
转ACO2基因优质棉生长特性及其对田间昆虫群落的影响
2016-11-28 09:03雒珺瑜吕丽敏王春义朱香镇李春花崔金杰
生态学报 2016年19期
雒珺瑜, 张 帅, 吕丽敏, 王春义, 朱香镇, 李春花, 崔金杰
中国农业科学院棉花研究所/棉花生物学国家重点实验室, 安阳 455000
转ACO2基因优质棉生长特性及其对田间昆虫群落的影响
雒珺瑜, 张 帅, 吕丽敏, 王春义, 朱香镇, 李春花, 崔金杰*
中国农业科学院棉花研究所/棉花生物学国家重点实验室, 安阳 455000
新型转基因棉花的生长特性及其对棉田昆虫群落的影响是转基因棉花环境安全评价的重点内容,也是转基因棉花环境安全评价过程必须评价的内容。选用新型转ACO2基因棉花及其亲本中棉所24为试验材料,于2013年和2014年在河南安阳进行了田间棉花生长势及其对昆虫群落影响的试验。在棉花苗期、蕾期、花铃期和铃期,转基因棉花叶片干重与其亲本均无显著差异,苗期、蕾期叶面积和比叶面积与亲本无显著差异,花铃期、铃期叶面积与比叶面积均显著高于亲本材料,其中2013年分别比对照高27.1%和26.2%,2014年分别高16.0%和19.9%,差异均达显著水平;株高、主茎叶片数、大铃数、小铃数和蕾铃脱落数均与亲本材料相当,但铃期果枝数显著高于亲本材料;3个棉铃虫(HelicoverpaarmigeraHubner)发生高峰期,转优质基因棉花对棉铃虫和甜菜夜蛾幼虫校正死亡率与亲本对照材料无显著差异;昆虫群落、害虫亚群落和天敌亚群落昆虫个体总数均显著高于亲本棉田,两年分别比对照高11.5%、10.5%、33.4%和12.0%、8.1%、63.1%,其中2013年差异达显著水平,2014年由于昆虫发生程度总体较轻,差异不显著,但所属“目”、“科”、“种’、生物多样性指数、优势种昆虫及其优势度与其亲本棉田无显著差异,表明外源基因ACO2导入后,对棉田昆虫发生及其群落结构没有影响,棉花生长势无明显优势,但产量构成性状在部分时期增强。上述研究初步明确了新型转基因棉花生存竞争势态和棉田昆虫群落发生规律与动态,可为新型转基因棉花环境安全评价提供理论依据,同时为转基因棉花环境安全评价积累科学数据。
转ACO2基因棉花;棉花生长;棉铃虫;甜菜夜蛾;生物多样性;优势种与优势度
随着转基因技术的发展,转基因棉花的种类也蜂拥出现[1]。外源基因的导入可能致使作物非预期潜在的多效性或诱变效应[2],从而使作物生理特性和次生物质发生变化,进而影响棉田生态系统和生物多样性[3-4]。单个毒素转基因棉花已不能满足棉花生产的需求,为预防棉铃虫种群对Bt毒素产生抗性,同时为应对工业生产对棉花质量的要求、棉花种植技术的变革和种植区域的转移,抗旱、耐盐碱、抗除草剂、新型抗虫棉花、优质高产、专用型转基因棉花已在全国各实验室展开转化。转基因棉花在带来巨大社会效益和经济效益的同时,环境安全性也不可忽视,随着新型转基因棉花的出现,其对非靶标害虫及其天敌昆虫的潜在影响研究越来越受到关注[5- 6], 围绕转基因棉花对棉田昆虫群落的影响做了大量的研究[7- 13]。目前对转基因棉花环境安全性的研究多集中在外源基因转入棉花后对棉田节肢动物群影响,而外源基因主要集中于转Bt基因,其它转基因棉花对棉田节肢动物群落影响仅有少数学者进行了研究[14]。
自2008年转基因新品种保护项目的大力支持下,国内涌现出一批新型转基因棉花新材料,本文对新型转优质基因棉花的生长势及其对棉田昆虫群落的影响的研究,以期为转基因棉花环境安全风险评估积累科学数据和提供技术支撑。
1 试验材料与方法
1.1 棉花材料
转ACO2基因棉花12HN- 4及其亲本非转基因棉花中棉所24均由中国农业科学院棉花研究所生物技术研究室转基因课题组提供,该转基因材料已进入中间试验阶段。ACO2基因为乙烯合成基因,来自于棉花纤维组织,有利于细胞伸长发育,提高棉花纤维品质[15]。试验采用随机区组试验,每棉花品种3个小区,每小区面积80 m2,株距28 cm,行长80 cm,试验地周围设有围墙,并派专人看管,除了试验收留棉籽单独轧花和保存外,其余均混收,秸秆粉碎还田。
1.2 叶面干重、叶面积及比叶面积测定
在棉花生长的苗期、蕾期、花铃期和铃期,分别采集转基因优质基因棉花12HN- 4及其亲本中棉所24顶部倒二叶,测量叶面积后,105 ℃杀青30 min移至80 ℃烘干,称量干重,比叶面积由叶面积与叶片的干重比值推导得到。每小区采集5片叶片。
1.3 棉花产量构成性状调查
在棉花生长的苗期、蕾期、花铃期和铃期,每小区选择具有代表性的连续10株棉花,测定花株高,调查棉花主茎叶片数、单株大铃数、小铃数和蕾铃脱落数。
1.4 棉铃虫和甜菜夜蛾生物测定
试虫来源 棉铃虫由中国农业科学院棉花研究所养虫室提供,系2012年9月采自棉田,在室内(温度(27±0.5)℃,相对湿度 70%—80%,光照周期;L∶D=14 h∶10 h)连续人工饲料饲养。试验用棉铃虫为1d棉铃虫幼虫。
生物测定 参照国家标准《转基因植物及其产品环境安全检测抗虫棉花第1部分:对靶标害虫的抗虫性》(农业部1943号公告- 3—2013)[16],在棉花苗期(二代棉铃虫发生期)、蕾期(三代棉铃虫发生期)和花铃期(四代棉铃虫发生期),采集棉花顶部嫩叶,每小区采集10片,每片叶片放入生物测定盒(直径10cm,高8cm),人工接入5头棉铃虫初孵幼虫或甜菜夜蛾初孵幼虫,置于恒温生长箱(温度(27±0.5)℃,相对湿度70%—80%,光照周期;L∶D=14 h∶10 h))5d后调查活虫数,计算死亡率。
1.5 害虫及其天敌昆虫调查方法
试验采用对角线五点取样方法[16],各处理棉田从5上旬开始至8月底,每5d调查1次2种棉田主要害虫及其天敌种群数量。每小区每次调查5个样点,每点调查5株棉花,共计25株棉花。详细调查取样范围内植株上昆虫的种类和数量,记录所有直接观察到的节肢动物的名称、发育阶段和数量。田间不易识别或者未知种类按统一编号进行记载归类。试验区棉花全生育期不进行化学农药防治,其他农艺措施按常规管理。
1.6 数据分析方法
1.6.1 昆虫群落分析
棉田昆虫“目”、“科”和“种”参照昆虫分类学标准进行分类,并统计分析各盐分处理棉田昆虫群落的结构与组成(科、目、种和个体数量)。
(1)个体总数用N表示
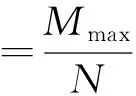
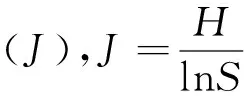
1.6.2 数据统计方法
利用 Excel 软件对数据进行基本处理和筛选,针对研究区测定和调查分析结果,运用 SPSS 17.0 软件对所有数据进行统计分析,采用单因素方差分析(One-way analysis,ANOVA) 和 Duncan′s 差异显著性分析,检验同一时期不同小区内棉花生长相关参数、棉田昆虫结构与组成、生物多样性等显著性差异分析。
2 结果与分析
2.1 转ACO2基因棉花对棉花叶片干重、面积及比叶面积的影响
在棉花生长的苗期(05-28)、蕾期(06-20)、花铃期(07-25)和铃期(08-20),转基因优质棉花12HN- 4叶片干重与其亲本中棉所24在4个棉花关键生长时期均无显著差异;苗期和蕾期转基因棉花叶面积和比叶面积与亲本无显著差异,但花铃期和铃期叶面积和比叶面积均显著高于其亲本材料中棉所24,2013年分别高27.1%和26.2%,2014年分别高16.0%和19.9%,差异均达显著水平(表1)。表明ACO2基因转入棉花后,棉花叶面积和比叶面积增大,引起棉花叶片碳代谢增强,从而增强了棉花的营养生长;2013年与2014年变化趋势一致,总体上2014年棉花生长势稍优于2013年,但差异不显著。2013年叶片干重、叶面积和比叶面积总体上低于2014年。
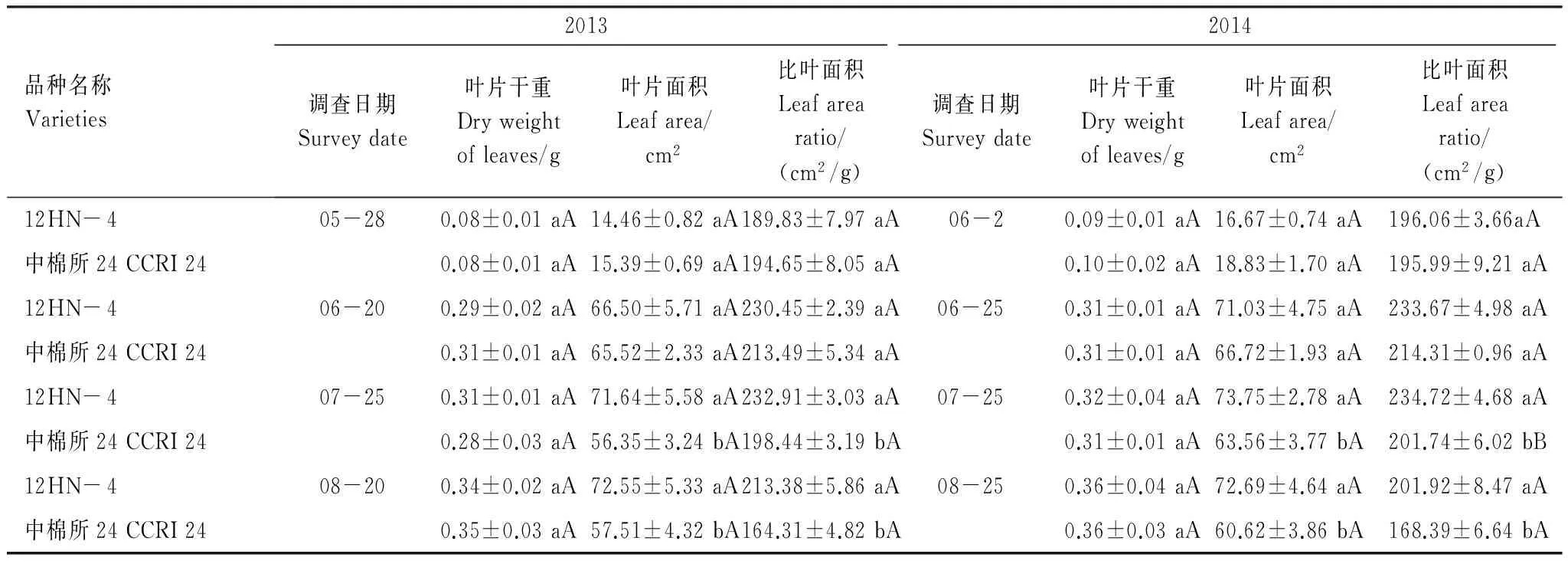
表1 转ACO2基因棉花对棉花叶片干重、面积及比叶面积的影响(2013—2014)
同列不同大小写字母表示品种间差异极显著和显著(P<0.01,P<0.05)
2.2 转ACO2基因棉花对棉花产量构成性状的影响
转基因棉花12HN- 4棉花株高、主茎叶片数、大铃数、小铃数和蕾铃脱落数均高于其亲本材料中棉所24,但差异均未达显著水平;2013年铃期转基因棉花12HN- 4果枝数显著高于其亲本材料。外源基因转入后,不仅增强了棉花的营养生长势,其生殖生长势也得到了增强,主要表现在果枝数的显著增多和铃数的增加(表2)。2a试验表明,2013年转基因棉花及亲本材料生长势均弱于2014年。
2.3 转ACO2基因棉花对棉铃虫和甜菜夜蛾幼虫校正死亡率的影响
在棉花生长的关键时期苗期(二代棉铃虫发生期)、蕾期(三代棉铃虫发生期)和花铃期(三代棉铃虫发生期),转基因棉花12HN- 4对棉铃虫和甜菜夜蛾的幼虫校正死亡率与亲本对照材料无显著差异(表3),表明外源基因ACO2基因转入后,没有改变棉花对棉田主要鳞翅目害虫的控制作用,主要对棉花的生长和品质有一定的增强作用。
2.4 转ACO2基因棉花对棉田昆虫结构与组成的影响
棉花生长势不同,棉田昆虫群落的结构与组成、昆虫种类等不同。转优质基因棉花12HN- 4棉田昆虫群落、害虫亚群落和天敌亚群落昆虫个体总数均显著高于其亲本对照棉田中棉所24,2013年分别高11.5%、10.5%和33.4%,差异均达显著水平,2014年分别高12.0%、8.1%和63.1%,差异均未达显著水平,表明转基因棉花有利于棉田害虫的取食和危害,从而引起食物链上天敌昆虫数量的增多;昆虫群落、害虫亚群落和天敌亚群落所属的目、科和种数与其亲本对照棉田无显著差异,表明转基因棉花对棉田害虫和天敌种类数的变化没有显著的影响(表4)。
2013年昆虫群落、害虫亚群落和天敌亚群落昆虫个体总数高于2014年,其余昆虫结构与组成各参数两年无显著差异。
2.5 转ACO2基因棉花对棉田生物多样性的影响
与对照棉田生物多样性相比,2013年和2014年转基因棉花对棉田昆虫群落、害虫亚群落和天敌亚群落昆虫的多样性指数、均匀性指数和优势集中指数无显著差异(表5),表明外源基因的转入,对棉田生物群落各营养层的多样性指数、均匀性指数和优势集中性指数均无明显的影响,即其棉田昆虫群落的稳定性无显著变化。

表2 转ACO2基因棉花对棉花株高及产量构成性状的影响(2013—2014)
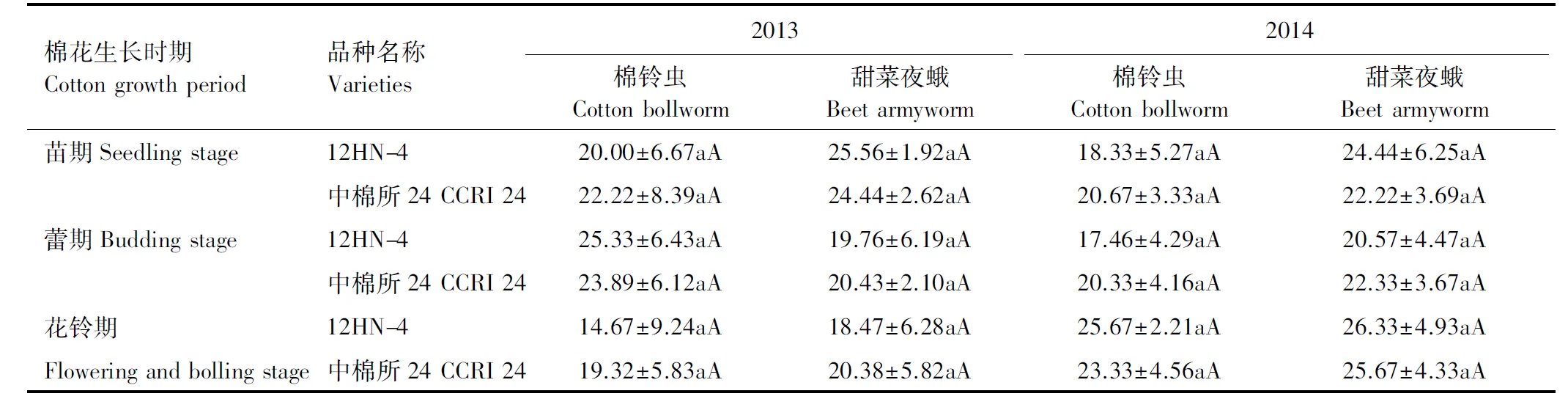
表3 转ACO2基因棉花对棉铃虫和甜菜夜蛾幼虫校正死亡率的影响(2013—2014)
同列不同大小写字母表示品种间差异极显著和显著(P<0.01,P<0.05)
2.6 转ACO2基因棉花对棉田优势种昆虫优势度的影响
2013年和2014年两年试验结果均发现棉田主要优势种害虫依次分别为棉蚜、烟粉虱和蓟马,在棉田害虫种群中占有主导地位。棉田主要天敌昆虫有小花蝽、草间小黑蛛、龟纹瓢虫和蚜茧蜂等。与对照品种相比,转基因棉田优势种害虫和优势种天敌种类相似,优势种的优势度无显著差异,但除了棉蚜和棉叶蝉2013年优势度低于2014年,其余昆虫的优势度2013年均高于2014年(表6)。表明外源基因ACO2导入后,对棉田昆虫优势种和优势度没有明显的影响。
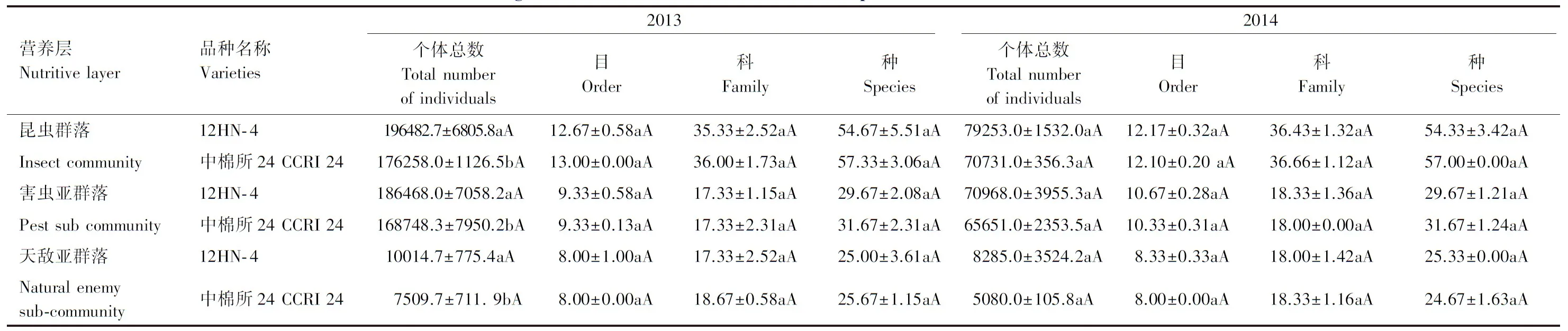
表4 转ACO2基因棉花对棉田昆虫结构与组成的影响(2013—2014)
同列不同大小写字母表示品种间差异极显著和显著(P<0.01,P<0.05)
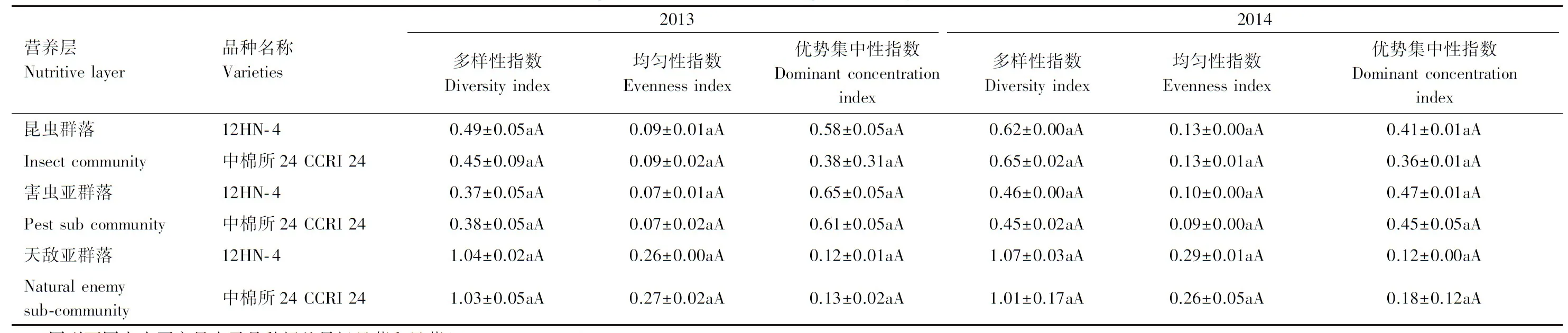
表5 转ACO2基因棉花对棉田生物多样性的影响(2013—2014)
同列不同大小写字母表示品种间差异极显著和显著(P<0.01,P<0.05)

表6 转ACO2基因棉花对棉田优势种昆虫优势度的影响(2013—2014)
同行不同大小写字母表示品种间差异极显著和显著(P<0.01,P<0.05)
3 讨论与结论
在棉花纤维发育过程中,ACO2基因是棉花乙烯合成关键基因,参与细胞的伸长发育,并在植物生长发育和抗逆方面起着重要的作用[15]。本文研究结果表明,ACO2基因转入棉花后,棉花的生长势没有表现出明显的优势,对产量构成性状只在铃期表现出果枝数显著增多,可见在高产、优质品种培育方面具有非常大的应用潜力[17]。
棉田是一种人工干扰下的生态系统,棉田昆虫群落是一个以棉花为中心多种害虫与其天昆虫共存的复杂系统[18]。棉田昆虫群落结构与组成的研究,是科学实施棉田害虫综合治理和充分保护利用天敌自然控害作用的重要基础。棉花品种、农田布局、害虫防治、水肥管理、外源基因等都会一定程度上影响棉田昆虫群落的结构与动态,而昆虫群落物种的组成与结构也会直接影响棉花的生长和发育[19]。研究结果表明,转ACO2基因棉花种植后,棉田昆虫群落、害虫亚群落和天敌亚群落的结构和组成与对照棉田无显著差异,但昆虫个体总数显著高于对照棉田,主要是因为转基因棉花的叶片面积与比叶面积较对照棉花增大,其棉花叶片的碳代谢良好,有利于棉田植食性害虫的取食和危害,为植食性害虫提供了良好的食物资源,从而引起食物链上天敌昆虫数量的增多。
外源基因的导入,作物非预期潜在的多效性或诱变效应也可能会发生改变[20],新型转基因棉花种植后,植株为了表达更高效的目的性状,其他营养物质、次生代谢产物及相关酶活性可能也会因此而改变,进而影响棉田昆虫群落的优势种和优势度,甚至引起棉田生态系统和生物多样性的变化[21]。研究结果表明,转ACO2基因棉花种植后,棉田昆虫群落、害虫亚群落和天敌亚群落多样性指数、均匀性指数和优势集中性指数均与对照棉田无显著差异,这与Saxena and Stotzky[22]、Baumgarte and Tebbe[23]、Marvier[24]、 O′Callaghan等[25]、Hilbeck 等[26]、Torres[27]的研究结果基本一致,也与雒珺瑜[17]研究的转RRm2基因棉花对棉田生物群落多样性影响和雒珺瑜等[28]转Cry1Ac+Cry2Ab棉田和常规棉田节肢动物群落、害虫亚群落和天敌亚群落的多样性指数均无明显差异的研究结果一致。
棉田生境不同,为棉田昆虫提供的食物资源和生活栖息地不同,棉田害虫及其天敌优势种和优势度不同[29];有研究表明,棉花生长势差,或棉花被害虫取食为千疮百孔后,其他害虫再次取食的时间和几率大大减少,即棉花生长势弱[30- 32],昆虫可利用的食物资源减少[33],棉田昆虫种类和数量减少,昆虫数量、优势种、物种丰富度、昆虫群落多样性会发生一定的变化[34],进而影响到食物链上天敌昆虫种类和数量的变化[35]。本文研究结果表明,转ACO2基因棉对主要鳞翅目害虫棉铃虫和甜菜夜蛾没有明显的控制作用,棉田主要优势种昆虫及其优势度与对照棉田无显著差异,表现出棉铃虫已不是棉田的主要害虫,而棉蚜、烟粉虱和蓟马等刺吸性害虫已上升为主要害虫,并在棉田连年重发危害。主要是由于大面积Bt棉的常年连续种植对棉铃虫等非Bt田主要害虫如棉铃虫和甜菜夜蛾等有较好的防控作用,使棉铃虫等害虫的种群数量大量减少,产生生态位的空缺,使一些非靶标害虫从次要害虫转变为主要害虫,从而导致重大经济损失[36]。
棉田生态系统节肢动物群落多样性受气候、外界逆境(虫害、外源杀虫基因、干旱、涝渍)等多种因素的影响[37],而外界因素及食物关系可引起昆虫群落内部害虫及其天敌种群动态变化,而昆虫群落的时空变化,会引起优势种发生演替,导致昆虫群落的多样性和稳定性也会发生变化[38- 39]。本文从棉田昆虫个体总数可见,2014年棉田昆虫数量总体上发生较轻,虽然2014年与2013年相比,棉花生长势和生物多样性等各项指标整体上升高,但虫源总数较低,引起其他昆虫种群数量及优势度等较2013年高。虽然在年度间有部分差异,可能是由于年度间单个或多个气候因素的共同变化而引起。
棉花-害虫-天敌三者的关系十分复杂,而且容易受环境、气候和人为活动等多种因素的影响。同时由于转基因生物风险特征决定了风险的复杂性,而目前转基因棉花对棉田昆虫群落多样性的安全性尚未达成一致的结论,外源基因的风险短期内较难判定,且植物的次生代谢是植物在长期协同进化过程中产生的自身具有抗虫性的物质,与棉田昆虫的发生也具有一定的关系。外源基因的导入,其次生代谢物质是否发生了变化,有必要从外源基因——植物生理-昆虫-天敌相互关系方面进行深入系统研究,对转基因棉花环境安全性的风险从其生理生态的角度进行评估,有助于更深入一层的了解棉田生态系统的细微变化,更完全的解释棉田生态系统昆虫生态位演化规律。
[1] 刘晨曦, 吴孔明. 转基因棉花的研发现状与发展策略. 植物保护, 2011, 37(6): 11- 17.
[2] Ren Y F, Wang T, Peng Y F, Xia B, Qu L J. Distinguishing transgenic from non-transgenicArabidopsisplants by1HNMR-based metabolic fingerprinting. Journal of Genetics and Genomics, 2009, 36(10): 621- 628.
[3] 杨世勇, 宋芬芳, 谢建春. 茉莉酸诱导棉花幼苗抗虫性对棉铃虫相对生长率的影响. 西北农林科技大学学报: 自然科学版, 2013, 41(5): 66- 74.
[4] 雒珺瑜, 崔金杰, 王春义, 辛惠江, 张帅, 吕丽敏. 棉花叶片蛋白质、可溶性糖和花青素含量及其与绿盲蝽抗性的关系. 西北农林科技大学学报: 自然科学版, 2011, 39(8): 75- 80.
[5] Rasmann S, Köllner T G, Degenhardt J, Hiltpold I, Toepfer S, Kuhlmann U, Gershenzon J, Turlings T C J. Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature, 2005, 434(7034): 732- 737.
[6] Quist D, Chapela I H. Transgenic DNA introgressed into traditional maize landraces in Oaxaca, Mexico. Nature, 2001, 414(6863): 541- 543.
[7] Zwahlen C, Nentwig W, Bigler F. Hilbeck A. Tritrophic interactions of transgenicBacillusthuringiensiscorn,Anaphothripsobscurus(Thysanoptera: Thripidae), and the predatorOriusmajusculus(Heteroptera: Anthocoridae). Environmental Entomology, 2000, 29(4): 846- 850.
[8] Dutton A, Klein H, Romeis J, Bigler F. Uptake of Bt-toxin by herbivores feeding on transgenic maize and consequences for the predatorChrysoperlacarnea. Ecological Entomology, 2002, 27(4): 441- 447.
[9] Al-Deeb M A. Effect ofBtcorn expressing the Cry3Bb1 toxin for corn rootworm control on aboveground nontarget arthropods. Environmental Entomology, 2003, 32(5): 1164- 1170.
[10] Jasinski J R, Eisley J B, Young C E, Kovach J, Willson H. Select nontarget arthropod abundance in transgenic and nontransgenic field crops in Ohio. Environmental Entomology, 2003, 32(2): 407- 413.
[11] Men X Y, Ge F, Liu X H, Yardim E N. Diversity of arthropod communities in transgenic Bt cotton and nontransgenic cotton agroecosystems. Environmental Entomology, 2003, 32(2): 270- 275.
[12] Sisterson M S, Tabashnik B E. Simulated effects of transgenic Bt crops on specialist parasitoids of target pests. Environmental Entomology, 2005, 34(4): 733- 742.
[13] Sisterson M S, Biggs R W, Olson C, Carrière Y, Dennehy T J, Tabashnik B E. Arthropod abundance and diversity in Bt and non-Bt cotton fields. Environmental Entomology, 2004, 33(4): 921- 929.
[14] Yu H L, Li Y H, Wu K M. Risk assessment and ecological effects of transgenicBacillusthuringiensiscrops on non-target organisms. Journal of Integrative Plant Biology, 2011, 53(7): 520- 538.
[15] 张萍. 棉花乙烯合成关键基因参与细胞伸长发育的功能分析[D]. 石河子: 石河子大学, 2010.
[16] 转基因植物及其产品环境安全检测抗虫棉花第1部分: 对靶标害虫的抗虫性农业部953号公告- 12.4- 2007.
[17] 雒珺瑜, 刘传亮, 张帅, 王春义, 吕丽敏, 李春花, 李付广, 崔金杰. 转RRM2基因棉生长势和产量及对棉田节肢动物群落的影响. 植物生态学报, 2014, 38(7): 785- 794.
[18] Guo Y Y. Progress in the researches on migration regularity of cotton bollworm and relationships between the pest and its host plants. Acta Entomologica Sinica, 1997, 40(S1): 1- 6.
[19] 陈海风. 棉田节肢动物群落的结构及扰动机理的系统研究[D]. 合肥: 安徽农业大学, 2013.
[20] Rengasamy P. Soil processes affecting crop production in salt-affected soils. Functional Plant Biology, 2010, 37(7): 613- 620.
[21] Sengupta S, Patra B, Ray S, Majumder A L. Inositol methyl tranferase from a halophytic wild rice,PorteresiacoarctataRoxb. (Tateoka): regulation of pinitol synthesis under abiotic stress. Plant, Cell & Environment, 2008, 31(10): 1442- 1459.
[22] Saxena D, Stotzky G.Bacillusthuringiensis(Bt) toxin released from root exudates and biomass of Bt corn has no apparent effect on earthworms, nematodes, protozoa, bacteria, and fungi in soil. Soil Biology and Biochemistry, 2001, 33(9): 1225- 1230.
[23] Baumgarte S, Tebbe C C. Field studies on the environmental fate of the Cry1Ab Bt-toxin produced by transgenic maize (MON810) and its effect on bacterial communities in the maize rhizosphere. Molecular Ecology, 2005, 14(8): 2539- 2551.
[24] Marvier M. Improving risk assessment for nontarget safety of transgenic crops. Ecological Application, 2002, 12(4): 1119- 1124.
[25] O′Callaghan M, Glare T R, Burgess E P J, Malone L A. Effects of plants genetically modified for insect resistance on nontarget organisms. Annual Review of Entomology, 2005, 50: 271- 292.
[26] Hilbeck A, Schmidt J E U. Another view on Bt proteins-how specific are they and what else might they do? Biopesticides International, 2006, 2(1): 1- 50.
[27] Torres J B, Ruberson J R. Interactions of Bt-cotton and the omnivorous big-eyed bugGeocorispunctipes(Say), a key predator in cotton fields. Biological Control, 2006, 39(1): 47- 57.
[28] 雒珺瑜, 崔金杰, 张帅, 陆雪君. 转cry1Ac+cry2Ab基因棉对棉田节肢动物群落的影响. 植物保护, 2011, 37(6): 90- 92.
[29] Finke D L, Denno R F. Intraguild predation diminished in complex-structured vegetation: Implications for prey suppression. Ecology, 2002, 83(3): 643- 652.
[30] Wang R S, Kang Y H, Wan S Q, Hu W, Liu S P, Liu S H. Salt distribution and the growth of cotton under different drip irrigation regimes in a saline area. Agricultural Water Management, 2011, 100(1): 58- 69.
[31] Wang R S, Kang Y H, Wan S Q, Hu W, Liu S P, Jiang S F, Liu S H. Influence of different amounts of irrigation water on salt leaching and cotton growth under drip irrigation in an arid and saline area. Agricultural Water Management, 2012, 110: 109- 117.
[32] Kang Y H, Wang R S, Wan S Q, Hu W, Jiang S F, Liu S P. Effects of different water levels on cotton growth and water use through drip irrigation in an arid region with saline ground water of Northwest China. Agricultural Water Management, 2012, 109: 117- 126.
[33] Pedigo L P, Rice M E. Entomology and Pest Management. Illinois: Waveland Press, 2014.
[34] Haddad N M, Tilman D, Haarstad J, Ritchie M, Knops J M H. Contrasting effects of plant richness and composition on insect communities: a field experiment. The American Naturalist, 2001, 158(1): 17- 35.
[35] Martinson H M, Fagan W F. Trophic disruption: a meta-analysis of how habitat fragmentation affects resource consumption in terrestrial arthropod systems. Ecology Letters, 2014, 17(9): 1178- 1189.
[36] Wu K M, Li W D, Feng H Q, Guo Y Y. Seasonal abundance of the mirids,LyguslucorumandAdelphocorisspp. (Hemiptera: Miridae) on Bt cotton in northern China. Crop Protection, 2002, 21(10): 997- 1002.
[37] 刘万学,万方浩,郭建英. 转Bt基因棉田节肢动物群落营养层及优势功能团的组成与变化. 生态学报, 2002, 22(5): 729- 735.
[38] 邝幸泉, 黄汉文, 张国安. 棉红铃虫自然种群的生命统计特征. 昆虫学报, 1993, 36(3): 308- 314.
[39] Romeis J, Dutton A, Bigler F.Bacillusthuringiensistoxin (Cry1Ab) has no direct effect on larvae of the green lacewingChrysoperlacarnea(Stephens) (Neuroptera: Chrysopidae). Journal of Insect Physiology, 2004, 50(2/3): 175- 183.
Effects of transgenic ACO2 cotton on growth characteristics and insect community
LUO Junyu, ZHANG Shuai, LYU Limin, WANG Chunyi, ZHU Xiangzhen, LI Chunhua, CUI Jinjie*
InstituteofCottonResearchofCAAS/StateKeyLaboratoryofCottonBiology,Anyang455000,China
The growth characteristics of transgenic cotton and its effects on the insect communities in cotton fields, which is a key aspect of the evaluation of the environmental safety of transgenic cotton, must be assessed. In this study, we selected the new transgenic ACO2 cotton and its parental CCRI 24 as samples to test cotton growth and changes in insect community in the city of Anyang in Henan province in 2013 and 2014. The results showed that, the leaf dry weight was not significantly different between transgenic cotton and the parental cotton plant at the seeding, bud, flowering or boll stages. In the seedling and bud stage, the leaf area and leaf area ratio of transgenic cotton was not clearly different from the parent. However, in the flowering and boll stage, the ratios were significantly different from those of the parents, specifically 27.1% and 26.2% higher in 2013, and 16% and 19.9% higher in 2014, respectively. There were no significant differences between the transgenic cotton and its parent plant with respect to plant height, leaf number, boll number, bell number or boll abscission number. However, boll branch number in transgenic plants was significantly higher in the parents at the boll stage. During the three adult peaks of occurrence ofHelicoverpaarmigera, the corrected mortality of cotton bollworm and beet armyworm in transgenic cotton was much lower but was not significantly different from the parental control. The total number of individuals comprising the insect, pest sub, and natural enemy sub communities in the transgenic cotton field were higher than that in parent cotton field, at 11.5%, 10.5%, 33.4% and 8.1%, 63.1%, 12% in 2013 and 2014, respectively, with a significant difference in 2013. Owing to the diminished occurrence of insects in 2014, the difference was not significant. However, the order, family, and species as well the indices for biodiversity, evenness, and dominant species were not significantly different. The above results demonstrated that no obvious advantage was apparent in the new transgenic cotton in respect to growth potential after the ACO2 gene was imported into the cotton. However, the yield component traits in some periods were enhanced. This preliminary study illuminates the rules and dynamics and competition of insect communities as well as the survival of transgenic cotton plants in the field. This could provide a theoretical basis for the evaluation of new transgenic cotton and environmental safety, and this could be the beginning of the accumulation of data concerning both these topics.
ACO2 transgenic cotton; cotton growth; cotton bollworm; beet armyworm; biodiversity; dominant species and dominance
转基因生物新品种培育重大专项(2016ZX08011-002)
2015- 05- 13;
日期:2016- 01- 15
10.5846/stxb201505130976
*通讯作者Corresponding author.E-mail: cuijinjie@126.com
雒珺瑜, 张帅, 吕丽敏, 王春义, 朱香镇, 李春花, 崔金杰.转ACO2基因优质棉生长特性及其对田间昆虫群落的影响.生态学报,2016,36(19):6113- 6121.
Luo J Y, Zhang S, Lü L M, Wang C Y, Zhu X Z, Li C H, Cui J J.Effects of transgenic ACO2 cotton on growth characteristics and insect community.Acta Ecologica Sinica,2016,36(19):6113- 6121.
猜你喜欢
长江蔬菜(2021年12期)2021-04-04
中国果业信息(2019年11期)2019-01-05
新疆农垦科技(2016年2期)2016-08-21
西南农业学报(2016年5期)2016-05-17
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年5期)2016-02-28
现代农业(2016年4期)2016-02-28
现代农业(2015年5期)2015-02-28
新疆农垦科技(2014年12期)2014-02-28
新疆农垦科技(2014年10期)2014-02-28
