
Counteraction of Nogo-A and axonal growth inhibitors by green tea polyphenols and other natural products
2016-12-02 06:04TiffanyK.Fan,UshaGundimeda,WilliamJ.Mack等
中国神经再生研究(英文版) 2016年4期
PERSPECTIVE
Counteraction of Nogo-A and axonal growth inhibitors by green tea polyphenols and other natural products
Neuronal injuries such as stroke, traumatic brain injury, and spinal cord injury are leading causes of major disability and death. Chronic therapy for these neuronal injuries requires the promotion of axonal regeneration from the remaining neurons (Schwab and Strittmatter, 2014). However, the local environment in the central nervous system (CNS) is unfavorable to this regeneration due to the presence of myelin-derived axonal growth inhibitors such as Nogo-A, myelin-associated glycoprotein (MAG), and oligodendrocyte myelin glycoprotein (OMgp). In addition, chondroitin sulfate proteoglycans (CSPGs) associated with astroglial scarring also inhibit axonal regeneration after neuronal injury. Several studies support the role of Nogo-A in limiting axonal regeneration. It has two distinct domains with antineuritogenic potential: N-terminal amino-Nogo and C-terminal Nogo-66, a 66 amino acid containing domain. Strittmatter and his associates identified a cell surface glycosyl phosphatidylinositol-anchored membrane protein, NgR1, as a receptor for Nogo-66 (Schwab and Strittmatter, 2014). Later studies have revealed that MAG, OMgp, and CSPGs also bind to NgR1 which may mediate their inhibiting activity on the axonal growth. NgR1 signals through interaction with p75 neurotrophin receptor (p75NTR) or a homologous protein, TROY, as well as a transmembrane protein, LINGO-1. The association between p75NTR and Rho-GDI leads to the release of RhoA and its activation. RhoA in turn activates its effector, Rho-associated protein kinase (ROCK). This ultimately leads to actin-cytoskeletal reorganization, which results in the collapse of growth cones and inhibition of neurite outgrowth (Figure 1).
Current approaches to overcome Nogo-A and need for natural products: Currently, there are no clinically proven synthetic neuroregenerative drugs for recovery from neuronal injuries. Various pharmacological agents acting through different mechanisms are currently being evaluated for blocking NgR1 or its downstream signaling to enhance axonal sprouting and functional recovery from stroke and spinal cord injury (Schwab and Strittmatter, 2014). Some examples include antibodies to block Nogo-A, Nogo-66 antagonistic peptides, NgR1 decoys, inhibitors to prevent RhoA and ROCK activities, and inosine to activate Mst3b protein kinase. Furthermore, agents that elevate intracellular cAMP are also being evaluated for inhibiting the antineuritogenic action of Nogo-A. An alternative and complementary approach to the development of synthetic drugs for stroke is to evaluate the efficacy of inexpensive and safe natural compounds and to elucidate their mechanisms of action to optimize their therapeutic effects.
Green tea polyphenols for the treatment of neuronal injuries: Green tea (Camelia sinesis), one of the most popular and widely consumed beverages in the world, may be well-suited for treating these neuronal injuries. Epidemiologic data suggest that daily consumption of green tea could prevent the onset of ischemic stroke (Kokubo et al., 2013). The polyphenols present in green tea have been shown to mediate its beneficial effects. Green tea polyphenols (GTPP) include both galloylated polyphenols epigallocatechin-3-gallate (EGCG) and (-)-epicatechin-3-gallate, as well as nongalloylated polyphenols, (-)-epigallocatechin and (-) epicatechin. EGCG, the major polyphenol present in GTPP, has neuroprotective, neurorescue, and neuroregenerative properties (Mandel et al., 2005). Both EGCG and unfractionated GTPP have been shown to decrease the extent of neuronal injury when administered during or immediately after ischemic brain injury in rodents (Hong et al., 2000). In addition, recent studies have presented evidence that GTPP and EGCG may enhance functional recovery after stroke and spinal cord injury in experimental models (Tian et al., 2013).
EGCG exhibits antioxidant activity in vitro, which may have some significance to neuroprotection. However, the effective antioxidant activity requires a concentration of EGCG two orders of magnitude higher than that reached in the plasma (<1 μM) following green tea consumption (Gundimeda et al., 2015). In contrast, only low micromolar or submicromolar concentrations of EGCG are required for induction of cell signaling significant for neuroprotection, neurorescue, and neuroregeneration (Mandel et al., 2005). EGCG may elicit some of its actions at low concentrations due to its high affinity binding to some cellular proteins. Tachibana and his associates have identified a high affinity binding of EGCG (Kd of 40 nM) to the 67-kDa laminin receptor (67LR), a nonintegrin-type cell surface-associated receptor (Tachibana et al., 2004). 67LR was originally discovered as a laminin-binding protein involved in cancer cell invasion and is currently being targeted for cancer prevention by EGCG. We have previously reported a neuroprotective role for 67LR in mediating EGCG-induced preconditioning of neuronal-like cells against cell death induced by oxygen-glucose deprivation (Gundimeda et al., 2012).
GTPP counteracts the antineuritogenic action of Nogo-A: Recently, we have shown that low concentrations of unfractioned GTPP and submicromolar concentrations of EGCG can prevent both the neurite outgrowth inhibiting activity and growth cone collapsing activity of Nogo-66 in the NGF-differentiated Neuroscreen-1 neuronal cells (Gundimeda et al., 2015). In addition, we found a synergistic interaction among GTPP constitutents. Studies reveal that the preventive action of EGCG is mediated through its binding to high-affinity cell-surface receptor 67LR. A series of experiments supported a possible second messenger role of H2O2in EGCG-mediated prevention of the antineuritogenic action of Nogo-66. Furthermore, a steady state generation of exogenous H2O2alone in micromolar concentrations mimicked EGCG action. However, the H2O2-mediated anti-Nogo-66 actions do not require 67LR suggesting H2O2may bypass 67LR and block the signaling pathway induced by Nogo-A. Among the proteins involved in the downstream signaling of Nogo-A, RhoA is one of the redox sensitive targets. Therefore, it is possible that H2O2generated in response to receptor-mediated actions of EGCG may inhibit RhoA through a redox sensitive mechanism and thereby block Nogo-A action (schematically depicted in Figure 1). Certainly, further studies are needed to understand the novel mechanism by which EGCG/GTPP can prevent the antineuritogenic action of Nogo-A.
Although both EGCG and GTPP are effective in preventing the antineuritogenic action of Nogo-A, they cannot block the antineuritogenic action of MAG in Neuroscreen-1 neuronal cells (Gundimeda et al., 2015). In this context, it is interesting to note that MAG also acts through binding to NgR2 and other receptors in addition to NgR1. Since CSPGs act, in part, by binding to NgR1 and activation of RhoA (Schwab and Strittmatter, 2014), it is interesting to determine whether EGCG may also inhibit their antineuritogenic action.
GTTP stimulation of adult neurogenesis - possible by a negative modulation of Nogo-A: Besides enhancing neuritogenesis, GTPP may also increase adult neurogenesis. Some studies have shown an increase in adult neurogenesis and survival of neural stem cells by EGCG (Wang et al., 2012). Given that Nogo-A/NgR1 inhibits neurogenesis (Rolando et al., 2012), GTPP may enhance neurogenesis by blocking the Nogo-A/ NgR1 signaling pathway. Furthermore, GTPP potentiates the neuritogenic action of brain-derived neurotrophic factor (BDNF) (Gundimeda et al., 2014). Besides enhancing neuritogenesis, BDNF also enhances neurogenesis. Moreover, BDNF has been shown to block Nogo-A action in vitro. Therefore, GTPP either directly or indirectly through neurotrophins may block Nogo-A/NgR1 signaling and thereby enhance both adult neurogenesis and axonal growth and can improve neuronal plasticity and functional recovery after neuronal injuries.
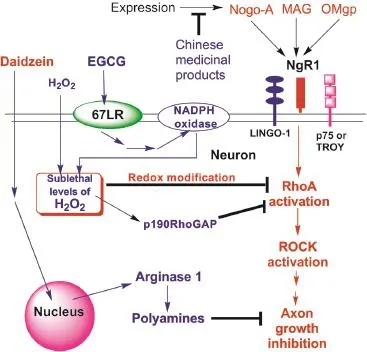
Figure 1 Schematic presentation of proposed mechanisms by which GTPP and other natural products counteract Nogo-A and other axonal growth inhibitors.
Other natural products counteracting Nogo-A and other axonal growth inhibitors: Besides GTPP, Nogo-A/NgR1 may be counteracted by a variety of natural products causing functional recovery after neuronal injuries. A recent review discussed intervention of Nogo-A action by a variety of Chinese medicines (Qin et al., 2012). The Fujian tablet (a mixture of extracts from buckwheat, barrenwort, and cassia seed) has been demonstrated to inhibit the expression of Nogo-A, thereby promoting neurogenesis (Liu et al., 2011). An administration of the Fujian tablet significantly decreased Nogo-A expression at various stages following cerebral infarction in rats (Qin et al., 2012). Another study witnessed the same effect and also demonstrated that lowering Nogo-A expression in the cervical spinal cord could greatly improve motor function following focal cerebral ischemia in rats (Qin et al., 2012). Similarly, panaxtrial saponins, found in ginseng roots, may also act to downregulate the expression of Nogo-A. Rats that were given panaxtrial saponins immediately after focal ischemia-reperfusion injury exhibited lower levels of Nogo-A-immunoreactive cells. Further, the Zuogui pill (which contains extracts from rehmania root, wolfberry, and yam) and Yougui pill (which additionally incorporates extracts from cinnamon) promote axonal regeneration by not only reducing the expression of Nogo-A and its receptor NgR1, but also by increasing nerve growth factor, a crucial factor in neuritogenesis (Qin et al., 2012).
Another natural product, daidzein, is a soy isoflavone that protects CNS neurons from the neurite outgrowth-inhibiting activity of MAG (Ma et al., 2010). In a novel study, Ma and associates screened a library of small molecular drugs for transcriptional induction of arginase 1, which has demonstrated protective effects on motor and sensory neurons during trophic factor deprivation and axonal growth inhibition respectively. Preconditioning hippocampal cells with daidzein before exposing them to MAG prevented its antineuritogenic action (Ma et al., 2010). Additionally, daidzein effectively promoted neuritogenesis in vivo and was able to cross the blood-brain barrier. Amphotericin B, a microbial product which is used as an antifungal drug, has been shown to inhibit not only myelin-derived axonal growth inhibitors Nogo-A and MAG, but also glial-scar-derived CSPGs (Gao et al., 2010). However, amphotericin B cannot effectively cross the blood-brain barrier. In addition, it is too toxic. Therefore, EGCG, daidzein and other natural products which have been shown to cross the blood-brain barrier and are safe may be useful for further development as effective neuroregenerative agents.
GTPP and other natural products discussed in this article may intervene the actions of Nogo-A and in some cases other axonal growth inhibitors such as MAG and CSPGs, thereby stimulating both axonal growth and neurogenesis. These neuroregenerative agents may be potential candidates for evaluating their efficacy in enhancing functional recovery after neuronal injuries in humans. Furthermore, identifying molecular targets for these natural products, such as 67LR in the case of EGCG, could open new avenues for developing novel therapeutic drugs for stroke and other neuronal injuries.
Tiffany K. Fan, Usha Gundimeda, William J. Mack, Rayudu Gopalakrishna*Department of Cell and Neurobiology (Fan TK , Gundimeda U, Gopalakrishna R) and Department of Neurological Surgery (Mack WJ), Keck School of Medicine, University of Southern California, Los Angeles, CA, USA
*Correspondence to: Rayudu Gopalakrishna, Ph.D., rgopalak@usc.edu.
Accepted: 2016-02-26
orcid: 0000-0002-0398-3166 (Rayudu Gopalakrishna)
Gao Y, Deng K, Cao Z, Graziani EI, Gilbert AM, Koehn FE, Wood A, Doherty P, Walsh FS (2010) Amphotericin B, identified from a natural product screen, antagonizes CNS inhibitors to promote axon growth via activation of an Akt pathway in neurons. J Neurochem 113:1331-1342.
Gundimeda U, McNeill TH, Elhiani AA, Schiffman JE, Hinton DR, Gopalakrishna R (2012) Green tea polyphenols precondition against cell death induced by oxygen-glucose deprivation via stimulation of laminin receptor, generation of reactive oxygen species, and activation of protein kinase Cepsilon. J Biol Chem 287:34694-34708.
Gundimeda U, McNeill TH, Barseghian BA, Tzeng WS, Rayudu DV, Cadenas E, Gopalakrishna R (2015) Polyphenols from green tea prevent antineuritogenic action of Nogo-A via 67-kDa laminin receptor and hydrogen peroxide. J Neurochem 132:70-84.
Gundimeda U, McNeill TH, Fan TK, Deng R, Rayudu D, Chen Z, Cadenas E, Gopalakrishna R (2014) Green tea catechins potentiate the neuritogenic action of brain-derived neurotrophic factor: role of 67-kDa laminin receptor and hydrogen peroxide. Biochem Biophys Res Commun 445:218-224.
Hong JT, Ryu SR, Kim HJ, Lee JK, Lee SH, Kim DB, Yun YP, Ryu JH, Lee BM, Kim PY (2000) Neuroprotective effect of green tea extract in experimental ischemia-reperfusion brain injury. Brain Res Bull 53:743-749.
Kokubo Y, Iso H, Saito I, Yamagishi K, Yatsuya H, Ishihara J, Inoue M, Tsugane S (2013) The impact of green tea and coffee consumption on the reduced risk of stroke incidence in Japanese population: the Japan public health center-based study cohort. Stroke 44:1369-1374.
Liu W, Zhou Y, Jia Q, Han B, Zhang G (2011) Effects of Fujian tablet on Nogo-A mRNA expression and plasticity of the corticospinal tract in a rat model of focal cerebral ischemia. Neural Regen Res 6:2577-2581.
Ma TC, Campana A, Lange PS, Lee HH, Banerjee K, Bryson JB, Mahishi L, Alam S, Giger RJ, Barnes S, Morris SM, Jr., Willis DE, Twiss JL, Filbin MT, Ratan RR (2010) A largescale chemical screen for regulators of the arginase 1 promoter identifies the soy isoflavone daidzeinas a clinically approved small molecule that can promote neuronal protection or regeneration via a cAMP-independent pathway. J Neurosci 30:739-748.
Mandel SA, Avramovich-Tirosh Y, Reznichenko L, Zheng H, Weinreb O, Amit T, Youdim MBH (2005) Multifunctional activities of green tea catechins in neuroprotection. Modulation of cell survival genes, iron-dependent oxidative stress and PKC signaling pathway. Neurosignals 14:46-60.
Qin X, Kang L, Liu Y, Huang Y, Wang S, Zhu J (2012) Chinese Medicine’s Intervention Effect on Nogo-A/NgR. Evid Based Complement Alternat Med 528482:528482.
Rolando C, Parolisi R, Boda E, Schwab ME, Rossi F, Buffo A (2012) Distinct roles of Nogo-a and Nogo receptor 1 in the homeostatic regulation of adult neural stem cell function and neuroblast migration. J Neurosci 32:17788-17799.
Schwab ME, Strittmatter SM (2014) Nogo limits neural plasticity and recovery from injury. Curr Opin Neurobiol 27c:53-60.
Tachibana H, Koga K, Fujimura Y, Yamada K (2004) A receptor for green tea polyphenol EGCG. Nat Struct Mol Biol 11:380-381.
Tian W, Han XG, Liu YJ, Tang GQ, Liu B, Wang YQ, Xiao B, Xu YF (2013) Intrathecal epigallocatechin gallate treatment improves functional recovery after spinal cord injury by upregulating the expression of BDNF and GDNF. Neurochem Res 38:772-779.
Wang Y, Li M, Xu X, Song M, Tao H, Bai Y (2012) Green tea epigallocatechin-3-gallate (EGCG) promotes neural progenitor cell proliferation and sonic hedgehog pathway activation during adult hippocampal neurogenesis. Mol Nutr Food Res 56:1292-1303.
10.4103/1673-5374.180729 http://www.nrronline.org/
How to cite this article: Fan TK, Gundimeda U, Mack WJ, Gopalakrishna R (2016) Counteraction of Nogo-A and axonal growth inhibitors by green tea polyphenols and other natural products. Neural Regen Res 11(4):545-546.

- 中国神经再生研究(英文版)的其它文章
- Local translation of cell adhesion molecules in axons
- Gait deterioration due to neural degeneration of the corticoreticular pathway: a case report
- Repositioning again of zonisamide for nerve regeneration
- A novel technique using hydrophilic polymers to promote axonal fusion
- Examining the properties and therapeutic potential of glial restricted precursors in spinal cord injury
- The choroid plexus-cerebrospinal fluid interface in Alzheimer’s disease: more than just a barrier