
坡位对浑善达克沙地小叶锦鸡儿空间分布影响
2016-11-14 11:47宋维峰陈治成
水土保持研究 2016年6期
魏 峥, 王 晓, 宋维峰, 陈治成
(1.西南林业大学 环境科学与工程学院, 昆明 650224; 2.北京林业大学 水土保持学院, 北京 100083 )
坡位对浑善达克沙地小叶锦鸡儿空间分布影响
魏 峥1, 王 晓2, 宋维峰1, 陈治成1
(1.西南林业大学 环境科学与工程学院, 昆明 650224; 2.北京林业大学 水土保持学院, 北京 100083 )
坡位对植物种子散布、繁殖和土壤水、热与营养再分配等生态、水文方面各有差异,不适坡位使其生态功能被破坏,造成水土流失、生物多样性降低等失衡生态结构。分析坡位对植物种群的空间分布格局的影响,有助于认识其潜在的生态学过程、种群的生物学特性及其与环境因子之间的相互关系,为合理规划不同坡位植被类型提供依据,对于荒漠化地区意义重大。采用空间格局分析方法,将浑善达克沙地坡位对小叶锦鸡儿空间分布的影响进行了分析。结果表明:(1) 在坡上及平地部位,小叶锦鸡儿种群在小尺度上呈现高度聚集分布,随着尺度增大,聚集强度逐渐减弱,最终表现为随机分布,而在坡中及坡下部位,小叶锦鸡儿种群在小尺度上表现为均匀或随机分布;(2) 根据冠幅、株高将小叶锦鸡儿分为大苗、幼苗,从坡上、坡中、坡下到平地,小叶锦鸡儿大苗灌丛沙堆与小叶锦鸡儿幼苗灌丛沙堆均表现为负相关关系,随着尺度增大,这种关系逐渐减弱,最后表现为无关联。
空间分布格局; 坡位; 小叶锦鸡儿; 浑善达克沙地
西北干旱、半干旱地区是我国典型的荒漠系统,它不仅是我国北方重要的生态屏障,也是我国六大生物多样性中心之一,拥有多种荒漠生态系统特有的科、属、种。近年来,由于全球气候变暖、过度放牧等不合理的土地利用政策,该地区出现了大规模的水土流失、荒漠化、草地退化等生态环境问题,严重影响到当地居民的生存、生产和生活[1]。灌木是荒漠区植被的主要组成部分,作为这一区域的优势物种,在维持荒漠生态系统的生物多样性、生态服务功能及稳定性等方面具有重要的作用[2],此外,灌木还是退化荒漠生态系统恢复与重建的关键生物资源,在发展畜牧业、改良土壤、防风固沙等方面发挥了不可代替的重大效能。尽管在过去的二十年,灌木入侵已作为土地退化的一个指示因子[3],但近些年来这种观点已有所改变[4],适生灌木已成为区域生态环境建设重要的营林类型。由于我国北方干旱、半干旱地区降水量偏少,大部分地区年降水量低于400 mm,乔木难以大面积栽植,从而决定了这些区域生态恢复主要以草本与灌木为主。在过去的几十年,我国在干旱、半干旱地区营造了大量的水土保持、防风固沙灌木林,对当地的水土保持、生态恢复起到了巨大作用[5]。而锦鸡儿属作为我国北方地区主要的防风固沙和水土保持造林树种,同时也是“三北”防护林体系建设中的一个主要灌木树种,在西北地区的水土保持、水源涵养、护沟护坡、防风固沙中发挥着巨大的作用。但随着林龄的增加,锦鸡儿林普遍出现林分衰老、生物量下降、林分老化等现象,其经济效益和生态效益不断下降[6]。如何针对不同区域不同类型的灌木林进行可持续经营及科学恢复,成为当前需要解决的主要问题。
植物种群的空间格局是指植物种群个体在水平空间的分布状况[7],是植物种群生物学特性对环境条件长期适应和选择的结果,也是研究种群间相互作用及种群与环境间相互关系的重要手段[8]。分析植物种群的空间分布格局有助于认识其潜在的生态学过程(如种子扩散、种内和种间竞争、干扰等)、种群的生物学特性(如生活史策略、喜光、耐阴等)及其与环境因子之间的相互关系(如小生境、植物与生长环境之间的适合度、环境异质性等)[9]。植被的空间分布是由多种环境因素在不同的空间尺度上决定的。地带性气候决定了全球范围内的植被空间分布格局,而在景观或更微细的尺度上,非地带性因素是决定植被空间分布的主要因素[10]。坡位作为一个重要的非地带性因素,一方面通过地貌的起伏直接改变植被的空间分布,另一方面,通过改变(光、热、水)等资源的再分配等因素,从而间接对植被的空间分布产生影响,因而坡位对植被的影响得到越来越多的关注。在水土流失及荒漠化治理中,灌木往往作为裸露坡面及流动沙丘恢复的首选材料,这些灌木能迅速固定斜坡坡面或形成灌丛带,从而作为宿主植物为其他植物提供存活条件,形成复杂的植物群落最终促使形成稳定的坡面或固定沙地。
本研究的主要问题如下:(1) 不同坡位条件下,小叶锦鸡儿种群的空间格局分布情况;(2) 不同坡位条件下,小叶锦鸡儿种群的种内竞争情况。通过这两个问题的研究,为不同地形条件下人工小叶锦鸡儿退化林的修复提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于浑善达克沙地,属于内蒙古正蓝旗,地理坐标为东经115°16′E,北纬42°50′N,植物区系为荒漠草原地带。海拔大约为1 320 m左右,属暖温带大陆季风气候,多年平均降水量在250~350 mm,其中70%以上降水集中在6—8月,年平均温度为1.7℃,极端最低温度为-38℃,年积温(≥10℃)是2 000℃,无霜期为105 d,年日照时间大于1 000 h,年平均风速4 m/s,西北风为主导风向。土壤类型主要为栗钙土。
小叶锦鸡儿属于多年生旱生灌木,广泛分布于内蒙古草原从东到西的荒漠及草原地带,花期为5月,时常为20 d,种子一般在7月份成熟。种子萌发主要有夏季降雨触发。
1.2 样地调查
在研究样地,选取一块以自然生长小叶锦鸡儿为主要建群种的固定斜坡,沿坡面自上而下,将坡面分成坡上、坡中、坡下和平地四个不同的坡位,在不同的坡面,分别建立一块100 m×100 m的固定样地,将每个样地划分为400个5 m×5 m的小样方,以样地固定1角为起始原点(0,0),记录每个固定样地内小叶锦鸡儿种的位置、高度、长短冠幅等,同时标记坡度、坡位、土壤等其他自然因子。
由于灌木没有标准的胸径,而在野外小叶锦鸡儿种群一般呈灌丛化趋势,无法判断其年龄,为分析小叶锦鸡儿种群的种内竞争关系,结合一些研究情况,根据灌木株高、长冠幅、短冠幅的均值将小叶锦鸡儿区分为大苗(均值≥40 cm),幼苗(<40 cm)。
1.3 统计分析方法
点格局分析法是把植物个体都视为二维空间平面中的点[11],Ripley′sK函数和g(r)函数都是常用的点格局分析方法。相比较于Ripley′sK函数,g(r)函数是用指定宽度的圆环区域取代了Ripley′sK函数中半径为R的圆形区域,从而减少了Ripley′sK函数的累积效应,在较小的尺度上g(r)函数的敏感性更强。在本文研究中,我们采用单变量g11(r)函数和双变量g12(r)函数分析不同坡位小叶锦鸡儿种群的空间分布格局及种内关联。
(1) 单变量格局分析。Ripley于1981年提出了Ripley′sK函数的统计学理论,后经Diggle等人于1983年发展简化为函数方程K(r):
式中:r是空间尺度;A 为样地面积;n为植物总数;wij为边缘效应校正的权重值;Ir(i,j)为指标函数,当dij≤r时为1,否则为0。
式中SOC团储为土壤团聚体有机碳储量,t/hm2;SOC团为土壤团聚体的有机碳含量,g/kg;W团为团聚体质量比例(W团=M团/M土;M团为团聚体质量;M土为土壤质量);r为土壤容重,g/cm3;H为土壤厚度,cm。
g函数由Stoyan于1994年应用K函数推导得到:
式中:r是空间尺度。
在完全随机分布下,g(r)在所有尺度下均应等于1,如果g(r)大于1,那么在对应尺度下,种群为聚集分布;反之,如果g(r)小于1,那么在对应尺度下,种群为均匀分布。
(2) 双变量格局分析。两个种的个体在特定尺度r内的格局可以写为g12(r):
g12(r)也是由K函数派生出来的,它的含义是以格局1的k个点为中心,r为半径的圆环中出现格局2的点的期望数。在完全随机分布下,g12(r)=1,表示两个物种在r尺度下无关联;g12(r)大于1,表示二者正相关;g12(r)小于1,表示二者负相关。
数据分析通过生态学软件Programita[12]完成,采用最小半径50m为研究尺度,通过完全随机空模型(CSR)下MonteCarlo199次模拟分析得到95%的置信区间。
2 结果与分析
2.1 不同坡位条件下小叶锦鸡儿种群空间分布情况
不同坡位条件下小叶锦鸡儿种群表现为不同的空间分布格局。在坡上地段,小叶锦鸡儿种群在0~27.5m的空间尺度上表现为高度聚集分布,但随着研究尺度的增加,聚集强度逐渐减弱,趋于随机分布;在坡中地段,小叶锦鸡儿种群在0~1m的尺度内表现为均匀分布,在1~2m的尺度条件下表现为聚集分布,而在其他尺度内则表现为随机分布;在坡下地段,小叶锦鸡儿种群在0~1m与17.5~50m的尺度内表现为均匀分布格局,在1.5~8.5m的尺度条件下表现为高度聚集分布状态,而在其他尺度条件下表现为随机分布;在平地地段,小叶锦鸡儿种群在0~8.5m的尺度内表现为高度聚集分布,随着尺度的增大,在其他范围内则表现为随机分布。综上可知,坡上与平地的小叶锦鸡儿种群在小尺度上呈现高度聚集分布的情况,随着尺度的增大,聚集程度减弱,逐渐趋于随机分布;对于坡中与坡下地段,小叶锦鸡儿种群主要表现为均匀分布和随机分布的情况,聚集尺度较少(图1)。
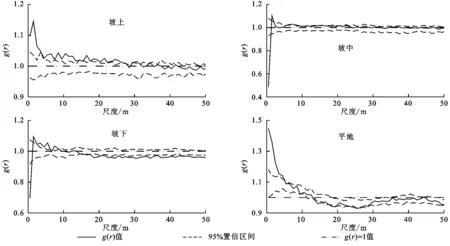
图1 不同坡位下小叶锦鸡儿(整个种群)的空间分布格局
从聚集强度来看,坡上最大聚集强度1.15,坡中最大聚集强度1.1,坡下最大聚集强度1.1,平地最大聚集强度1.48。因此,不同坡位条件下小叶锦鸡儿种群的聚集强度表现为:平地>坡上>坡中=平地。
2.2 不同坡位条件下小叶锦鸡儿种群(大苗)的分布
对小叶锦鸡儿种群大苗空间分布格局进行单变量g(r)函数统计分析,结果表明不同坡位条件下小叶锦鸡儿(大苗),在不同尺度上的空间格局不同,坡上地段的小叶锦鸡儿(大苗),在0~1.5 m尺度范围内表现为随机分布格局,在1.5~48.5 m的尺度上呈现为高度聚集分布格局,在48.5~50 m的尺度范围内则表现为随机分布格局;在坡中地段,小叶锦鸡儿(大苗)在0~1 m的尺度内表现为均匀分布格局,在1~2 m的尺度内表现为聚集分布,而在其他尺度内则表现为随机分布格局;在坡下地段,小叶锦鸡儿(大苗)在0~1 m的尺度内表现为均匀分布,在其他尺度内表现为随机分布;在平地地段,小叶锦鸡儿(大苗)在0~12.5 m的尺度内表现为高度聚集分布,在17.5~28.5 m的尺度内为均匀分布,在其他范围内表现为随机分布(图2)。
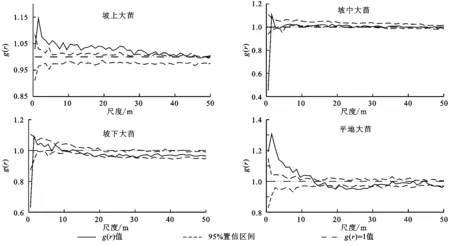
图2 不同坡位条件下小叶锦鸡儿(大苗)的空间分布格局
从最大聚集强度来看,坡上最大聚集强度1.15,坡中最大聚集强度1.1,坡下最大聚集强度1.09,平地最大聚集强度1.31。聚集强度表现为:平地>坡上>坡中>坡下。
2.3 不同坡位条件下小叶锦鸡儿种群(幼苗)的分布
对于小叶锦鸡儿种群的幼苗空间分布格局进行单变量g(r)函数统计分析(见图3),结果显示,坡上地段的幼苗在0.5~17.5 m的尺度上表现为高度聚集分布,在其他尺度内表现为随机分布;在坡中地段,在0.5~2.5 m,10.5~12.5 m的尺度内表现为高度聚集分布,其他尺度内表现为随机分布;在坡下地段,小叶锦鸡儿在0.5~2.5 m,10.5~11.5 m的尺度内表现为高度聚集分布,在其他尺度内表现为随机分布;在平地地段,小叶锦鸡儿种群在0.8~8.5 m的尺度内表现为高度聚集分布,在其他范围内表现为随机分布。
从最大聚集强度来看,坡上最大聚集强度7,坡中最大聚集强度3.5,坡下最大聚集强度14,平地最大聚集强度9.2。最大聚集强度表现为:坡下>平地>坡上>坡中。
2.4 不同坡位条件下小叶锦鸡儿种群种内竞争的分布
在坡上地段,小叶锦鸡儿(大苗)与小叶锦鸡儿(幼苗)在4.5~7.5 m及17.5~50 m的尺度内表现为负相关,其他尺度内无关联;在坡中地段,小叶锦鸡儿(大苗)与小叶锦鸡儿(幼苗)在0~0.5 m的尺度内表现为负相关,在其他尺度内无关联;在坡下地段,小叶锦鸡儿(大苗)与小叶锦鸡儿(幼苗)在18.5~50 m的尺度内表现为高度负相关,在其他尺度内无关联;在平地地段,小叶锦鸡儿(大苗)与小叶锦鸡儿(幼苗)在0~1.5 m的尺度内表现为正相关,在9.5~19.5 m的尺度内表现为高度负相关,其他尺度内无关联(图4)。

图3 不同地形条件下小叶锦鸡儿(幼苗)的空间分布格局
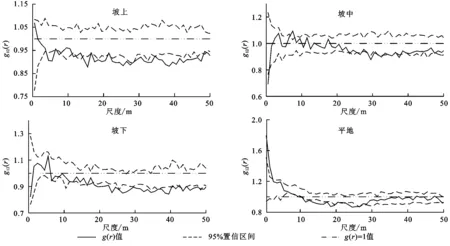
图4 不同地形条件下小叶锦鸡儿(大苗—幼苗)的空间关联分析
3 讨 论
3.1 坡位对小叶锦鸡儿种群空间分布的影响
植物种群的空间格局不仅因种而异,而且同一个种在不同发育阶段、不同生境条件下也有明显差异,我们通过分析物种种群的空间分布格局,可以研究物种分布与生境的相互关系,在本文中我们主要探讨不同坡位条件下小叶锦鸡儿的空间分布情况。如上研究可知:坡上与平地的小叶锦鸡儿种群在小尺度表现为高度聚集分布,随着尺度的增大,聚集强度逐渐减弱,最后趋于随机分布。一般情况下,植物以母株为中心的繁殖方式使低龄植株呈现聚集分布,随植株年龄的增加,对光因子和营养条件的要求加大加之自疏、它疏等作用,种群会趋于均匀分布,随年龄的继续增加,种群最终将趋于随机分布。在干旱、半干旱地区,水分是影响植被生长的重要因素[13],而地貌通过自身的起伏等作用,不仅影响了自身土壤水分的分布,还会对降雨进行再分配,从而最终决定了其上灌木对水分的利用效率。由于重力作用,坡上部位雨水储存量较小,大部分雨水顺流而下,储存在坡中下部,这就造成了不同坡位间水分分布的空间异质性。在坡中下部位,由于水分条件较好,小叶锦鸡儿种群盖度高,个体较大,由于种内竞争的不对称性,小叶锦鸡儿种群与其他个体争夺水分、土壤等资源,相邻的植株容易产生排斥,这导致新生的幼苗往往与母体保持一定距离,从而使得小叶锦鸡儿种群聚集程度较低,整个群落呈现一种稳定的状态,在空间分布格局上,就表现为小尺度均匀分布,而大尺度随机分布的情况;反之,在坡上部位,水分条件较差,小叶锦鸡儿种群个体较小,盖度较低,由于较小个体对资源的竞争较弱,以及幼小锦鸡儿应对恶劣环境能力较弱,个体依赖性增强,植物种群往往呈现聚集分布;而对于平地部位,一方面平地部位由于太阳直射,同等区域的蒸发量明显大于坡中下部,另一方面平地部位的小叶锦鸡儿受人为干扰的程度最强,从而导致在平地部位小叶锦鸡儿种群呈现高度聚集分布的状态。反过来说:由于坡上部位植被盖度和生物量低于坡中、坡下部位,对雨水的拦蓄作用小,导致坡顶、坡上部位地表径流系数低于坡中、坡下部位,这也造成坡中下部位水分条件好于坡上部位。
从小叶锦鸡儿种群大苗、幼苗的空间分布来看,通常幼小个体对水分等资源的竞争相对微弱,同时为抵御风沙危害和提高存活机会它们需要相互庇护,往往表现为集群分布[14],这也是为什么在4块样地中,小叶锦鸡儿幼苗均表现为高度聚集分布的原因,而随着龄级的增加,高大个体对土壤水分或其他资源的竞争会加剧,生长过密的个体间会产生自疏效应,同时,由于它们独立抵御风沙危害的能力明显增强,对相互庇护作用的依赖性降低,从而导致聚集强度的降低,表现为均匀分布或者随机分布。这与我们研究中在中下坡小叶锦鸡儿大苗呈现均匀、随机分布的结果一致。然而由于水分条件的限制及人为干扰的增强,在坡上及平地的小叶锦鸡儿大苗,仍然要面对恶劣的生存环境,从而表现为聚集分布。
3.2 坡位对小叶锦鸡儿种群种内相互关系的影响
种内联结用来表示同一物种不同龄级在空间分布上的相互关联性,是对一定时期内植物种群组成个体之间相互关系的静态描述,这种关系不仅包括空间分布关系,也包含着个体间的功能关系,揭示了种群的现状,并能反映种群内个体间的相互作用[15]。而竞争是干旱区生态系统中一种普遍存在的现象。在我们的研究中,在不同的坡位,小叶锦鸡儿大苗与小叶锦鸡儿幼苗几乎都表现为负相关,呈现相互竞争的关系,这是因为在干旱、半干旱区,植被的生长环境恶劣,在自然资源的限制下,幼苗与大苗之间存在激烈竞争,此外,小叶锦鸡儿种群不同于其他树种,在野外,小叶锦鸡儿种群更多的以灌丛化存在,最终形成灌丛化沙包,而灌丛沙包往往产生肥岛效应[16],这就导致灌丛沙包内植物生长的水分及土壤因素明显好于灌丛沙包外,这种异质性的差异导致小叶锦鸡儿大的灌丛沙包与新形成的小灌丛沙包之间往往是一种资源不对等性,这种资源异质性也导致小叶锦鸡儿种群大苗与幼苗之间不是相互依存的关系,而是一种互相竞争的关系,从而导致了大部分坡度上小叶锦鸡儿种群大苗与幼苗的空间关联表现为负相关,而在平地上,由于肥岛效应的不明显,资源差异不大,而小叶锦鸡儿种群可以无性生殖,由根萌发,这就形成了在小尺度上小叶锦鸡儿大苗与幼苗之间反而呈现正相关相互庇护生长的关系。
4 结 论
小叶锦鸡儿作为我国干旱、半干旱区一种常见的水土保持、防风固沙植物,过去几十年被大面积的栽植。由于坡位对降水、养分等自然资源的再分配,导致了不同坡位情况下小叶锦鸡儿种群的空间分布出现明显的差异性。坡中下部的小叶锦鸡儿种群长势较好,空间分布格局相对稳定,而坡上及平地部分的小叶锦鸡儿种群长势较差,相互关系不稳定,在下一步对锦鸡儿属林的抚育管理中,应更加关注坡上及平地部分的林分经营,人为提高这部分立地的水分、土壤情况,改善立地条件。
由于小叶锦鸡儿种群的灌丛化现象导致的资源异质性,致使小叶锦鸡儿种群大苗与幼苗间往往表现为竞争关系,在小叶锦鸡儿林更新及再植过程中,应注意引进新的灌木树种,形成混交的模式,避免单一的小叶锦鸡儿造林,最终导致整个林分结构的不稳定。
[1] 姜晔,毕晓丽,黄建辉,等.内蒙古锡林河流域植被退化的格局及驱动力分析[J].植物生态学报,2010,34(10):1132-1141.
[2] Li X R. Study on shrub community diversity of Ordos Plateau, Inner Mongolia, Northern China[J]. Journal of Arid Environments, 2001,47(3):271-279.
[3] Sankey J B, Ravi S, Wallace C S A, et al. Quantifying soil surface change in degraded drylands: Shrub encroachment and effects of fire and vegetation removal in a desert grassland[J]. Journal of Geophysical Research Biogeosciences, 2012, 117(G2):194-215.[4] D′Odorico P, Okin G S, Bestelmeyer B T. A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands[J]. Ecohydrology, 2012,5(5):520-530.
[5] 王葆芳,王志刚,江泽平,等.干旱区防护林营造方式对沙漠化土地恢复能力的影响研究[J].中国沙漠,2003,23(3):236-241.
[6] 张瑜,郑士光,贾黎明,等.晋西北低效柠条林老龄复壮技术及能源化利用[J].水土保持研究,2013,20(2):160-164.
[7] Dale M R T. Spatial Pattern Analysis is in Plant Ecology[M]. Cambridge:Cambridge University Press, 2000.
[8] 王磊,孙启武,郝朝运,等.皖南山区南方红豆杉种群不同龄级立木的点格局分析[J].应用生态学报,2012,21(2):272-278.
[9] Nathan R. Long-distance dispersal of plants[J]. Science, 2006,313(5788):786-788.
[10] 邝高明,朱清科,刘中奇,等.黄土丘陵沟壑区微地形对土壤水分及生物量的影响[J].水土保持研究,2012,19(3):74-77.
[11] 张金屯,孟东.平芦芽山华北落叶松林不同龄级立木的点格局分析[J].生态学报,2004,24(1):35-40.
[12] Wiegand T, Moloney, K A. Handbook of spatial point-pattern analysis in ecology [M]. Boston:CRC Press, 2013.
[13] 彭海英,李小雁,童绍玉.内蒙古典型草原小叶锦鸡儿灌丛化对水分再分配和利用的影响[J].生态学报,2014,34(9):2256-2265.
[14] 王彦阁,杨晓晖,慈龙骏.西鄂尔多斯高原干旱荒漠灌木群落空间分布格局及其竞争关系分析[J].植物资源与环境学报,2010,19(2):8-14.
[15] 叶尔江,张春雨,赵秀海.天山云杉林优势种空间格局及其空间关联性[J].林业资源管理,2013(3):58-62.
[16] 初玉,杨慧玲,朱选伟,等.浑善达克沙地小叶锦鸡儿灌丛的空间异质性[J].生态学报,2005,25(12):3294-3300.
Impact of Different Terrain Conditions on Population Structure and Pattern ofCaraganaMirophyllain Otindag Sandland
WEI Zheng1, WANG Xiao2, SONG Weifeng1, CHEN Zhicheng1
(1.CollegeofEnvironmentScienceandEngineering,SouthwestForestryUniversity,Kunming650224,China; 2.SchoolofSoilandWaterConservation,BeijingForestryUniversity,Beijing100083,China)
Slope position has different effects on the ecohydrological processes, such as seed dispersal, reproduction, soil water dynamics, heat transfer and nutrient redistribution, etc. In appropriate slope position will unbalance the ecological function, such as soil and water loss and decreased biodiversity, researches about the effect of slope position on the spatial pattern of plant population will contribute to understand the potential ecological processes and biological characteristics of plant populations and their relationships with environmental factors, and be helpful to the rational planning of vegetation types of different slope positions in desertification-prone area. Based on the method of spatial pattern we studied the impact of different slope positions on the structure and pattern ofCaraganamirophyllain Otindag Sandland. The results showed that: (1) the population ofCaraganamirophyllain the upper part of the slope or on the flat part showed high aggregation within the small scale, as the scale increased, the aggregation intensity gradually decreased and finally became random distribution, while in the middle and lower parts of the slope, the population presented uniform or random distribution within the small scale; (2) according to the crown and height,Caraganamirophyllawas divided into the adult and the youth, from the upper, the middle and the lower slope parts down to the flat part, the negative correlation between the adult and the youth existed, with the increase of the scale, the relationship between them gradually weakened and finally showed no association.
spatial pattern; slope position;Caraganamirophylla; Otindag Sandland
2016-01-21
2015-03-12
国家国际科技合作专项“中哈丝绸之路经济带土地退化监测与评价技术合作研究”(2015DFR31130)
魏峥(1987—),女,内蒙赤峰翁牛特旗人,硕士研究生,主要从事生态环境工程研究。E-mail:wei-zheng-wo@163.com
宋维峰(1967—),男,甘肃会宁县人,博士,教授,主要从事生态环境工程和森林水文研究。E-mail:songwf85@126.com
Q14,Q948
A
1005-3409(2016)06-0102-06
猜你喜欢
种子(2022年5期)2022-06-27
黄河之声(2021年6期)2021-06-18
防护林科技(2020年5期)2020-07-21
水土保持研究(2020年1期)2020-04-27
西藏农业科技(2018年4期)2018-04-25
当代工人(2018年21期)2018-03-06
文学港(2018年1期)2018-01-25
绿色科技(2017年3期)2017-03-14
绿色科技(2017年1期)2017-03-01
湖南林业科技(2017年1期)2017-02-06
