
药用植物三七对土壤中砷的吸收运转及形态转化
2016-12-09 09:06杜丽娟米艳华尹本林和丽忠叶艳萍
生态与农村环境学报 2016年6期
杜丽娟,米艳华①,陈 璐,尹本林,马 杰,2,和丽忠,叶艳萍
(1.云南省农业科学院质量标准与检测技术研究所,云南 昆明 650223;2.中国科学院地理科学与资源研究所,北京 100101)
药用植物三七对土壤中砷的吸收运转及形态转化
杜丽娟1,米艳华1①,陈 璐1,尹本林1,马 杰1,2,和丽忠1,叶艳萍1
(1.云南省农业科学院质量标准与检测技术研究所,云南 昆明 650223;2.中国科学院地理科学与资源研究所,北京 100101)
以栽培药用植物三七(Panaxnotoginseng)为主要研究对象,采用高效液相色谱-原子荧光联用(HPLC-HG-AFS)技术,研究了三七产地土壤及植株样品中砷(As)含量及其赋存的化学形态,以及三七对As的吸收运转及形态转化。结果表明,无机态的As(Ⅴ)和As(Ⅲ)是三七种植土壤中的主要赋存形态,且As(Ⅴ)含量远高于As(Ⅲ),约占总量的95%。三七植株中总As的分布规律表现为根条>剪口>主根>叶>茎,说明根部吸收是土壤As进入三七的主要途径;而三七植株中各形态As分布规律因植株部位的不同而存在较大差异, As(Ⅲ):叶>根条>剪口>茎>主根;As(Ⅴ):剪口>根条>主根>叶>茎;二甲基砷(MMA):根条>茎>剪口>叶>主根。降低三七种植土壤As的生物有效性,深入研究As(Ⅴ)、As(Ⅲ)和MMA在三七不同部位相互转化的调控机制是缓解三七As毒害的重要途径。
土壤;三七;砷形态;吸收运转
砷(As)是自然界中丰度排第20位的一种具有较强毒性的类金属元素,人体长期暴露在As环境中可导致皮肤癌和肾、肝、膀胱等内脏器官的癌变,被世界卫生组织(WHO)和美国环境保护署(USEPA)定级为一种已知人类致癌物质[1]。As的毒性与其赋存形态密切相关,不同形态As毒性相差甚远,一般无机态As的毒性强于有机态As[2]。为了适应As污染日益严重的生存环境,自然界中的生物在一定程度上进化形成了各种As解毒和忍耐机制[3]。随着食物As污染导致人类健康问题加剧,国内外许多专家对作物体内As代谢过程的研究越来越多,尽管微生物细胞内As解毒机制的研究已非常深入,但植物细胞中As的代谢机制仍不清楚。目前已有研究报道仅限于水稻,部分水生植物和蜈蚣草、拟南芥等As污染修复植物[4-5],而对于三七(Panaxnotoginseng)和茶叶等一些特殊作物,虽然产品中已存在As污染问题,但针对这些作物的污染机制等研究较少。
As在自然环境中无所不在,尤其是广泛存在于土壤中,土壤-植物转运系统是人类受As毒害的一个最基本途径。有研究证明,植物有助于降低As向其地上部分的转运,且不同植物对As的积累和转运机制不同,为了降低As对植物的毒害作用,植物可将吸收态As进行转化,如氧化、还原和甲基化等[6-8]。竹子的地上部存在着无机As转化为有机As的过程,在其地上部二甲基砷〔DMA,(CH3)2AsO(OH)〕是主要的As存在形态,占总As的13.9%~44.9%[9]。水稻地上部分也存在着无机As被甲基化、As(Ⅴ)被还原为As(Ⅲ)的过程,在无有机As的培养介质中生长的水稻体(尤其是籽粒)中,DMA也是As的主要存在形态之一[10]。也有研究表明,在许多植物的根际土壤和水体中没有As的甲基化产物,但在植物体内能检测到多种As的甲基化产物,如一甲基砷〔MMA,CH3AsO(OH)2〕和DMA[11]。拟南芥[12]、绒毛草[13]和水稻[14]中发现了某些植物蛋白具有As(Ⅴ)的还原能力,已确认由ACR2和Arath:CDC25基因控制,还原过程需要谷胱甘肽(GSH)以及谷氧还蛋白提供电子将As(Ⅴ)还原为As(Ⅲ)[15],在这一过程中会形成GSH-As的结合态。黄泽春等[16]采用EXAFS方法证实大叶井口边草存在与GSH结合的As,这在印度芥菜中也有报道[17],多数耐As植物叶片中GSH-As大量存在,部分植物则在根部大量存在,叶片和根部的还原作用强。
三七是我国名贵药材之一,具有散瘀止血和消肿定痛等功效。前期大量研究表明,三七产地土壤As污染和三七As超标现象日趋严重[18-19];由于产地土壤As含量过高,不仅导致三七主要药用部位的健康风险增加[20-21];还会对三七的药用质量安全和出口贸易造成较大影响。此外,祖艳群等[22]探讨了三七土壤As含量的空间分布特征及土壤理化性质对三七不同部位As含量的影响。笔者以采自人工种植区的药用植物三七为主要研究对象,采用高效液相色谱-原子荧光联用技术(HPLC-HG-AFS)研究了三七种植区土壤及植株样品中As的含量及赋存形态,系统分析了三七种植土壤和植株As含量特征及As形态分布规律,对土壤-三七系统中As的吸收运转以及As在三七体内的形态转化等进行探讨,以期为药用植物三七安全种植、控制As污染、降解As毒性以及提高产品质量提供重要的基础研究理论。
1 材料与方法
1.1 样品采集
云南省文山壮族苗族自治州位于云南省东南部,属于亚热带气候区,冬无严寒,夏无酷暑。该地区地处北纬22°40′~24°48′,东经103°35′~106°12′之间,平均海拔1 000~1 800 m,年平均气温19 ℃,年平均降水量779 mm,全年无霜期356 d,年日照时数2 228.9 h。该地区土壤有暗棕壤、棕壤、黄棕壤、黄壤、红壤、赤红壤、砖红壤、石灰岩和紫色土9个类型。
在该三七种植区随机选取10个三七种植地块,在每个地块随机采集10~20株三七样品,同时分别采集相对应的表层(0~20 cm)和深层(>20~40 cm)土壤5点混合样。取样区域见图1。混合土样采集后自然风干,剔除样品中植物根系、有机残渣以及可见侵入体,用研钵研磨,装入密封袋待测。
图1 三七采样点位置示意
新鲜三七采集后用自来水冲洗干净,洗去根部泥土,并用细毛刷清洗,再用去离子水冲洗。将三七植株分成主根、根条、剪口、茎和叶5个不同部位,冷冻干燥至恒重,在冷冻条件下研磨,保存于4 ℃条件下备用。
1.2 分析方法
1.2.1 总As分析方法
采用文献[23]中的方法进行消解。称取0.2 g土壤样品于50 mL锥形瓶中,加入HNO3和H2O2,在电热板上消解,消解完全后用超纯水定容至50 mL,使用HPLC-HG-AFS测定土壤中总As含量;称取0.5 g植物样品于50 mL锥形瓶中,加入HNO3和HClO4,在电热板上消解,消解完全后用超纯水定容至50 mL,使用HPLC-HG-AFS测定植物中总As含量。
1.2.2 As形态分析方法
(1)土壤样品
用万分之一天平准确称取0.5 g样品于100 mL三角瓶中,按1∶50的固液质量比,用1 mol·L-1硝酸溶液浸提,若样品中As含量较低,则适当提高称样量或减少浸提剂体积(如1∶12.5的固液质量比),将三角瓶置于电热板上,以120 ℃加热2 h,冷却后转入50 mL离心管以2 000 r·min-1离心15 min(离心半径为40.57 mm)。重复以上提取步骤3次,合并提取液,过0.45 μm孔径滤膜,用HPLC-HG-AFS测定土壤As含量。HPLC-HG-AFS测定条件为:选用阴离子交换柱(PRP-X100),以15 mmol·L-1磷酸溶液(pH值为6.0)作为流动相,流速为1.0 mL·min-1。
(2)植物样品
用万分之一天平准确称取1.0~2.0 g样品于100 mL三角瓶中,按1∶5~1∶1的固液质量比,用体积比为1∶9的甲醇水溶液浸提,在25 ℃条件下振荡(振荡速度为180 r·min-1)提取1 h,50 ℃条件下超声1 h,转入50 mL离心管以2 000 r·min-1离心15 min(离心半径为40.57 mm)。重复以上提取步骤3次,合并提取液,过0.45 μm孔径滤膜,用HPLC-HG-AFS测定植物As含量。HPLC-HG-AFS测定条件与土壤样品相同。
1.3 数据处理
采用Excel 2003软件对数据进行处理,采用SPSS 12.0软件对数据进行方差分析、相关性分析和显著性检验。
2 结果与分析
2.1 土壤中总As及As存在形态
三七作为一种多年生宿根性植物,对种植环境土壤条件有一定的要求,除酸白泥土和黏重土外,土质疏松、排灌方便的土壤均可种植[24],三七根系主要的分布区域为浅层土壤,因此2个层次土壤As形态特征存在明显差异。SADIQ[25]研究指出,土壤中As的存在形态主要为3价和5价的无机As,有机As含量很低,且主要以MMA和DMA形态存在。在氧化条件下,As(Ⅴ)为主要形态,As(Ⅴ)/As(Ⅲ)比值可达10~100[25-28]。
该研究中三七种植土壤样本包括了南方特有的红壤、砖红壤、黄壤和棕壤4种土壤类型。10个地块表层(0~20 cm)和深层土壤(>20~40 cm)总As和各形态As的分布及含量见表1。土壤样本中As含量总体特征表现为:表层和深层土壤中MMA和DMA均未检出,可能是其含量太低,小于仪器最低检出量;As(Ⅴ)和As(Ⅲ)是三七种植土壤中As赋存的主要形态,这与前人的研究结论[6-8,25,29]相似。2个层次土壤As分布存在一定差异,土壤As污染情况严重,总As和As(Ⅲ)含量在表层土壤中较高,As(Ⅴ)含量在深层土壤中较高。10个地块的表层土壤中总As最大值与最小值的比值为8.9,As(Ⅲ)含量最大值为最小值的22.0倍,As(Ⅴ)含量最大值仅为最小值的2.6倍;深层土壤中总As、As(Ⅴ)和As(Ⅲ)含量最大值与最小值的比值分别为4.8、4.8和5.6。
可以看出,各形态As中As(Ⅴ)占主导地位,这主要是因为土壤中存在Fe、Mn等氧化性强的元素,会把部分As(Ⅲ)氧化成As(Ⅴ)[30],且土壤中As(Ⅴ)化合物较易被磷酸溶液提取出来。As(Ⅲ)的毒性约是As(Ⅴ)的几十倍,有机As的毒性比无机As要小得多,土壤中As(Ⅲ)被氧化成As(Ⅴ)、无机As被甲基化成有机As都是有效降低As毒性的自然反应。
表1 土壤样本中总As和不同形态As含量
Table 1 Contents of total arsenic and different forms of arsenic in the soil samples mg·kg-1

As形态 平均值标准差最小值最大值表层土壤 总As33.560.4510.2490.84 As(Ⅲ)0.280.140.020.44 As(Ⅴ)10.373.046.3916.76深层土壤 总As25.6712.0210.2648.84 As(Ⅲ)0.290.130.090.50 As(Ⅴ)9.904.513.9318.76
2.2 三七植株中总As及As存在形态
三七对土壤中As的吸收不仅受污染来源、土壤基本性质和气候条件的影响,而且与三七本身的遗传特性、主动吸收功能、对元素的富集能力、根际分泌物、根际环境微生物的种类和数量有关,甚至不同基因型的三七对As的吸收和富集都有明显差异。对三七植株样本的主根、根条、剪口、茎和叶5个部位总As及各形态As含量的测定结果(图2)表明,三七所有部位均能检测到As(Ⅴ)、As(Ⅲ)和MMA,DMA则未检出。三七植株中总As的分布规律表现为根条>剪口>主根>叶>茎,与前人研究结论[6-8,29]相似。而三七植株中各形态As分布规律不一致,具体表现为:As(Ⅲ),叶>根条>剪口>茎>主根;As(Ⅴ),剪口>根条>主根>叶>茎;MMA,根条>茎>剪口>叶>主根。根条和剪口中As(Ⅴ)含量明显高于其他部位;As(Ⅲ)含量在主根、剪口和茎中的变化不大,而叶中As(Ⅲ)含量明显高于其他部位;根条中MMA含量明显高于其他部位。整个三七植株中未能检出DMA,可能是因为有一部分As储存在植物组织的细胞壁中,而在土壤中生长的多年生草本植物细胞壁较厚,导致甲醇/水溶液提取有机态As时提取率不高,使得DMA未能检出。附着在植物组织的细胞质和细胞器中的部分As参与了植物自身的新陈代谢,且各形态As在三七植株各部位存在迁移转化,迁移运转快慢不尽相同,使得三七植株各部位的As含量不同。笔者试验表明,三七根条中MMA含量较高,且在三七的某些品种中尤为突出,这与唐艳梅等[31]的研究结论不同,究其原因,可能是某些三七品种中As甲基化作用较明显,无机态向有机态转化的As含量较高,但具体原因还需要进一步研究。
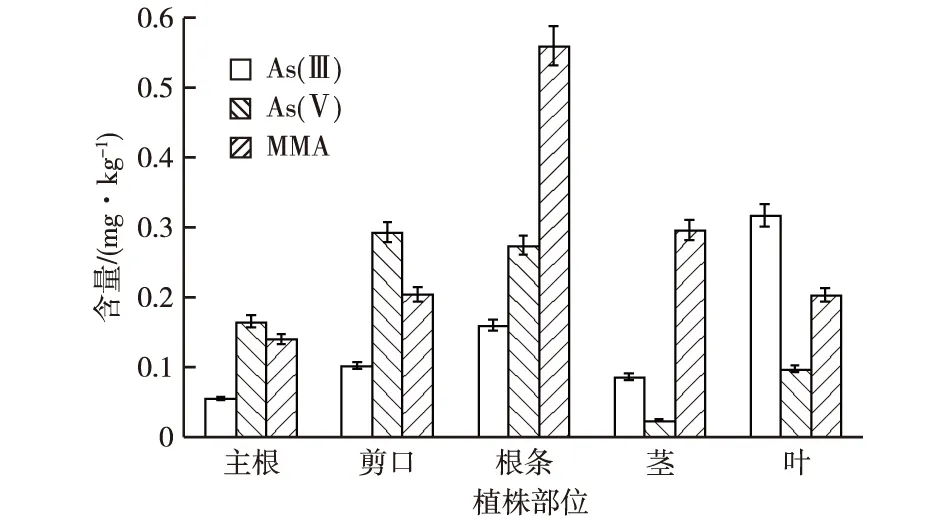
图2 三七植株各部位不同形态As含量
从同一植株部位各形态As含量的比值分析,根条中MMA/As(Ⅴ)和As(Ⅲ)/As(Ⅴ)含量比值都低于0.5,说明As(Ⅴ)占主导地位,这可能是因为三七生长土壤氧化反应状态较好,不具备As还原反应的条件。主根部位由于As(Ⅴ)含量有所减少,因此MMA/As(Ⅴ)和As(Ⅲ)/As(Ⅴ)比值略有上升;剪口部位MMA含量超过As(Ⅴ),说明剪口部位的甲基化作用较根条和主根强;茎部MMA和As(Ⅲ)的含量都高于As(Ⅴ),且MMA含量占据主导位置,As(Ⅲ)次之。叶片中以As(Ⅲ)含量为主,MMA次之,因此MMA/As(Ⅲ)比值骤降为0.6,说明叶片的还原作用要强于其他各部位。从三七植株各部位各形态As含量分析来看,茎、剪口和根条中MMA含量差别不大,茎中MMA占总As比例高达61%,在剪口中的比例也达30%;叶片中As(Ⅲ)含量最高,占总As的45%。
综上所述,三七植株中As主要存在形态为As(Ⅲ)、As(Ⅴ)和MMA,且不同部位其形态含量变化也存在较大差异,说明在三七植株体内同时存在As的还原和甲基化作用。然而,三七植株的茎部MMA含量远高于As(Ⅴ)和As(Ⅲ),这一现象是否能说明茎部的还原作用强于其他部位,还需要进行蛋白鉴定和基因分析试验加以证实。
2.3 As在土壤和三七各部位间的吸收运转及形态转化
自然环境下,As在土壤-植物系统中的迁移和吸收转运受到土壤-植物屏障的有效控制[32]。根部吸收是As进入三七的主要途径,随后在其体内进行转运和再分配,土壤中As的生物有效性一定程度上决定了三七的受害程度和生物毒性。从三七植株生长情况来看,根条、剪口和主根属三七的地下部分,也是三七的主要药用部位,三七地下部分As含量大于地上部分,也在一定程度上说明三七As污染的主要来源是土壤。主根/土壤、根条/土壤、剪口/土壤、茎/剪口、叶/茎总As含量比值可在一定程度上揭示As从土壤到三七植株以及植株各部位间的吸收转运过程[20]。三七植株不同部位As(Ⅲ)、As(Ⅴ)和MMA含量比值变化分析见表2。
表2 三七植株各部位As的转运系数
Table 2 As translocation coefficient relative to tissue ofPanaxnotoginseng

As形态主根/土壤根条/土壤剪口/土壤主根/根条剪口/主根茎/剪口叶/茎总As0.0290.0840.0360.3211.3900.5591.952As(Ⅴ)0.0160.02410.6200.2881.4550.1244.320As(Ⅲ)0.4301.2600.6490.3850.7340.8173.637DMA0.0000.0000.0000.0000.0000.0000.000MMA0.0000.0000.0000.3067.9740.6293.485
从As转运系数的变化情况来看,主根、根条和剪口3个部位与土壤总As含量的比值均小于1,说明三七地下部分对土壤中As的吸收能力不强。与剪口、主根2个地下部位相比,根条/土壤总As含量比值较高,说明三七的根条较剪口和主根更易吸收土壤中的As。剪口和茎是连接三七地下部分和地上部分的2个主要部位,茎/剪口各形态As含量比值均小于1,说明As不容易从剪口转运到茎,可能剪口和茎相连的节存在某种阻止As转运的障碍因素。剪口/主根和叶/茎总As含量比值大于1,说明As在剪口、主根之间和在叶、茎之间的氧化作用较强,除了要排除三七种植过程中因大量喷施含As农药而导致叶片As积累量增加外,还要考虑这2个部位间是否存在某种特殊的影响As吸收转运机制的因素存在。叶/茎As(Ⅲ)、As(Ⅴ)含量比值远大于1,剪口/主根和叶/茎MMA含量比值也远大于1。
剪口是连接三七主根和地上茎之间的关键部位,起着运输和贮存养分、支撑茎叶的作用,是三七有效成分的主要贮存场所。因此,分析As从三七主根到剪口的迁移转运、各形态As的氧化还原和甲基化对三七中As毒性降解有重要意义。分别对土壤和三七主根、剪口的总As和各形态As含量进行Pearson相关性分析。结果(表3)表明,剪口中总As、As(Ⅲ)、As(Ⅴ) 含量分别与主根中对应形态As含量呈显著相关,说明As容易从主根转运迁移到剪口中。
表3 土壤和三七植株主根、剪口中总As和无机As的Pearson相关系数
Table 3 Pearson correlation of total As and inorganic arsenics in soil with their corresponding ones in taproot and rhizome ofPanaxnotoginseng
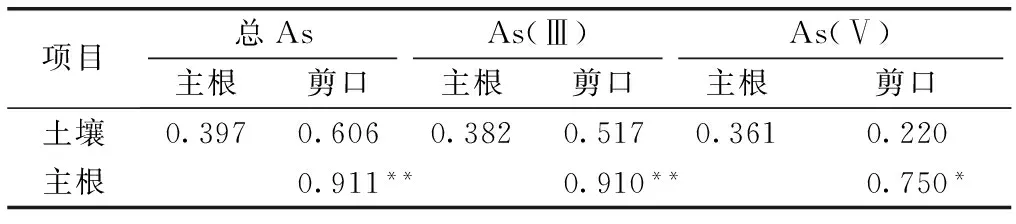
项目总AsAs(Ⅲ)As(Ⅴ)主根剪口主根剪口主根剪口土壤0.3970.6060.3820.5170.3610.220主根0.911**0.910**0.750*
*表示在 0.05 水平(双侧)显著相关;**表示在 0.01 水平(双侧)显著相关。
各形态As在植株中的迁移速率取决于植物品种,且不同研究得到的结论也不尽相同。LIU等[33]研究发现,无机As比螯合态As化合物更容易迁移转运;而LI等[34]研究发现,MMA和DMA(尤其是DMA)比无机As的迁移速度快很多。笔者研究中,从土壤到三七剪口As(Ⅴ)的迁移速率最快(转运系数为10.620),从主根到剪口MMA迁移速率最快(转运系数为7.974),As(Ⅲ)从土壤到三七根条的迁移速率较快(转运系数为1.260)。土壤中总As和各形态As含量与三七主根、剪口中As含量相关性未达显著水平,由此可以得出,并不是土壤中As含量超标,三七中As含量就会超标,只能说明不同种植土壤中As含量不同,通过As在土壤和三七间的转运迁移,不同地块三七中As含量也会有差别。
3 结论
采用HPLC-HG-AFS测定三七植株及其种植土壤中总As含量及各形态As含量,结果表明,三七种植土壤中As的主要赋存形态为无机态的As(Ⅴ)和As(Ⅲ),且As(Ⅴ)占主导地位。三七植株中As的主要存在形态为As(Ⅲ)、As(Ⅴ)和MMA,且不同部位其形态含量变化存在较大差异,说明在三七植株体内同时存在着各形态As间的转化和迁移转运作用。根部吸收是土壤As进入三七的主要途径,随着As在三七植物体内的转运,部分As(Ⅴ)转化为As(Ⅲ)和MMA形态存在,且不同植株部位间As存在形态差异,改变土壤中各形态As的生物有效性,在一定程度上可以有效缓解土壤As对三七的生物毒害。
[1] SMITH A H,GOYCOLEA M,HAQUE R,etal.Marked Increase in Bladder and Lung Cancer Mortality in a Region of Northern Chile Due to Arsenic Indrinking Water[J].American Journal of Epidemiology,1998,147 (7):660-669.
[2] YUAN Chun-gang,JIANG Gui-bin,HE Bin.Evaluation of the Extraction Methods for Arsenic Speciation in Rice Straw,OryzasativaL.,and Analysis by HPLC-HG-AFS[J].Journal of Analytical Atomic Spectrometry,2005,75(20):103-110.
[3] ROSEN B P.Biochemistry of Arsenic Detoxification[J].FEBS Letters,2002,529(1):86-92.
[4] 段桂兰,王利红,陈玉,等.水稻砷污染健康风险与砷代谢机制的研究[J].农业环境科学学报,2007,26(2):430-435.
[5] ELLIS D R,GUMAELIUS L,INDRIOLO E,etal.A Novel Arsenate Reductase From the Arsenic Hyper Accumulating FernPterisvittata[J].Plant Physiology,2006,141(4):1544-1554.
[6] TONG J T,GUO H M,WEI C.Arsenic Contamination of the Soil-Wheat System Irrigated With High Arsenic Groundwater in the Hetao Basin,Inner Mongolia[J].Science of the Total Environment,2014,496:479-487.
[7] YAN X L,LIN L Y,LIAO X Y,etal.Arsenic Accumulation and Resistance Mechanism inPanaxnotoginseng,a Traditional Rare Medicinal Herb[J].Chemosphere,2012,87(1):31-36.
[8] MA J,MI Y H,LI Q W,etal.Reduction,Methylation,and Translocation of Arsenic inPanaxnotoginsengGrown Under Field Conditions in Arsenic-Contaminted Soils[J].Science of the Total Environment,2016,550(1):893-899.
[9] ZHAO R,ZHAO M,WANG H,etal.Arsenic Speciation in Moso Bamboo Shoot:A Terrestrial Plant That Contains Organoarsenic Species[J].Science of the Total Environment,2006,371(1/2/3):293-303.
[10]LIU W J,ZHU Y G,HU Y,etal.Arsenic Sequestration in Iron Plaque,Its Accumulation and Speciation in Mature Rice Plants (OryzasativaL.)[J].Environment Science and Technology,2006,40(18):5730-5736.
[11]DHANKHER O P,ROSEN B P,MCKINNEY E C,etal.Hyperaccumulation of Arsenic in the Shoots of Arabidopsis Silenced for Arsenate Reductase (ACR2)[J].Proceedings of the National Academy of Sciences,2006,103(14):5413-5418 [12]BLEEKER P M,HAKVOORT H W J,BLIEK M,etal.Enhanced Arsenate Reduction by a CDC25-Like Tyrosine Phosphatase Explains Increased Phytochelatin Accumulation in Arsenate-TolerantHolcuslanatus[J].Plant Journal,2006,45(6):917-929.
[13]DUAN G L,ZHOU Y,TONG Y P,etal.A CDC25 Homologue From Rice Functions as an Arsenate Reductase[J].New Phytologist,2007,174(2):311-321.
[14]DUAN G L,ZHU Y G,TONG Y P,etal.Characterization of Arsenate Reducase in the Extract of Roots and Fronds of Chinese Brake Fern,an Arsenic Hyperaccumulator[J].Plant Physiology,2005,138(1):461-469.
[15]PICKERING I J,PRINCE R C,GEORGE M J,etal.Reduction and Coordination of Arsenic in Indian Mustard[J].Plant Physiology,2000,122(4):171-178.
[16]黄泽春,陈同斌,雷梅,等.砷超富集植物中砷化学形态及其转化的EXAFS研究[J].中国科学(C辑),2003,33(6):488-494.
[17]SCHMOGER M E V,OVEN M,GRILL E.Detoxification of Arsenic by Phytochelatins in Plants[J].Plant Physiology,2000,122(3):793-802.
[18]李卫东.文三州三七GAP种植区环境质量状况调查[J].云南环境科学,2004,23(增刊2):168-170.
[19]张文斌,曾鸿超,冯光泉等.不同栽培地区的三七总砷及无机砷含量分析[J].中成药,2011,33(2):291-293.
[20]阎秀兰,廖晓勇,于冰冰,等.药用植物三七对土壤中砷的累积特征及其健康风险[J].环境科学,2011,32(3):880-885.
[21]陈璐,米艳华,林昕,等.土壤-三七系统重金属污染调查及相关分析[J].中国中药杂志,2014,39(14):2608-2613.
[22]祖艳群,孙晶晶,郭先华,等.文山三七(Panaxnotoginseng)种植区土壤As空间分布特征及理化性质对三七As含量的影响[J].生态环境学报,2014,23(6):1034-1041 [23]Health Risk United States Environmental Protection Agency (USEPA).Method 3050B:Acid Digestion of Sediments,Sludges and Soils (Revision 2)[S].USEPA,1992.
[24]崔秀明,雷绍武.三七GAP栽培技术[M].昆明:云南科技出版社,2002:68.
[25]SADIQ M.Arsenic Chemistry in Soils:An Overview of Thermodynamic Predictions and Field Observations[J].Water Soil-Air Pollution,1997,93(1):117-136.
[26]ANDREAE M O.Arsenic Speciation in Seawater and Interstitial Waters:The Influence of Biological-Chemical Interactions on the Chemistry of a Trace Element[J].Limnology and Oceanography,1979,24(3):440-452.
[27]PETERSON M L,CARPENTE R.Biogeochemical Processes Affecting Total Arsenic and Arsenic Species Distributions in an Intermittently Anoxic Fjord[J].Marine Chemistry,1983,12(4):295-321.
[28]PETTINE M,CAMUSSO M,MARTINOTTI W.Dissolved and Particulate Transport of Arsenic and Chromium in the Po River,Italy[J].Science of the Total Environment,1992,119(92):235-280.
[29]WEI C Y,GE Z F,CHU WS,etal.Speciation of Antimony and Arsenic in the Soils and Plants in an Old Antimony Mine[J].Environmental and Experimental Botany,2015,109:31-39.
[30]彭昌军,姜秀丽,计红芳.铁锰复合氧化物对As(Ⅲ)、As(Ⅴ)的吸附研究及其在沼液中的应用[J].化工学报,2014,65(5):1848-1855.
[31]唐艳梅,黎其万,刘宏程.离子交换树脂-原子荧光光谱法测定三七中的4种形态砷[J].中成药,2012,34(3):513-517.
[32]LISE D,BERRY W L,KAPLAN I R,etal.Mineral Exploration:Biogeological Systems and Organic matter[J].Applied Geochemistry,1986,1(5):627-627.
[33]LIU W,WOOD B,RAAB A,etal.Complexation of Arsenite With Phytochelations Reduces Arsenite Effux and Translocation From Roots to Shoots in Arabidopsis[J].Plant Physiology,2010,152(4):2211-2221.
[34]LI R,STROUD J L,MCGRATH S P,etal.Mitigation of Arsenic Accumulation in Rice With Water Management and Silicon Fertilization[J].Environment Science and Technology,2009,43(10):3778-3783.
(责任编辑: 许 素)
Absorption,Translocation and Transformation of Arsenic in Medicinal Herb Panax notoginseng-Soil System.
DU Li-juan1, MI Yan-hua1, CHEN Lu1,YIN Ben-lin1, MA Jie1,2,HE Li-zhong1, YE Yan-ping1
(1.Institute of Agricultural Quality Standard & Testing Technique, Yunnan Academy of Agricultural Sciences, Kunming 650223, China;2.Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101,China)
In the present study, arsenic in soil andPanaxnotoginsengwas analyzed with the high performance liquid chromatography-atomic fluorescence (HPLC-HG-AFS), for total and species arsenic in an attempt to determine how the plant adsorbs, translocates and transforms arsenic. Results show that inorganic arsenate [As(Ⅴ)] and arsenite [As(Ⅲ)] were the main forms inP.notoginsenggrowing soil, and As (Ⅴ) was much higher than As (Ⅲ) in content, accounts for about 95% of the total. Arsenic distributed unevenly in the plant and the tissues of a plant displayed an order of branch root > rhizome > taproot > leaves > stems in terms of content of total arsenic, showing that roots is the main pathway of arsenic flowing from soil toP.notoginsengplant. The distribution of arsenic also varied with species [As (Ⅴ), As (Ⅲ) and MMA] and plant tissue. For As (Ⅲ), an order of leaves > branch root > rhizome > stems > taproot; for As (Ⅴ), an order of rhizome > branch root > taproot > leaves > stems and for MMA an order of root > stems > rhizome > leaves > taproot was found. Therefore, to reduce bioavailability of the arsenic in soil and to study in depth how to regulate transformation of arsenic between species [As (Ⅴ), As (Ⅲ) and MMA] in different parts ofP.notoginsengare the important ways to mitigate the hazard on safety caused by arsenic inP.notoginseng.
soil;Panaxnotoginseng; arsenic species; translocation
2016-04-01
国家自然科学基金(21267024);云南省科技计划青年项目(2014FD063);云南省科技创新人才培养计划(2014HB059,2015HC025)
X56
A
1673-4831(2016)06-0997-06
10.11934/j.issn.1673-4831.2016.06.020
杜丽娟(1981—),女,山西晋中人,助理研究员,硕士,主要从事农产品质量安全研究。E-mail: 36violet@163.com
① 通信作者E-mail: zhoumiqu@163.com
猜你喜欢
广西植物(2022年5期)2022-06-18
文山学院学报(2022年2期)2022-05-27
课外生活·趣知识(2022年5期)2022-05-14
池州学院学报(2017年3期)2017-10-16
河北果树(2016年2期)2016-08-12
西北园艺(果树)(2016年1期)2016-02-19
中国民族民间医药·下半月(2014年4期)2014-09-26
中国药业(2014年12期)2014-06-06
乡村科技(2014年21期)2014-03-04
