution of mulching plastic film and controlling measures[J]. Transactions of the CSAE, 2006, 22(11): 269–272
[4] 严昌荣, 刘恩科, 舒帆, 等. 我国地膜覆盖和残留污染特点与防控技术[J]. 农业资源与环境学报, 2014, 31(2): 95–102 Yan C R, Liu E K, Shu F, et al. Review of agricultural plastic mulching and its residual pollution and prevention measures in China[J]. Journal of Agricultural Resources and Environment, 2014, 31(2): 95–102
[5] 张丹, 胡万里, 刘宏斌, 等. 华北地区地膜残留及典型覆膜作物残膜系数[J]. 农业工程学报, 2016, 32(3): 1–5 Zhang D, Hu W L, Liu H B, et al. Characteristics of residual mulching film and residual coefficient of typical crops in North China[J]. Transactions of the CSAE, 2016, 32(3): 1–5
[6] 马辉, 梅旭荣, 严昌荣, 等. 华北典型农区棉田土壤中地膜残留特点研究[J]. 农业环境科学学报, 2008, 27(2): 570–573 Ma H, Mei X R, Yan C R, et al. The residue of mulching plastic film of cotton field in North China[J]. Journal of Agro-Environment Science, 2008, 27(2): 570–573
[7] 李仙岳, 史海滨, 吕烨, 等. 土壤中不同残膜量对滴灌入渗的影响及不确定性分析[J]. 农业工程学报, 2013, 29(8): 84–90 Li X Y, Shi H B, Lü Y, et al. Effects of different residual plastic film quantities in soil on drip infiltration and its uncertainty analysis[J]. Transactions of the CSAE, 2013, 29(8): 84–90
[8] 王志超, 李仙岳, 史海滨, 等. 残膜埋深对滴灌条件下粉砂壤土水分入渗影响的试验研究[J]. 土壤, 2014, 46(4): 710–715 Wang Z C, Li X Y, Shi H B, et al. Study on effect of plastic film residue at different buried depths on silt loam drip infiltration[J]. Soils, 2014, 46(4): 710–715
[9] 王志超, 李仙岳, 史海滨, 等. 农膜残留对土壤水动力参数及土壤结构的影响[J]. 农业机械学报, 2015, 46(5): 101–106 Wang Z C, Li X Y, Shi H B, et al. Effects of residual plastic film on soil hydrodynamic parameters and soil structure[J]. Transactions of the CSAM, 2015, 46(5): 101–106
[10] 李元桥, 何文清, 严昌荣, 等. 点源供水条件下残膜对土壤水分运移的影响[J]. 农业工程学报, 2015, 31(6): 145–149 Li Y Q, He W Q, Yan C R, et al. Effect of residual film on soil infiltration under drip irrigation[J]. Transactions of the CSAE, 2015, 31(6): 145–149
[11] 刘建国, 李彦斌, 张伟, 等. 绿洲棉田长期连作下残膜分布及对棉花生长的影响[J]. 农业环境科学学报, 2010, 29(2): 246–250 Liu J G, Li Y B, Zhang W, et al. The distributing of the residue film and influence on cotton growth under continuous cropping in oasis of Xinjiang[J]. Journal of Agro- Environment Science, 2010, 29(2): 246–250
[12] 解红娥, 李永山, 杨淑巧, 等. 农田残膜对土壤环境及作物生长发育的影响研究[J]. 农业环境科学学报, 2007, 26(S1): 153–156 Xie H E, Li Y S, Yang S Q, et al. Influence of residual plastic film on soil structure, crop growth and development in fields[J]. Journal of Agro-Environment Science, 2007, 26(S1): 153–156
[13] 董合干, 刘彤, 李勇冠, 等. 新疆棉田地膜残留对棉花产量及土壤理化性质的影响[J]. 农业工程学报, 2013, 29(8): 91–99 Dong H G, Liu T, Li Y G, et al. Effects of plastic film residue on cotton yield and soil physical and chemical properties in Xinjiang[J]. Transactions of the CSAE, 2013, 29(8): 91–99
[14] 高玉山, 孙云云, 窦金刚, 等. 残膜对玉米出苗及根系伸长的研究[J]. 吉林农业科学, 2013, 38(6): 22–24 Gao Y S, Sun Y Y, Dou J G, et al. Effects of plastic film residue on emergence and root growth of corn seedlings[J]. Journal of Jilin Agricultural Sciences, 2013, 38(6): 22–24
[15] 辛静静, 史海滨, 李仙岳, 等. 残留地膜对玉米生长发育和产量影响研究[J]. 灌溉排水学报, 2014, 33(3): 52–54 Xin J J, Shi H B, Li X Y, et al. Effects of plastic film residue on growth and yield of maize[J]. Journal of Irrigation and Drainage, 2014, 33(3): 52–54
[16] 韩凡香, 常磊, 柴守玺, 等. 半干旱雨养区秸秆带状覆盖种植对土壤水分及马铃薯产量的影响[J]. 中国生态农业学报, 2016, 24(7): 874–882 Han F X, Chang L, Chai S X, et al. Effect of straw strip covering on ridges on soil water content and potato yield under rain-fed semiarid conditions[J]. Chinese Journal of Eco-Agriculture, 2016, 24(7): 874–882
[17] 崔宁博, 杜太生, 李忠亭, 等. 不同生育期调亏灌溉对温室梨枣品质的影响[J]. 农业工程学报, 2009, 25(7): 32–38 Cui N B, Du T S, Li Z T, et al. Effects of regulated deficit irrigation at different growth stages on greenhouse pear-jujube quality[J]. Transactions of the CSAE, 2009, 25(7): 32–38
[18] 董合干, 王栋, 王迎涛, 等. 新疆石河子地区棉田地膜残留的时空分布特征[J]. 干旱区资源与环境, 2013, 27(9): 182–186 Dong H G, Wang D, Wang Y T, et al. Spatial and temporal distribution characteristics of mulch residues in cotton field in Shihezi, Xinjiang[J]. Journal of Arid Land Resources and Environment, 2013, 27(9): 182–186
[19] 钱永强, 孙振元, 韩蕾, 等. 基于Logistic与Gompertz非线性模型的野牛草克隆生长模拟与分析[J]. 中国农业科学, 2011, 44(11): 2252–2259 Qian Y Q, Sun Z Y, Han L, et al. Simulation of clonal growth curve based on nonlinear models of Logistic and Gompertz for buffalograss [(Nutt.) Engelm Texoka][J]. Scientia Agricultura Sinica, 2011, 44(11): 2252–2259
[20] 王贺, 白由路, 杨俐苹, 等. 利用有效积温建立夏玉米追肥时期决策模型[J]. 中国生态农业学报, 2012, 20(4): 408–413 Wang H, Bai Y L, Yang L P, et al. A summer maize dressing decision-making model based on effective accumulated temperature[J]. Chinese Journal of Eco-Agriculture, 2012, 20(4): 408–413
[21] 赵亚丽, 薛志伟, 郭海斌, 等. 耕作方式与秸秆还田对土壤呼吸的影响及机理[J]. 农业工程学报, 2014, 30(19): 155–165 Zhao Y L, Xue Z W, Guo H B, et al. Effects of tillage and crop residue management on soil respiration and its mechanism[J]. Transactions of the CSAE, 2014, 30(19): 155–165
[22] Nosalewicz A, Lipiec J. The effect of compacted soil layers on vertical root distribution and water uptake by wheat[J]. Plant and Soil, 2014, 375(1/2): 229–240
[23] Gupta B, Shah D O, Mishra B, et al. Effect of top soil wettability on water evaporation and plant growth[J]. Journal of Colloid and Interface Science, 2015, 449: 506–513
[24] 张静, 刘娟, 陈浩, 等. 干湿交替条件下稻田土壤氧气和水分变化规律研究[J]. 中国生态农业学报, 2014, 22(4): 408–413 Zhang J, Liu J, Chen H, et al. Change in soil oxygen and water contents under alternate wetting and drying in paddy fields[J]. Chinese Journal of Eco-Agriculture, 2014, 22(4): 408–413
[25] Cook F J, Orchard V A. Relationships between soil respiration and soil moisture[J]. Soil Biology and Biochemistry, 2008, 40(5): 1013–1018
[26] Kuster T M, Arend M, Günthardt-Goerg M S, et al. Root growth of different oak provenances in two soils under drought stress and air warming conditions[J]. Plant and Soil, 2013, 369(1/2): 61–71
[27] 王靖, 李湘阁, 刘恩民, 等. 华北平原冬小麦相对蒸散与叶面积指数及表层土壤含水量的关系[J]. 中国生态农业学报, 2003, 11(2): 32–34 Wang J, Li X G, Liu E M, et al. The relationship between relative evapotranspiration and leaf area index and surface soil water content in winter wheat field of North China Plain[J]. Chinese Journal of Eco-Agriculture, 2003, 11(2): 32–34
[28] 薛丽华, 段俊杰, 王志敏, 等. 不同水分条件对冬小麦根系时空分布、土壤水利用和产量的影响[J]. 生态学报, 2010, 30(19): 5296–5305 Xue L H, Duan J J, Wang Z M, et al. Effects of different irrigation regimes on spatial-temporal distribution of roots, soil water use and yield in winter wheat[J]. Acta Ecologica Sinica, 2010, 30(19): 5296–5305
[29] 丁红, 张智猛, 戴良香, 等. 干旱胁迫对花生生育中后期根系生长特征的影响[J]. 中国生态农业学报, 2013, 21(12): 1477–1483 Ding H, Zhang Z M, Dai L X, et al. Effects of drought stress on root growth characteristics of peanut during mid-to-late growth stages[J]. Chinese Journal of Eco-Agriculture, 2013, 21(12): 1477–1483
[30] 马旭凤, 于涛, 汪李宏, 等. 苗期水分亏缺对玉米根系发育及解剖结构的影响[J]. 应用生态学报, 2010, 21(7): 1731–1736 Ma X F, Yu T, Wang L H, et al. Effects of water deficit at seedling stage on maize root development and anatomical structure[J]. Chinese Journal of Applied Ecology, 2010, 21(7): 1731–1736
[31] 杨婷, 谢志霞, 喻琼, 等. 局部根系盐胁迫对冬小麦生长和光合特征的影响[J]. 中国生态农业学报, 2014, 22(9): 1074–1078 Yang T, Xie Z X, Yu Q, et al. Effects of partial root salt stress on seedling growth and photosynthetic characteristics of winter wheat[J]. Chinese Journal of Eco-Agriculture, 2014, 22(9): 1074–1078
[32] 刘晚苟, 山仑. 土壤机械阻力对玉米根系导水率的影响[J]. 水利学报, 2004, 35(4): 114–117 Liu W G, Shan L. Effect of soil mechanical resistance on hydraulic conductivity of maize root[J]. Journal of Hydraulic Engineering, 2004, 35(4): 114–117
[33] 韩国君, 陈年来, 李金霞, 等. 番茄叶片气孔导度及渗透调节物质对快速水分胁迫的响应[J]. 中国生态农业学报, 2013, 21(9): 1100–1106 Han G J, Chen N L, Li J X, et al. Response of stomatal conductance and osmotic adjustment substances accumulation to rapid drough stress in tomato leaves[J]. Chinese Journal of Eco-Agriculture, 2013, 21(9): 1100–1106
[34] 孙宏勇, 张喜英, 陈素英, 等. 亏缺灌溉对冬小麦生理生态指标的影响及应用[J]. 中国生态农业学报, 2011, 19(5): 1086–1090 Sun H Y, Zhang X Y, Chen S Y, et al. Effects of deficit irrigation on physio-ecological indices of winter wheat[J]. Chinese Journal of Eco-Agriculture, 2011, 19(5): 1086–1090
[35] 王林林, 陈炜, 徐莹, 等. 氮素营养对小麦干物质积累与转运的影响[J]. 西北农业学报, 2013, 22(10): 85–89 Wang L L, Chen W, Xu Y, et al. Nitrogen nutrition affects dry matter accumulation and transportation in winter wheat[J]. Acta Agriculturae Boreli-occidentalis Sinica, 2013, 22(10): 85–89
[36] Flexas J, Bota J, Galmés J, et al. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress[J]. Physiologia Plantarum, 2006, 127(3): 343–352
[37] 谭念童, 林琪, 姜雯, 等. 限量灌溉对旱地小麦旗叶光合特性日变化和产量的影响[J]. 中国生态农业学报, 2011, 19(4): 805–811 Tan N T, Lin Q, Jiang W, et al. Effect of limited irrigation on diurnal variation in flag-leaf photosynthesis and yield of dryland wheat[J]. Chinese Journal of Eco-Agriculture, 2011, 19(4): 805–811
[38] Acevedo E, Hsiao T C, Henderson D W. Immediate and subsequent growth responses of maize leaves to changes in water status[J]. Plant Physiology, 1971, 48(5): 631–636
[39] 王利彬, 祖伟, 董守坤, 等. 干旱程度及时期对复水后大豆生长和代谢补偿效应的影响[J]. 农业工程学报, 2015, 31(11): 150–156 Wang L B, Zu W, Dong S K, et al. Effects of drought stresses and times on compensation effect after re-watering in soybean[J]. Transactions of the CSAE, 2015, 31(11): 150–156
Effect of residual plastic film on growth of tomato at seedling and blooming and fruit-setting stages*
ZOU Xiaoyang1, NIU Wenquan1**, LIU Jingjing2, LI Yuan3, GUAN Yahui1,LIANG Bohui2, GUO Lili2, ZHANG Mingzhi4
(1. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling 712100, China; 2. College of Water Conservancy and Architectural Engineering, Northwest A&F University, Yangling 712100, China; 3. Institute of Soil and Water Conservation, Northwest A&F University, Yangling 712100, China; 4. Henan Province Water Conservancy Research Institute, Zhengzhou 450002, China)
Although plastic film mulching is critical for crop production, its widespread use has produced a great deal of residual plastic film and destroyed soil structure, damaged the physical structure of soil and seriously impeded the movement of soil water and solute as well as crop growth. In order to determine the negative effects of residual plastic film on growth of tomato (Miller) at seedling and blooming and fruit-setting stages, plot experiment was conducted with different amounts of residual plastic film in 2015–2016 in a solar greenhouse of the Key Laboratory of Agricultural Soil and Water Engineering in Arid Areas (108°02′E, 34°17′N). Six treatments, including residual plastic film amounts of 0 kg∙hm-2, 80 kg∙hm-2, 160 kg∙hm-2, 320 kg∙hm-2, 640 kg∙hm-2and 1 280 kg∙hm-2, were set in triplicate plots. The total area of plot filed was 75.6 m2(6.0 m × 12.6 m) with three replications for each treatment, with each micro-plot size of 1.0 m´4.2 m. The height and stem diameter of the tomato plants were collected during the period from October 2015 to January 2016 and analyzed and then fitted with nonlinear Logistic models to determine the effects of residual plastic film on the biomass accumulation at seedling stage, blooming and fruit-setting stage of tomato. During the experiment, root morphological characteristics, plant height, stem diameter and biomass accumulation of the tomato plants were measured. The results showed that residual plastic film hindered the growth of tomato root at both seedling and blooming and fruit-setting stages. Root volume, root length density and root dry weight density decreased with increasing residual plastic film amount. In addition, as the amount of plastic film residue increased, plant height and stem diameter decreased significantly (< 0.05). Besides, the rates of increase in plant height and stem diameter decreased with increasing residual plastic film amount. Logistic model accurately simulated the growth process of tomato in fields with residual plastic film less than 1 280 kg∙hm-2(2> 0.87). As the amount of residual plastic film increased, beginning period and full period of tomato biomass accumulation was ahead of time. This implied that the suitable time of fertilization of tomato should be brought forward. Overall, biomass accumulation decreased for the whole tomato growth period. Contrary to biomass accumulation of leaf, that of root, stem, flower and young fruit decreased with increasing residual plastic film amount. The hindering effects of residual plastic film on tomato growth and dry matter accumulation at seedling stage were higher than at blooming and fruit-setting stage. The early period and middle period of tomato biomass accumulation all occurred in advanced. In summary, strengthening water and fertilizer management at seedling stage and bringing forward the application of fertilizer and irrigation were recommended as the proper measures to weaken the detrimental effects of residual plastic film on tomato.
Tomato; Root; Dry matter accumulation; Growth parameter; Residual plastic film
10.13930/j.cnki.cjea.160567
S184; S154.4
A
1671-3990(2016)12-1643-12
2016-06-24 接受日期: 2016-09-10
* 国家科技支撑计划项目(2015BAD24B01)资助
**通讯作者:牛文全, 主要从事灌溉理论与节水技术研究。E-mail: nwq@vip.sina.com 邹小阳, 主要从事土壤水分运移研究。E-mail: zouxiaoyang14@mails.ucas.ac.cn
* The study was supported by the National Key Technology R&D Program of China (2015BAD24B01).
** Corresponding author, E-mail: nwq@vip.sina.com
Received Jun. 24, 2016; accepted Sep.10, 2016
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
今日农业(2021年9期)2021-11-26
今日农业(2021年8期)2021-07-28
今日农业(2020年23期)2020-12-15
今日农业(2020年16期)2020-12-14
河北果树(2020年2期)2020-01-09
今日农业(2019年11期)2019-08-15
今日农业(2019年11期)2019-08-15
今日农业(2019年12期)2019-08-13
今日农业(2019年4期)2019-01-04
