
喀斯特地区植物演替过程的多样性
2016-12-07 07:03:31熊康宁
浙江农业科学 2016年5期
檀 迪,熊康宁
(贵州师范大学中国南方喀斯特研究院 贵州省喀斯特山地生态环境国家重点实验室培育基地,贵州 贵阳 550001)
喀斯特地区植物演替过程的多样性
檀 迪,熊康宁*
(贵州师范大学中国南方喀斯特研究院 贵州省喀斯特山地生态环境国家重点实验室培育基地,贵州 贵阳 550001)
贵州毕节、花江地区喀斯特石漠化地区植被演替阶段分为草丛阶段、草灌阶段、藤刺灌丛阶段、乔灌林阶段和顶级群落阶段。不同的石漠化程度对应着群落演替的不同阶段。本文采取空间代替时间的方法研究不同演替阶段草本层、灌木层、乔木层和群落的物种丰富度、优势度Simpson指数、多样性Shannon指数和均匀度Pielou指数研究。结果表明,草本和灌木层植物多样性呈波浪式上升趋势;乔木层植物多为次生,分布稀少,变化趋势比较简单,呈直线上升趋势;群落的多样性与乔木层类似;喀斯特地区植被多样性并不是随演替阶段不断增长,而是在乔灌林阶段最大,顶级群落阶段有轻微下降;草本层在植物演替过程中表现出较好态势,在石漠化治理过程中要注重对草木的保护和利用;狗尾草、金丝桃、铁仔、白栎等植物分别作为强度、中度、轻度和潜在石漠化样地的优势种,在石漠化治理过程中发挥着重大作用,应在石漠化治理不同阶段予以重视。
石漠化;物种多样性;乔木层;优势种植被;演替
植被是一个地区生态环境的综合反映,其演替和分布格局会受到生态过程、植被演化历史等因素的影响[1]。在自然及人为作用的控制下,植被之间由于长期的相互作用导致了不同层次的时空变异,形成有规律的植被分布格局,这种分布格局既能反映植被对资源的利用及对环境的适应,也反映植被内部的更新与竞争关系。植被在时间和空间上的分布是典型的分维数体[2],因而其空间分布格局也因尺度依赖性变得更加复杂[3]。
中国西南是世界上湿润-半湿润、热带-亚热带中喀斯特地貌发育最大的地区[4]。我国喀斯特分布面积超约全国总面积的13%,其中以贵州为中心的桂北、滇东及湘西等地的喀斯特面积已超过55万km2。在南方喀斯特山地典型脆弱区中,贵州省所占面积高达73.8%。从生态环境的角度看,贵州是中国的 “喀斯特省”[5]。
中国西南喀斯特顶极群落为亚热带喀斯特常绿落叶阔叶混交林,由于受地质运动、气候等因素的影响,此地区适生植物表现出明显的嗜钙性、耐旱性和石生性等特点,生态系统的稳定性和抗干扰性差,与黄土、沙漠、寒漠并列为中国四大生态环境脆弱区。一般认为,喀斯特地区植被恢复须经历草丛阶段→草灌阶段→藤刺灌丛阶段→乔灌林阶段→顶级群落阶段的演变过程[6],但人地矛盾导致许多地方表现出生态环境退化的“石漠化”现象,是我国植被恢复和生态重建的重点和难点[7-8]。
进入21世纪以来,随着喀斯特的危害和成因进一步明确,对地区植被自然恢复过程群落演替规律也有了一定的研究[9-11]。由于自然条件和治理模式的差异,因而不同地区选取的植物也存在差异,同时对不同地貌单元、不同植被演替阶段与不同石漠化程度双重影响下物种多样性变化研究较少[12]。本研究以贵州喀斯特高原山地、高原峡谷石漠化环境为研究对象,通过野外实地监测,对不同程度石漠化、不同群落演替阶段植物多样性响应机制进行研究,以期为喀斯特地区生态系统恢复重建提供理论支持。
1 材料与方法
1.1 研究区概况
毕节撒拉溪示范区地处滇东高原向黔中高原过渡的东斜坡地带,地层出露较齐全,地质构造复杂,最高海拔1742.3m,最低海拔1400m,相对高差为342.3m,地貌类型为典型的喀斯特高原山地。该区属亚热带湿润季风气候区,流域内年平均气温14.03℃,年均降雨量863mm,主要集中在7—9月,占全年总降雨的52.4%。岩石以石灰岩及紫色砂页岩为主。受成土母质影响,一般分布黄壤土及紫砂土。野生植被是以大白杜鹃、小果十大功劳、来江藤、金丝桃等为主的藤、刺、灌丛,以及零星分布的青冈、核桃、通脱木为主。
关岭-贞丰花江示范区位于北盘江,海拔500~1200m,相对高差700m,为典型的喀斯特高原峡谷。该地区属亚热带湿热河谷气候,冬春温暖干旱,夏秋湿热,热量资源丰富,年均温 18.4℃,年均极端最高气温 32.4℃,年均极端最低气温6.6℃。年均降水量1100mm。年降雨量时空分布不均,5—10月降雨量占全年总降雨量的83%。平地占2%,山地占98%,耕地资源短缺,地下水埋深300m以下,喀斯特干旱现象严重。土壤以石灰土为主,其结构不良,质地黏重,质量差。植被为亚热带常绿落叶针阔混交林,原生植被基本上被破坏,现以次生林为主。野生植被是以刺梨、花椒、忍冬等为主的藤、刺、灌丛,以及零星分布的构树、臭椿、油桐为主。
1.2 样地设置
研究根据贵州喀斯特石漠化地区土地植被恢复自然演替的特征,在对研究区详细调查的基础上,根据代表性和典型性原则,结合石漠化演替5个典型阶段 (无石漠化、潜在石漠化、轻度石漠化、中度石漠化、强度石漠化)为对象,设置2个20m×20m的样方。用插值法将样方详细分为4个10m×10m的小型样方和16个5m×5m的微型样方,以微型样方为基本单位,调查每个胸径(DBM≥1cm)的个体;抽取8个小样方对灌木层进行调查;按梅花形随机抽取10个微型样方对草本层进行调查。调查内容包括乔、灌木的种类、数量、胸径 (乔木)、基径 (灌木)、高度、冠幅;草本植物种类、高度、盖度。同时调查记录地理位置、海拔高度、土壤类型、坡度坡向等环境因子,各样地基本情况见表1。

表1 研究区日的基础信息
1.3 植被分析方法
通过对试验区进行常规生态监测,将森林群落分为乔木层、灌木层和草本层;灌丛群落分灌木层与草本层;草丛群落中因有一定盖度的灌木也分为灌木层与草本层。分别计算各群落乔木、灌木、草本层的重要值。乔木植物重要值=相对密度+相对优势度+相对频度;灌木和草本植物重要值=相对高度+相对盖度。同时,采用α多样性测度来测量所查区域内森林群落的物种多样性。
丰富度 (R)=S;
多样性Shannon-Wiener指数(H)=-∑(PilnPi);
均匀度Pielou指数(E)=H/lnS;
优势度Simpson指数(D)=1-∑[Ni(Ni-1)/N(N-1)]。
式中,S为群落中的总种数;Pi为种i的相对重要值,即Pi=ni/N,ni为中i的个体数,N为观察到的个体数总数。
2 结果与分析
2.1 不同等级石漠化环境植被物种组成及植物多样性
2.1.1 植物组成
由表2—3可以看出,喀斯特地区植被组成简单,随着石漠化程度的加深,植被类型相应减少。在毕节试验区,有草本植物15科15属17种,木本植物14科16属17种。而在花江试验区,草本植物只有7科11属12种,木本植物仅11科3属13种,多数情况下植被呈现出1科1属1种的形态。研究发现,禾本科、毛茛科、菊科表现出多属多种形态,说明以上植物对喀斯特环境具有较好的适应性,在生态治理中可适当应用。而像井栏边草、蜈蚣草、构树等植物在不同等级石漠化地区的重要值呈显著差异,说明同一种植物在不同石漠化地区的适应程度也不同。重要值研究可为研究石漠化地区植物的优势种和建群种提供依据。
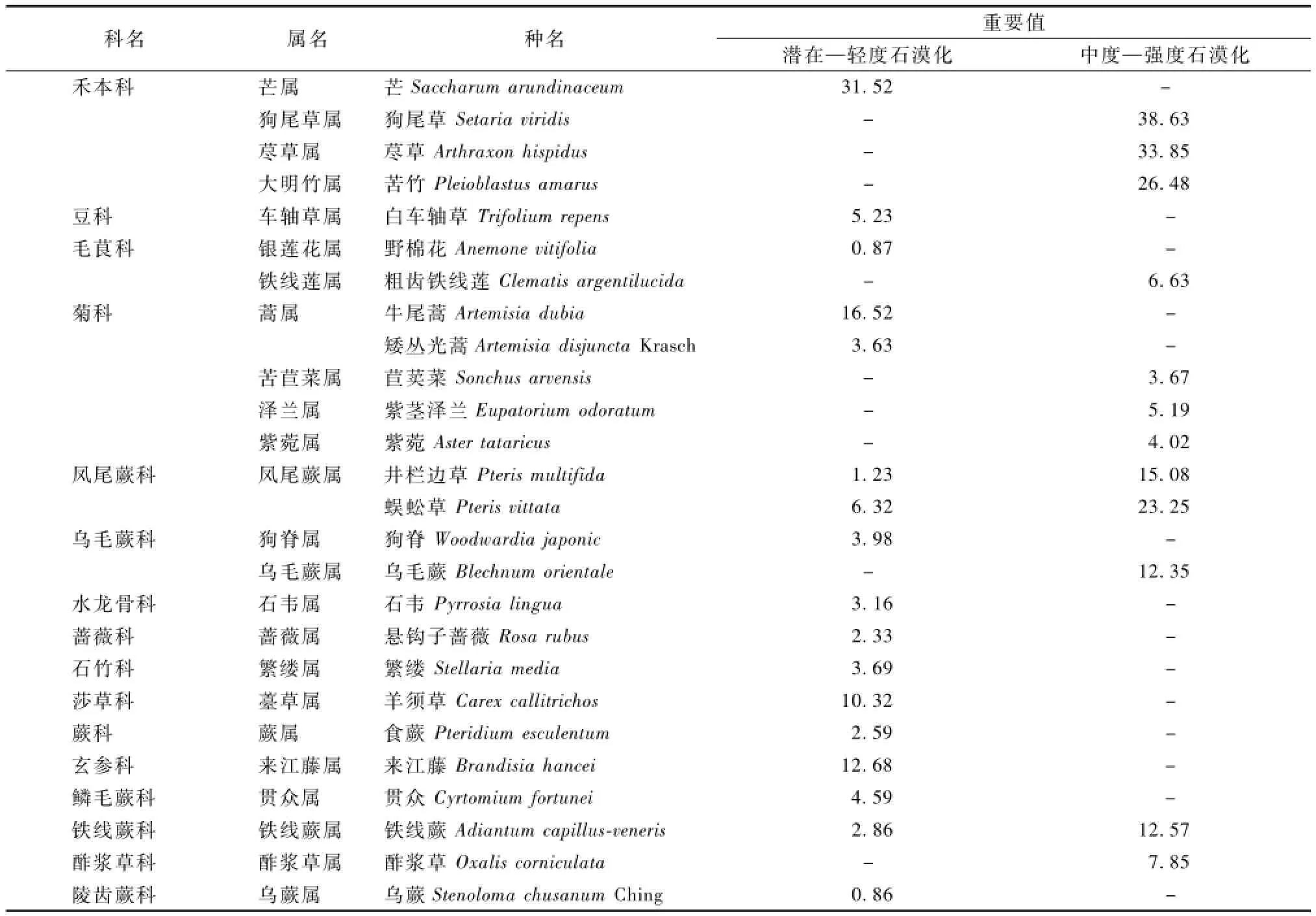
表2 草本层石漠化环境植被群落的物种情况
在潜在—轻度石漠化地区,芒、牛尾蒿、女贞等是本地区植物群落的建群种或优势种,而在中度—强度石漠化地区狗尾草、荩草、苦竹等则是本地区植物群落的建群种或优势种。
2.1.2 不同等级石漠化地区植物群落物种多样性分析
实地调查发现,研究中5个石漠化演替等级(无石漠化、潜在石漠化、轻度石漠化、中度石漠化、强度石漠化)较好地与喀斯特地区植被恢复须经历阶段 (草丛阶段→草灌阶段→藤刺灌丛阶段→乔灌林阶段→顶级群落阶段)吻合,遂分别测度乔木、灌木、草本植物的多样性指数。
由图1—2可以看出,草本层物种各项多样性测度指数变化规律大体一致,即在由草丛阶段发展到草灌阶段的过程中,各项指数均呈下降趋势,在草灌阶段达到最小值,然后逐渐升高,并最终达到稳定状态。其原因是在草丛阶段喀斯特地区生境恶劣,仅适合部分一年生草木的生长。草灌阶段由于植被返还土壤的有机质极少,一年生草本逐渐退出群落,金丝桃作为此阶段的建群种,在群落中的作用突出,降低了群落的均匀度,致使物种多样性差。同时灌木的生长增加了环境的郁闭度,使得部分阳性草种死亡。随着演替阶段的进行,灌木层、乔木层的加入,增加了喀斯特环境的复杂性,物种分布更均匀,物种多样性随之上升,并最终达到相对稳定。
灌木层物种多样性指数除丰富度外,另3个指数基本呈现出相同的变化趋势 (图1,3)。灌木层丰富度在乔灌林阶段达到峰值,之后随演替进行逐渐下降并达到稳定。而Simpson指数、Shannon指数和均匀度在演替过程中总体表现为波动上升的趋势。3指数在草灌阶段出现第1个峰值,然后下降到藤刺灌丛阶段出现低值,之后上升到乔灌林阶段出现第2个高峰值,然后下降最终达到稳定状态。原因是在由草丛阶段向灌丛阶段发展的过程中,草木逐渐被灌木所替代,灌木物种增多,多样性达到峰值。直至藤刺灌丛阶段,乔木层的出现缩小了灌木的生长空间,使其多样性在藤刺灌丛阶段急剧下滑;在乔灌林阶段,中性和耐荫的物种得以发展,灌木多样性上升并达到最大值;在顶级群落阶段,由于乔木群落中的作用更加突出,阳性物种基本退出群落,灌木多样性物种在此阶段有轻微下降并最终达到稳定。
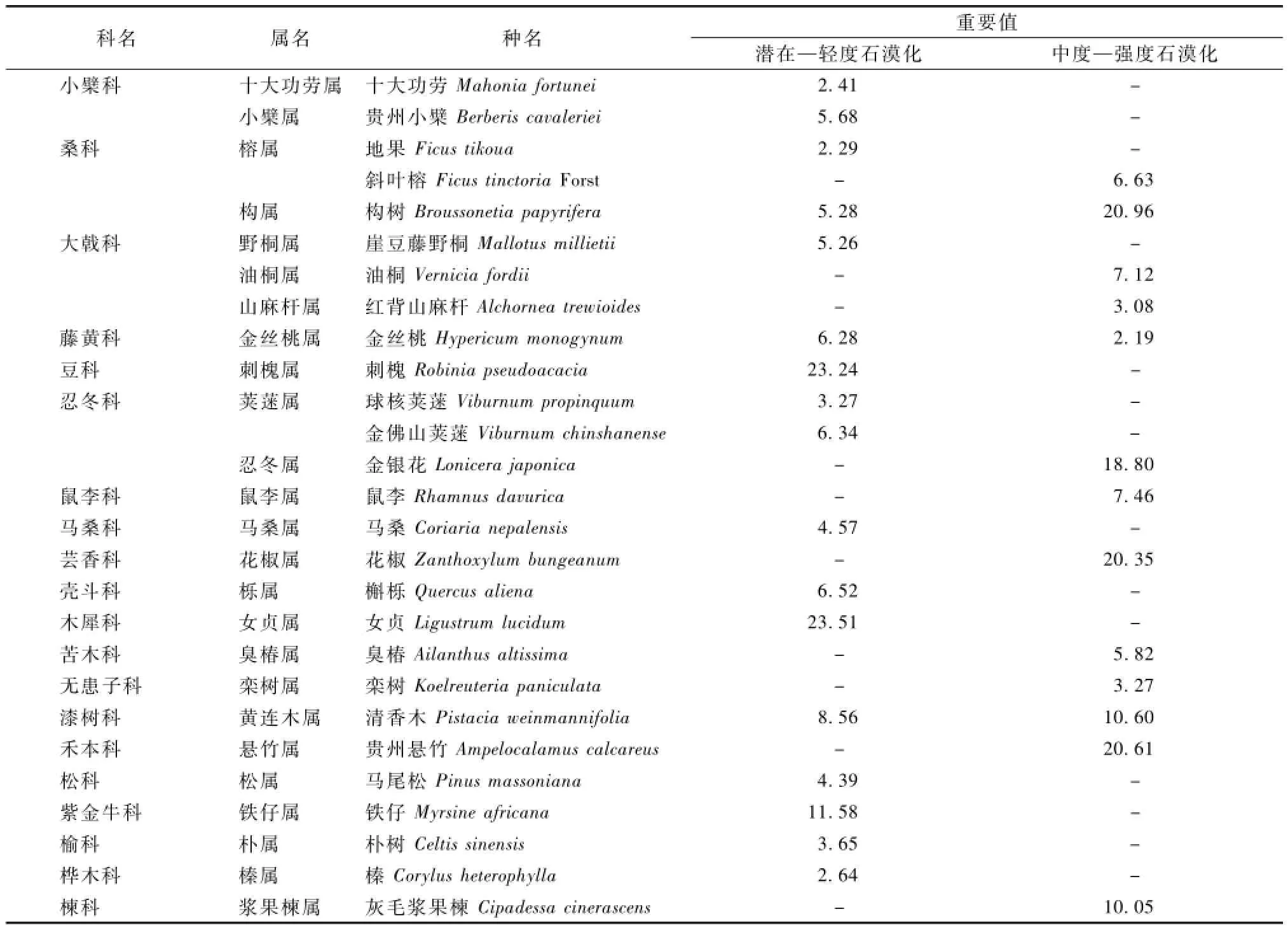
表3 木本层石漠化环境植被群落的物种情况
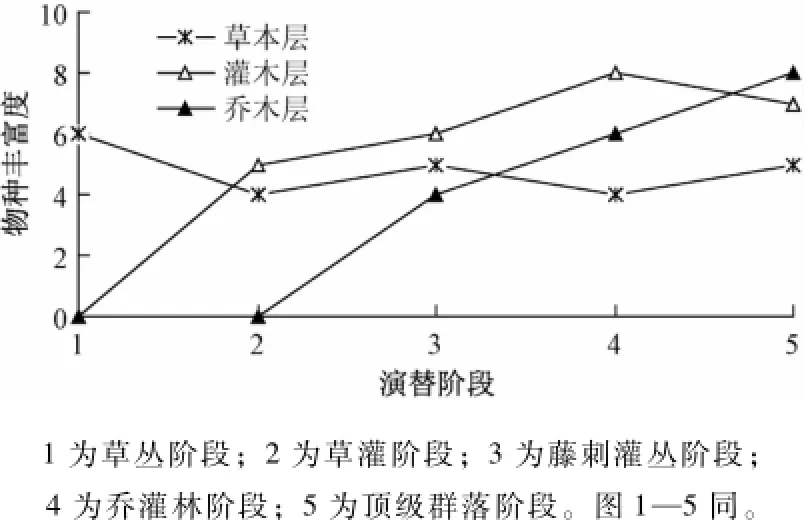
图1 不同演替阶段不同群落的物种丰富度

图2 草本层物种的多样性分析
由图4可知,在整个植被演替过程中,乔木丰富度逐渐升高到顶级群落阶段有部分的回落。这是由于乔木先锋树种的出现,使林下生境发生改变,同时还能改善土壤条件。群落内的小气候达到中性树种的生长要求,这时既有进入衰退期的阳性植物,又有缓慢发展的中性植物,所以乔木多样性指数在乔灌林阶段达到顶峰;进入顶极群落阶段,由于大部分中性树种进入乔木中上层,削减了投射阳光的数量,阻碍了阳性树种的生长,阳性树种逐渐退出群落,故顶极群落阶段的乔木层物种多样性会有一定程度的降低。

图3 灌木层物种的多样性分析
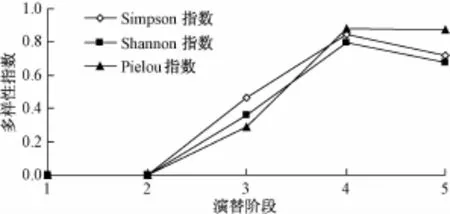
图4 乔木层物种的多样性分析
从图5可以看出,物种多样性并非随着演替阶段的发展不断增大,而是从草丛阶段开始增加,至乔灌林阶段达到峰值,在演替到顶极群落阶段时物种多样性会有一定程度的下降。这是因为在顶级群落之前的演替阶段中,随着植物对环境的改良作用,物种多样性总体得到提高。而达到顶极群落阶段之后,由于群落的稳定性增大,各个层位的生态位逐渐饱和,新物种浸入比较困难,同时一些物种由于在竞争中处于不利地位,逐渐退出群落,这使得物种多样性会有一定程度的降低。但这种降低不代表群落的衰退,相反却是群落的成熟和稳定的表现。
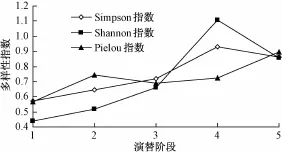
图5 群落的多样性分析
2.1.3 不同演替阶段物种多样性指数的相关性分析
为了验证多样性指数的有效性,对喀斯特地区不同演替阶段的物种多样性指数进行相关性分析。由表4可看出,喀斯特地区不同演替阶段物种多样性指数间的相关性差异较大,部分物种多样性指数间的相关性极强,达到显著或极显著水平。其中,R与D,D与H的相关性最大,相关系数分别为0.960和 0.984,且均达到极显著水平 (P<0.01),而R与H达到显著相关水平 (P<0.05),说明物种丰富度指数、Simpson指数和Shannon指数能较好地反映喀斯特地区植被不同演替阶段的物种多样性特征。
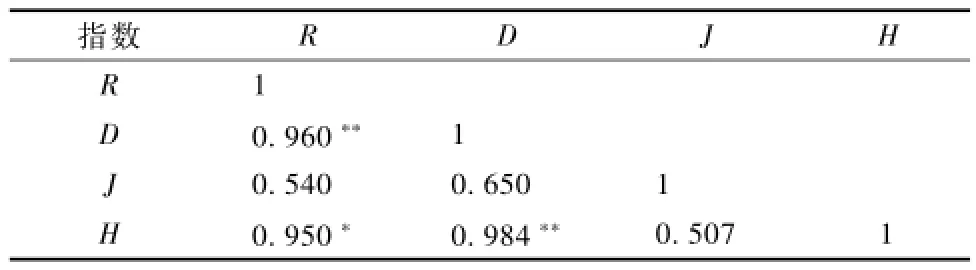
表4 物种多样性指数间的相关系数
3 小结
在植被恢复初期,一些根系发达的先锋植物如狗尾草、荩草出现,不断地改良生境的土壤水分状况,为更多物种的浸入奠定基础。到草灌群落阶段,物种的多样性降低。随着演替进程的发展及枯枝落叶的积累,土壤肥力得到提高,生境进一步改善。进入乔灌林阶段,群落内的物种种类和数量进一步增加。乔木层的出现重新分配了生境内的空间、光照等资源,从而形成上层、中层、底层的林相结构。顶极群落阶段系统处于稳定的动态平衡中,物种结构相对稳定。
群落演替过程与物种多样性密切相关。不同研究的结论不同。一种观点认为,随着群落演替阶段发展,群落的物种多样性越来越高,顶极群落的稳定性一般要高于演替中的群落[13-16]。另一种观点则认为,在演替后期多样性出现下降。Odum[17]认为,群落多样性指数并非随演替进展一直增加,在群落演替的初期和中期阶段,物种多样性与演替阶段呈正相关,但随后会出现下降趋势。此外,通过对缙云山常绿阔叶林次生演替序列群落物种多样性动态研究发现,在群落的发育过程中,物种多样性指数高并不能与群落稳定画等号,物种多样性指数在乔灌林阶段最高,但这个阶段并不是群落稳定性最高的阶段[18]。本文研究与第2种观点相吻合。
通过对草本层、灌木层和乔木层的对比发现,除了最终的顶级群落阶段乔木层的物种多样性略多于草本层之外,其余阶段草本层的多样性指数最大。这也说明草木在适应不同喀斯特环境中体现出较强的优势,因此在对喀斯特环境的治理中要加强对草本植物的保护和利用。
从贵州喀斯特地区植被自然演替过程可以看出,草丛阶段一年生的草本植物为优势种,对于喀斯特环境的改善起着不可小觑的作用,结合试验区植物的重要值,对于强度石漠化地区应加强对狗尾草、荩草的利用。在草灌阶段,灌木加入,对环境的改良作用更加显著。在此阶段,先锋种金丝桃、优势种平枝栒子、火棘和马桑都占有很大比重,因此对于中度石漠化地区应加强对上述植物的利用。在藤刺灌丛阶段,尽管存在少量乔木 (如马尾松),但中性灌木铁仔和金佛山荚蒾则是次阶段的优势种,因此对于轻度石漠化地区应加强对铁仔和荚蒾的利用。在乔灌林阶段,乔木层在环境改良中则占有更重要的位置。白栎、青冈、通脱木等成为此阶段优势种,在潜在石漠化治理过程中可以加强对上述植物的利用。
[1] QIANH,RICKLEFSRE.Acomparisonofthetaxonomic richnessofvascularplantsinChinaandtheUnitedStates[J]. TheAmericanNaturalist,1999,154(2):160-181.
[2] 柳锦宝,姚云军,张永福.基于分维的多尺度植被空间格局分析 [J].水土保持研究,2007,14(1):155-158.
[3] 杜华强,汤孟平,周国模,等.天目山物种多样性尺度依赖及其与空间格局关系的多重分形 [J].生态学报,2007,27(12):5038-5049.
[4] 朱华.中国南方石灰岩 (喀斯特)生态系统及生物多样性特征 [J].热带林业,2007,35(S1):44-47.
[5] 熊康宁,黎平,周忠发,等.喀斯特石漠化的遥感:GIS典型研究 [M].北京,地质出版社,2002.
[6] 曾馥平,彭晚霞,宋同清,等.桂西北喀斯特人为干扰区植被自然恢复22年后群落特征 [J].生态学报,2007,27(12):5110-5119.
[7] 袁道先.现代岩溶学在我国的发展 [J].中国科学基金,2005,19(3):139-141.
[8] 贺庆棠,陆佩玲.中国岩溶山地石漠化问题与对策研究[J].北京林业大学学报,2006,28(1):117-120.
[9] 喻理飞,叶镜中.退化喀斯特森林自然恢复评价研究[J].林业科学,2000,36(6):12-19.
[10] 区智,李先琨,吕仕洪,等.桂西南岩溶植被演替过程中的植物多样性 [J].广西科学,2003(1):63-67.
[11] 杨汉奎,程仕泽.贵州茂兰喀斯特森林群落生物量研究[J].生态学报,1991(4):307-312.
[12] 李晨,熊康宁,吴光梅,等.岩溶山区石漠化生态治理的植物多样性动态响应:以贵州省花江示范区顶坛小流域为例 [J].热带地理,2012,32(5):487-492.
[13] MARGALEFR.Informationtheoryinecology[J].General Syst,1957(3):37-71.
[14] 马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数 [J].生态学报,1995,15(3):268-277.
[15] 宋同清,彭晚霞,曾馥平,等.木论喀斯特峰丛洼地森林群落空间格局及环境解释 [J].植物生态学报,2010,34(3):298-308.
[16] 彭晚霞,宋同清,曾馥平,等.喀斯特常绿落叶阔叶混交林植物与土壤地形因子的耦合关系 [J].生态学报,2010,30(13):3472-3481.
[17] ODUMEP.Thestrategyofecosystemdevelopment[J]. Science,1969,164:262-270.
[18] 韩玉萍,李雪梅,刘玉成.缙云山常绿阔叶林次生演替序列群落物种多样性动态研究 [J].西南师范大学学报 (自然科学版),2000,25(1):62-68.
(责任编辑:张瑞麟)
S688
A
0528-9017(2016)05-0788-06
2016-01-31
国家科技支撑计划重大课题 (2011BAC09B01)
檀 迪 (1990—),男,河北廊坊人,在读硕士,研究方向植物生态学,E-mail:494976338@qq.com。
熊康宁,E-mail:584572163@qq.com。
文献著录格式:檀迪,熊康宁.喀斯特地区植物演替过程的多样性 [J].浙江农业科学,2016,57(5):788-793.
10.16178/j.issn.0528-9017.20160553
猜你喜欢
长江科学院院报(2023年8期)2023-08-29 07:05:22
建筑与预算(2022年6期)2022-07-08 13:59:18
内蒙古林业调查设计(2021年2期)2021-06-29 02:38:12
阅读(低年级)(2020年8期)2020-11-06 06:23:19
乡村地理(2019年2期)2019-11-16 08:49:32
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26 13:44:20
中国水土保持(2018年11期)2018-11-06 11:52:38
学术论坛(2016年5期)2016-05-17 05:44:46
文化月刊·下旬刊(2014年6期)2014-08-28 00:58:59
建筑与预算(2014年5期)2014-04-11 07:29:28
