
贵州喀斯特森林三种植物对不同坡位环境的光合生理响应
2016-11-11 06:51:33邢德科吴沿友付为国杭红涛赵玉国吴沿胜
广西植物 2016年10期
邢德科,吴沿友,付为国,杭红涛,赵玉国,吴沿胜
( 1. 教育部现代农业装备与技术国家重点实验室,江苏大学 农业装备工程学院,江苏 镇江 212013;2. 环境地球化学国家重点实验室环境生物科技研究中心,中国科学院地球化学研究所,贵阳 550002 )
贵州喀斯特森林三种植物对不同坡位环境的光合生理响应
邢德科1,吴沿友2*,付为国1,杭红涛2,赵玉国1,吴沿胜1
( 1. 教育部现代农业装备与技术国家重点实验室,江苏大学 农业装备工程学院,江苏 镇江 212013;2. 环境地球化学国家重点实验室环境生物科技研究中心,中国科学院地球化学研究所,贵阳 550002 )
该研究以贵州普定喀斯特森林中、下坡位生长的构树(Broussonetiapapyrifera)、朴树(Celtissinensis)和光滑悬钩子(Rubustsangii)为材料,通过对碳酸酐酶(CA)活性、光合作用日变化、净光合速率对CO2与光的响应曲线、叶绿素荧光特性以及稳定碳同位素组成等指标的测定,进而对比分析三种植物不同的光合生理响应特性。结果表明:构树光合作用过程的无机碳源既可来自大气中的CO2,也可以在气孔部分闭合的情况下利用细胞内的HCO3-,下坡位的构树较高的CA活性使其利用HCO3-的效率会更高,并能在较低光强下具有较高的光能利用效率。这可能与下坡位的构树具有较高的CA活性有关,对下坡位具有更好的适应性。朴树光合无机碳的同化能力最低,且光合无机碳源较单一,主要利用大气CO2,其较慢的生长速率使其对无机碳的需求最低,且能保持较稳定的无机碳同化速率。相对来说,中坡位的朴树具有相对较高的净光合速率和光能利用效率,对中坡位表现出较好的适应性。光滑悬钩子主要利用大气中的CO2进行光合作用。中坡位的光滑悬钩子具有较强的光能利用效率,并表现出较高的净光合速率,光滑悬钩子对中坡位同样表现出较好的适应性。该研究结果为喀斯特生态脆弱区植被重建过程中树种的选择及合理配置提供了科学依据。
植被恢复,碳酸酐酶,表观量子效率,羧化效率,稳定碳同位素组成
贵州碳酸盐岩面积达13 × 104km2,占全省土地总面积的73%,是我国喀斯特地貌最发育的省份,同时也是土壤侵蚀严重和生态环境脆弱的地区,而碳酸盐岩山区土壤侵蚀主要发生在山坡,是一种选择性的侵蚀过程(白占国等,1998)。该区面临着经济和生态环境的双重压力(王世杰,2002)。由于喀斯特生境岩石裸露率高、土壤浅薄、土被不连续、干湿频繁交替等特点,使喀斯特山区的植被恢复非常困难(朱守谦,2003)。因此,开展该森林植被对喀斯特生境的适应性研究,可为喀斯特生态脆弱区现存森林保护与严重退化区植被重建过程中树种的选择及合理配置提供科学依据(旷远文等,2010)。

在喀斯特适生植物的光合无机碳利用过程中,CA发挥着举足轻重的作用。CA是一种含锌金属酶,广泛存在于动物、植物、原核细菌与真核细菌中,参与各种生理过程,催化细胞中HCO3-向CO2和H2O的转化(Badger & Price,1994;Sasaki et al,1998)。不同抗旱植物CA活性在干旱逆境下有不同的灵敏度和柔韧性,CA活性越高,植物转化利用HCO3-的能力越强(Xing & Wu, 2012)。植物光合作用过程中同化利用的无机碳源一旦发生变化,其稳定碳同位素组成(δ13C)会相应发生变化。在光合作用一定的情况下,植物同化利用无机碳源的δ13C值越偏负,其叶片的δ13C值会相应地变得更为偏负,反之叶片的δ13C值就越偏正(Wu & Xing, 2012)。植物稳定碳同位素组成的变化已被成功地应用于光合作用的研究中,δ13C的测定可用于追踪光合作用过程中无机碳源的代谢途径(Motomura et al, 2008;Tcherkez et al, 2009)。
构树(Broussonetiapapyrifera)为荨麻目桑科落叶乔木,拥有较高的自然生长速率,相比于桑科中的其它物种,构树对各种不利环境具有更好的适应能力,喜光、耐干旱瘠薄,能生长于水边,多生于石灰岩山地,也能在酸性土及中性土上生长,广泛分布于中国南北各地(Wu & Xing, 2012;叶波等,2014),是喀斯特石漠化地区植被恢复的先锋树种之一。构树还有较强的HCO3-利用能力(吴沿友等,2011),得益于其较高的CA活性,在高重碳酸盐处理下的构树利用HCO3-的份额达30%(Wu & Xing, 2012)。朴树(Celtissinensis)则为荨麻目榆科落叶乔木,喜光照、微耐荫、喜温暖气候和粘质壤土,pH在4.5~7.5范围内生长良好,抗旱力强、也耐水湿和瘠薄、耐寒(任叔辉,2007)。朴树有很好的经济、药用与环保价值,广泛分布于江苏、安徽、山东、河南等省区,在以落叶阔叶林为主要植被类型的南京地区,朴树是主要的建群种之一(高邦权和张光富,2005)。光滑悬钩子(Rubustsangii)是蔷薇科悬钩子属的攀援灌木,喜光、耐旱,适应性强,多分布在400~1 500 m的向阳山坡,对土壤要求不严,pH值5.5~7.5均可生长(李志琴,2010)。悬钩子属植物具有很高的开发利用价值,是良好的水土保持及生态造林树种,全国各地均有分布(王浩波等,2009)。构树、朴树和光滑悬钩子都能在喀斯特地区正常生长,并能很好地适应该区的异质性环境。然而,朴树和光滑悬钩子在喀斯特异质性环境下的不同光合生理响应特性却仍未可知。
为了更深入了解喀斯特适生植物的不同适应性特征,并据此针对不同环境合理配置植被构成,结合喀斯特地区高pH、高重碳酸盐以及山区“天无三日晴”的阴湿气候等荫生环境特点,同时考虑到植物光合生理对高pH及重碳酸盐的响应特性可用于分析不同适生植物的不同无机碳利用特性,以及适生植物对不同喀斯特环境的适应能力强弱的比较,因此,本研究以构树、朴树和光滑悬钩子为材料,通过测定中坡位及下坡位两种不同环境下三种植物的CA活性、光合作用日变化、净光合速率对CO2浓度及光合有效辐射强度的响应曲线、叶绿素荧光特性以及稳定碳同位素组成等指标,探讨三种植物对喀斯特环境的光合生理响应特点,分析其不同适应性特征,为喀斯特生态脆弱地区适生植物的选择以及合理配置提供依据。
1 材料与方法
1.1 研究区概况
普定县位于贵州省中部安顺市西北部,105°27′49″~105°58′51″ E、26°9′36″~26°31′42″ N,东西长51.4 km,南北宽40 km,海拔1 100~1 600 m,属北亚热带季风湿润性气候,季风交替明显,气候温和,年均降雨量1 390 mm,年均气温在15.1 ℃左右。全县喀斯特地貌广泛发育,碳酸盐岩出露面积占全县国土面积的79.2%,土壤以石灰土和黄壤为主,分别占土壤总面积的63.7%和20.1%。普定县植被类型丰富多样,但由于自然和人为双重因素的影响,该县土壤侵蚀严重,裸露和半裸露石山连片出现,在多数地区植被退化严重(旷远文等,2010)。研究区的土壤为石灰土,微生物活动较强,易于有机质的降解(田丽艳等,2013)。采样点位于阳面的中(海拔1 300 m)、下(海拔1 190 m)坡位,两种坡位环境下的土壤均呈现弱碱性,HCO3-浓度在0.5 mmol·L-1左右。中坡位的光合有效辐射强度平均为202 μmol·m-2·s-1,最高达570 μmol·m-2·s-1;而下坡位平均为155 μmol·m-2·s-1,最高达350 μmol·m-2·s-1。两种坡位环境下的大气CO2浓度均为398 μmol·mol-1左右。中坡位的空气相对湿度平均为23.9%,而下坡位平均为31.1%。
1.2 材料
供试材料为处于遮荫环境下生长的成熟期的3~5年生的构树、朴树和光滑悬钩子的植物叶片。以两种不同坡位环境作为对比,并分别对应选取各自环境下生长的三种植物进行测定,每种环境下每种植物各随机选取5棵进行试验,每棵植物每个指标重复测定5次。将每种环境下每种植物根系周围重复3次采集的土样各自风干、研磨后按相同比例混匀,过0.25 mm筛后再分成3份,重复3次测定各自环境的理化性质,对比分析两种环境下土壤理化性质的差异。
1.3 方法
1.3.1 土壤pH和HCO3-浓度的测定土壤pH用电极法测定,水土比为5∶1;HCO3-浓度采用中和滴定法测定,各指标重复测定3次。
1.3.2 光合作用日变化用便携式光合测量系统Li-6400(LI-COR, Lincoln, NE, USA)测定三种植物第4片完全展开叶的净光合速率(Pn)、气孔导度(Gs)与蒸腾速率(E)的日变化,各指标重复测定5次。
水分利用效率通过方程式(1)计算获得:
WUE=Pn/E
(1)
式中,Pn是净光合速率,E是蒸腾速率。
1.3.3 净光合速率对CO2响应曲线的测定用便携式光合测量系统Li-6400(LI-COR, Lincoln, NE, USA)测定三种植物第4片完全展开叶的净光合速率对CO2的响应曲线,重复测定5次,据此拟合直角双曲线方程(2),再通过方程计算Amax、CE以及Rp:
(2)
式中,Ci为胞间CO2浓度(μmol·mol-1),Amax为CO2饱和时的净光合速率,Rp为叶片的光呼吸速率,羧化效率(CE)为曲线的起始斜率。用于测定CO2浓度梯度为400、300、200、100、50、100、200、300、400、600、800、 1 000、1 200、1 500、1 800 μmol·mol-1。叶室光合有效辐射与温度分别设定为1 000 μmol·m-2·s-1和18 ℃。
1.3.4 净光合速率对光响应曲线的测定用便携式光合测量系统Li-6400(LI-COR, Lincoln, NE, USA)测定三种植物第4片完全展开叶的净光合速率对光的响应曲线,重复测定5次,利用CO2控制系统将CO2浓度设定为400 μmol·mol-1,温度为18 ℃,利用人工光源控制叶室内的光合有效辐射,用于测定的光合有效辐射强度的梯度为2 000、1 750、1 500、1 250、1 000、800、600、400、200、150、120、100、80、60、40、20、0 μmol·m-2·s-1。采用非直角双曲线方程(3)对该响应曲线先进行拟合,再通过方程计算Pmax、Q以及Rd:
(3)
式中,Pmax为光饱和时的净光合速率,Rd为叶片的暗呼吸速率,表观量子效率(Q)为曲线的起始斜率,I为光合有效辐射强度,K为反映光响应曲线弯曲程度的曲角参数。

1.3.6 δ13C测定植物的第1片完全展开叶被用于δ13C的测定,重复测定5次。植物叶片经烘干、粉碎、过筛后,称取一定量转化成供质谱仪分析的CO2气体;在MAT-252质谱仪(Mat-252, Finnigan MAT, Germany)上进行测定,测量精度为小于±0.2‰。分析结果以δ13CPDB表示。
1.3.7 CA活性植物第4、第5片完全展开叶被用于CA活性测定,每种植物重复5次。称取植物叶片0.1~0.2 g,放到预冷的研钵中,先迅速加入液氮,再加入3 mL 10 μmol·L-1巴比妥钠缓冲液(含巯基乙醇50 μmol·L-1,pH 8.3)进行研磨,取研磨液倒入5 mL的离心管中,离心管置于冰浴中20 min后,在12 000 ×g离心5 min,取上清液,冷藏待用。
CA活性测定采用改进的pH计法(Wu et al, 2011)。保持反应系统在0~2 ℃,取待测上清液50~1 000 μL,加入到含4.5 mL的巴比妥钠缓冲液(20 μmol·L-1,pH8.3)的反应容器中,迅速加入3 mL预冷的(0~2 ℃)饱和CO2蒸馏水,用pH电极监测反应体系pH值变化,记下pH值下降一个单位(如pH值从8.2到7.2)所需的时间(记为t),同时记录在酶失活条件下pH值下降一个单位所需的时间(记为t0),酶的活力用WA(U·g-1FW)表示。
WA=t/t0-1
(4)
1.4 数据分析
所有数据采用SPSS 13.0统计软件及Excel软件进行差异显著性检验(LSD法)及做图。
2 结果与分析
2.1 三种植物光合参数的日变化
在中坡位,构树的Pn日变化呈现双峰趋势,两个峰值分别出现在12:00和15:00,分别为9.67和10.21 μmol·m-2·s-1,朴树的Pn则表现出随时间变化逐渐下降的趋势,9:00时的Pn最高,为5.55 μmol·m-2·s-1,光滑悬钩子的Pn日变化大致表现为单峰曲线,最高值出现在14:00,为6.71 μmol·m-2·s-1,构树、朴树和光滑悬钩子的日平均Pn分别为5.87、3.38与4.00 μmol·m-2·s-1(图1)。在下坡位,构树、朴树和光滑悬钩子的Pn日变化都表现为单峰曲线,构树和光滑悬钩子的最高值都出现在13:00,分别为8.44和5.34 μmol·m-2·s-1,朴树的最高值则出现在12:00,为5.64 μmol·m-2·s-1,构树、朴树和光滑悬钩子的日平均Pn分别为3.98、2.01与3.15 μmol·m-2·s-1。
在中坡位,构树的Gs日变化也呈现出双峰趋势,两个峰值分别出现在11:00和15:00,分别为0.08和0.05 mol·m-2·s-1,朴树的Gs在9:00~15:00间变化相对较为稳定,其值在0.03~0.04 mol·m-2·s-1之间波动,从16:00开始下降为0.01 mol·m-2·s-1,光滑悬钩子的Gs日变化表现为单峰曲线,峰值出现在11:00,为0.06 mol·m-2·s-1,构树、朴树和光滑悬钩子的日平均Gs分别为0.04、0.03与0.03 mol·m-2·s-1。在下坡位,构树的Gs则随时间变化而逐渐减小,9:00时的Gs最高,为0.06 mol·m-2·s-1,朴树和光滑悬钩子的Gs日变化则都表现为单峰曲线,最高值分别出现在12:00和11:00,分别为0.03和 0.05 mol·m-2·s-1。构树、朴树和光滑悬钩子的日平均Gs分别为0.03、0.02与0.03 mol·m-2·s-1。
三种植物在中坡位及下坡位两种环境下的WUE日变化均出现双峰趋势。在中坡位,构树的两个峰值分别出现在9:00和12:00,朴树出现在9:00和13:00,光滑悬钩子则出现在9:00和14:00,构树、朴树和光滑悬钩子的日平均WUE值分别为6.04、4.20与4.37 mmol·mol-1。在下坡位,构树的两个峰值分别出现在13:00和16:00,朴树出现在12:00和15:00,光滑悬钩子则出现在9:00和13:00,构树、朴树和光滑悬钩子的日平均WUE值分别为5.05、3.26与4.17 mmol·mol-1。

图 1 中坡位与下坡位三种植物的光合特性日变化A. 净光合速率; B. 气孔导度; C. 水分利用效率。Fig. 1 Diurnal changes of photosynthetic traits of three plant species on the middle slope and lower slopeA. Net photosynthetic rate (Pn); B. Stomatal conductance (Gs); C. Water-use efficiency (WUE).
2.2 三种植物光合作用对CO2浓度的响应
由表1可知,在下坡位生长的构树和光滑悬钩子的CE、Rp和Amax都显著高于中坡位,而朴树的CE、Rp和Amax则没有显著差异。在中坡位,光滑悬钩子的CE显著高于朴树和构树,后两者则不存在显著差异;构树的Rp则显著高于朴树和光滑悬钩子;而朴树和光滑悬钩子的Amax则显著高于构树。在下坡位,三种植物的CE、Rp和Amax大小均依次为光滑悬钩子>构树>朴树。
2.3 三种植物光合-光响应的特性
由表2可知,在中坡位,三种植物的Q大小依次为朴树>光滑悬钩子>构树,构树的Rd与光滑悬钩子没有显著差异,两者均小于朴树,朴树的Pmax与光滑悬钩子没有显著差异,两者均小于构树。在下坡位,三种植物的Q大小依次为朴树>构树>光滑悬钩子,光滑悬钩子的Rd与朴树无显著差异,两者均大于构树,而光滑悬钩子的Pmax与构树没有显著差异,两者大于朴树。构树在下坡位的Q和Rd分别大于其各自在中坡位的Q和Rd,两种环境下生长的构树的Pmax则没有显著差异,朴树在下坡位的Pmax以及光滑悬钩子在下坡位的Q分别小于其各自在中坡位的值,光滑悬钩子在下坡位的Rd和Pmax则均大于其各自在中坡位的值。

表 1 三种植物净光合速率对CO2浓度响应曲线的特征参数
注:平均值±标准误差(n=5)后面字母表示在同一显著水平(P≤0.05),通过单因素方差分析与t检验对同一列数据进行差异显著性分析。下同。
Note: Mean ± SE(n=5) followed by different letters in the same row differ significantly (P≤0.05), according to one-way ANOVA and t test. The same below.

表 2 三种植物净光合速率对光合有效辐射响应曲线的特征参数
2.4 三种植物叶绿素荧光参数的比较
由表3可知,中坡位的朴树拥有最高的Fo值,下坡位的构树则拥有最高的Fv/Fm与Fv/Fo值。在中坡位,ΦPSⅡ值大小均依次为构树>朴树>光滑悬钩子。在下坡位,ΦPSⅡ值大小均依次为构树>光滑悬钩子>朴树。构树在下坡位的Fo值小于中坡位,其余荧光参数值则没有显著差异。朴树在下坡位的Fo和ΦPSⅡ值均小于中坡位。光滑悬钩子在下坡位的ΦPSⅡ值显著高于中坡位。
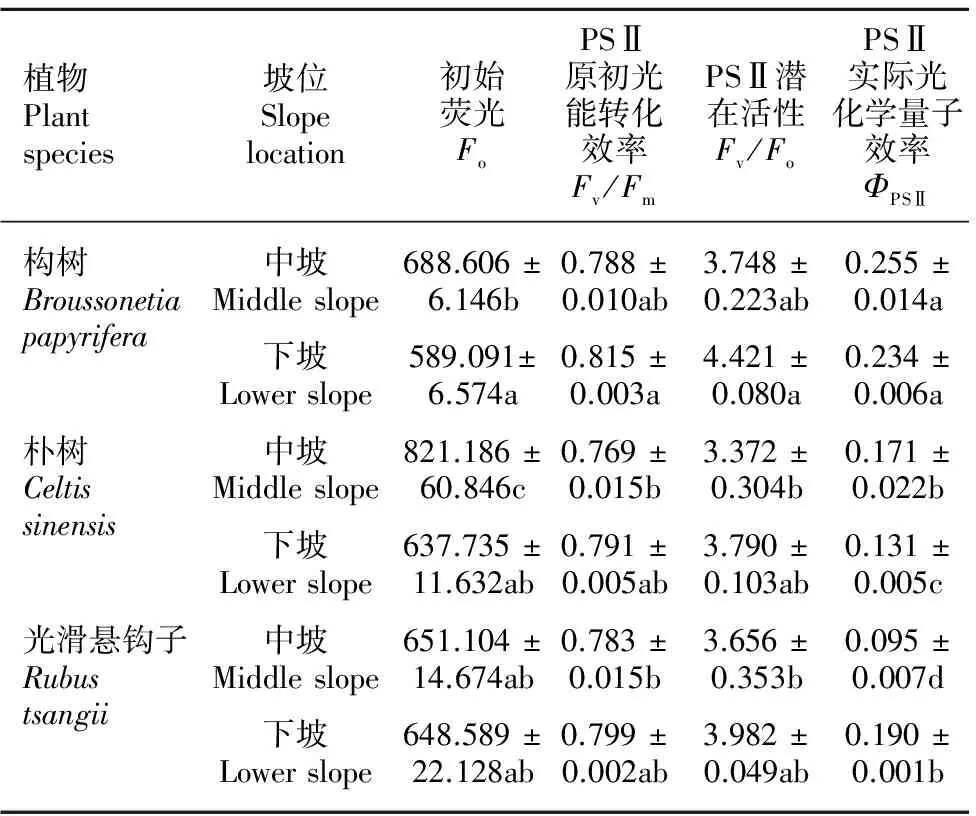
表 3 三种植物的初始荧光、PSⅡ原初光能转化效率、PSⅡ潜在活性与PSⅡ实际光化学量子效率
2.5 三种植物叶片的δ13C和CA活性
朴树的δ13C值最高,整体上较为偏正,且中坡位与下坡位生长的朴树叶片的δ13C值之间无显著差异(表4)。相对于朴树来说,构树叶片的δ13C值较为偏负,下坡位生长的光滑悬钩子叶片的δ13C值最低,中坡位生长的构树和光滑悬钩子叶片的δ13C值都比下坡位生长的构树和光滑悬钩子叶片的δ13C值要高。构树叶片的CA活性显著高于朴树和光滑悬钩子,而下坡位生长的构树叶片的CA活性又显著高于中坡位。朴树和光滑悬钩子叶片的CA活性均很难被检测到。

表 4 三种植物叶片的稳定碳同位素组成和碳酸酐酶活性
3 讨论
在喀斯特地区生长的植物因为经常遭受各种各样的逆境而造成Gs的减小或关闭,气孔阻力增加,使得大气中的CO2通过自由扩散的方式进入植物细胞间隙变得更为困难。在该地区干旱或者高重碳酸盐等逆境胁迫下,受气孔部分闭合的影响,进入植物叶片细胞的CO2浓度降低,导致细胞中的激活而升高。本研究中,三种植物的气孔均出现不同HCO3-向CO2和H2O的转化,经此诱导,CA活性将被程度部分闭合现象,然而作为喀斯特适生植物,因为构树本身拥有较高的CA活性,且容易在喀斯特逆境(干旱或高重碳酸盐等逆境胁迫)下被激活,高效催化转化细胞内的HCO3-为H2O和CO2,弥补因为Gs减小或关闭而造成的碳源和水源的不足,维持构树光合作用的正常进行(Wu & Xing, 2012),而朴树和光滑悬钩子因为活性较难被检测到,并因此被视为没有CA活性,无法被激活。所以构树表现出比朴树和光滑悬钩子更强的光合效率,朴树受气孔限制最明显,其光合效率也最低。中坡位的日均光合有效辐射强度高于下坡位,考虑到三种植物均具有喜光性,所以三种植物在中坡位的日均光合效率都较高。下坡位生长的构树比中坡位生长的构树拥有更高的CA活性,其催化转化细胞内的HCO3-为H2O和CO2的效率也更高,更能弥补中午因环境变化引起Gs减小而导致的水分不足,所以与中坡位生长的构树不同的是,下坡位生长的构树没有出现“光合午休”现象可能与此有关。朴树则属于慢生树种,光合效率也较低,所以朴树对CO2或者H2O的需求相对较低,两种环境下朴树均没有出现“光合午休”现象,说明中午环境的变化没有造成朴树光合作用过程中H2O和CO2的不足。此外,与下坡位相比,中坡位的构树和朴树拥有平均较高的水分利用效率,那么中坡位的构树和朴树在同样单位水分损失的情况下能够获取固定更多的无机碳。光滑悬钩子的枝叶生有皮刺,在干旱环境下具有减小水分散失的功能,其Gs不会随中坡位空气相对湿度的减小而降低,并能保持较为稳定的WUE。
CE可以反映叶片对进入细胞间隙CO2的同化状况,CE越高,说明光合作用对CO2的利用就越充分(叶子飘和于强,2008)。下坡位的构树和光滑悬钩子在气孔导度较小,无机碳源受限的情况下,都能够高效利用从大气进入细胞间隙的CO2或者是通过自身呼吸而产生的CO2。光呼吸产生的CO2可被光合作用回收利用(Loreto et al, 2001)。下坡位生长的光滑悬钩子通过光呼吸而为光合作用提供CO2的能力比中坡位高,同时也高于构树和朴树。这可能与中坡位光滑悬钩子增大的Gs有关,气孔打开有利于大气中CO2进入细胞间隙,光呼吸就可能会受到抑制。叶片暗呼吸是植物分解有机物并释放能量的过程(张富华等,2014),构树和光滑悬钩子在中坡位的Rd均低于各自在下坡位的值,可能与中坡位拥有较高的日均光合有效辐射强度有关,增强的光合有效辐射强度对暗呼吸产生了一定程度上的抑制。朴树拥有平均较高的Rd,不利于有机物的累积和生物量的增加。Q则可以反映植物在遮荫环境下对光能的利用效率(叶子飘和于强,2008),遮荫环境对朴树光能利用效率的影响不大,构树则在低光强下体现出较高的光能利用效率。
PSⅡ反应中心的活性能够通过Fo的值来反映,Fo值的增加说明PSⅡ反应中心活性降低(孙晓方等,2008),本研究结果表明,与下坡位相比,中坡位的构树和朴树的PSⅡ反应中心发生一定程度的失活,而三种植物Fv/Fm、Fv/Fo值的变化各自都不显著,说明两种环境下三种植物的光合结构都未受损伤。而中坡位的朴树和下坡位的光滑悬钩子较高的ΦPSⅡ值则有利于提高光能的转化效率,为暗反应的光合同化积累更多所需的能量,以促进碳同化的高效运转和有机物的积累(罗青红等,2006)。
WUE的增加以及海拔的升高都会引起δ13C值的增加(Farquhar & Richards,1984;杨成等,2007),然而中坡位构树的δ13C值并未显著高于下坡位。据分析,下坡位的构树具有显著较高的CA活性,这有利于构树在气孔部分闭合碳源受限的情况下更加高效地利用细胞内的HCO3-,而喀斯特地区土壤中的HCO3-的δ13C值比大气CO2的δ13C值更为偏正(Clark & Fritz, 1997),所以对HCO3-的高效利用使得下坡位构树的δ13C值并没有变的明显偏负。朴树的光能利用效率及光合能力不受坡位环境变化的影响,且其光合作用过程中的无机碳源只能来自大气中的CO2,海拔的升高和WUE的增加并未引起朴树δ13C值的变化,这可能与其稳定的CO2固定及同化效率有关,且作为慢生树种,朴树在光合作用的过程中发生的稳定碳同位素分馏效应也最小。光滑悬钩子在中坡位的δ13C值比下坡位显著偏正,在WUE较为稳定的情况下,主要是受海拔升高的影响,那么下坡位过于偏负的δ13C值可能还受其它无机碳源的影响。据研究,CO2释放点靠近叶绿体内侧,且没有边界层、气孔以及细胞壁的限制,因此,光呼吸产生的CO2极易被光合作用重新利用(Loreto et al, 2001),但经过光合作用过程中同位素的分馏作用,稳定碳同位素组成将变得比大气CO2的δ13C值更为偏负,如果光呼吸产生的CO2被光滑悬钩子的光合过程重新利用后,其叶片的δ13C值就将变得更为偏负,关于这方面的推测有待进一步研究证实。
4 结论
三种植物对不同的喀斯特坡位环境具有不同的适应特点及无机碳利用特性。构树光合作用过程中所利用的无机碳源既可以来自大气中的CO2,亦可以在气孔部分闭合的情况下利用细胞内的HCO3-,下坡位的构树较高的CA活性使得其利用HCO3-的效率也会更高,并能在光强相对较低的情况下具有较高的光能利用效率,这与喀斯特适生植物最重要的一个适应性特征-较高的CA活性相一致。构树对下坡位具有相对较好的适应性。朴树缓慢的生长速度以及无机碳同化利用效率,使朴树对无机碳源的需求最低,且能保持较稳定的光合效率,但朴树光合作用过程中的无机碳来源较单一,主要利用大气中的CO2。中坡位的朴树具有相对较高的净光合速率和光能利用效率,所以朴树较适合生长在中坡位环境。光滑悬钩子主要利用大气中的CO2进行光合作用。下坡位的光滑悬钩子拥有较高的呼吸速率,能更加有效地分解有机物并释放能量。但中坡位的光滑悬钩子表现出较高的净光合速率和光能利用效率。因为光滑悬钩子的枝叶具有皮刺,能有效保持水分,所以中坡位相对较低的空气相对湿度也不会影响光滑悬钩子的PSⅡ的活性、气孔开度以及WUE。同时考虑到两种环境下的大气CO2浓度无显著差别,而中坡位的光合有效辐射强度显著较高,所以选择中坡位作为光滑悬钩子的适宜种植环境。
BADGER MR, PRICE GD, 1994. The role of carbonic anhydrase in photosynthesis [J]. Ann Rev Plant Physiol, 45:369-392.
BAI ZG,WAN GJ,1998. Study on watershed erosion rate and its environmental effects in Guizhou karst region [J]. J Soil Eros Soil Water Conserv,4(1):1-7. [白占国,万国江,1998. 贵州碳酸盐岩区域的侵蚀速率及环境效应研究 [J]. 土壤侵蚀与水土保持学报,4(1):1-7.]
CLARK ID, FRITZ P, 1997. Environmental Isotopes in Hydrogeology [M]. New York: CRC Press: 152.
FARQUHAR GD, RICHARDS RA, 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes [J]. Aust J Plant Physiol, 11(6):539-552.
GALLE A, FLOREZ-SARASA I, THAMEUR A, et al, 2010. Effects of drought stress and subsequent rewatering on photosynthetic and respiratory pathways in Nicotiana sylvestris wild type and the mitochondrial complex l-deficient CMSII mutant [J]. J Exp Bot, 61(3):765-775.
GAO BQ,ZHANG GF,2005. Studies on population structure and distribution pattern ofCeltissinensisin Laoshan National Forest Park of Nanjing [J]. Guihaia,25(5):406-412. [高邦权,张光富,2005. 南京老山国家森林公园朴树种群结构与分布格局研究 [J]. 广西植物,25(5):406-412.]
HU HH, BOISSON-DERNIER A, ISRAELSSON-NORDSTRÖM M, et al, 2011. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells [J]. Nat Cell Biol, 13(6):734-734.
KUANG YW,WEN DZ,YAN JH,et al, 2010. Characteristics of element contents in leaves of 3 dominant species in karst forest in Puding, Guizhou, China [J]. Chin J Appl Environ Biol,16(2):159-163. [旷远文,温达志,闫俊华,等,2010. 贵州普定喀斯特森林三种优势树种叶片元素含量特征 [J]. 应用与环境生物学报,16(2):159-163.]
LI ZQ,2010. Development and protection of Rubus wild plants in Guizhou [J]. Agr Technol Serv,27(1):100. [李志琴,2010. 悬钩子属野生植物在贵州的开发与保护 [J]. 农技服务,27(1):100.]
LORETO F, VELIKOVA V, MARCO GD, 2001. Respiration in the light measured by12CO2emission in13CO2atmosphere in maize leaves [J]. Aust J Plant Physiol, 28(11):1103-1108.
LUO QH,LI ZJ,WU WM,et al, 2006. Comparative study of photosynthetic and chlorophyll fluorescence characteristics ofPopulouseuphraticaandP.pruinosa[J]. Acta Bot Boreal-Occident Sin,26(5):983-988. [罗青红,李志军,伍维模,等,2006. 胡杨、灰叶胡杨光合及叶绿素荧光特性的比较研究 [J]. 西北植物学报,26(5):983-988.]
MOONEY HA, 1972. The carbon balance of plants [J]. Ann Rev Ecol Syst, 3:315-346.
MOTOMURA H, UENO O, KAGAWA A, et al, 2008. Carbon isotope ratios and the variation in the diurnal pattern of malate accumulation in aerial roots of CAM species ofPhalaenopsis(Orchidaceae) [J]. Photosynthetica, 46(4):531-536.
PAN RC,WANG XJ,LI NH,2012. Plant physiology [M]. Beijing:Higher Education Press:112-113. [潘瑞炽,王小菁,李娘辉,2012. 植物生理学 [M]. 北京:高等教育出版社:112-113.]
REN SH,2007. Applied research ofCeltissinensisin the construction of ecological landscape architecture [J]. Prot For Sci Technol,(4):108-109. [任叔辉,2007. 朴树在生态园林建设中的应用研究 [J]. 防护林科技,(4):108-109. ]

SASAKI H, HIROSE T, WATANABE Y, et al, 1998. Carbonic anhydrase activity and CO2-transfer resistance in Zn-dificient rice leaves [J]. Plant Physiol, 118(3):929-934.
SUN XF,HE JQ,HUANG XD,et al, 2008. Growth characters and chlorophyll fluorescence of goldenrod (Solidagocanadensis) in different light intensities [J]. Acta Bot Boreal-Occident Sin,28(4):752-758. [孙晓方,何家庆,黄训端,等,2008. 不同光强对加拿大一枝黄花生长和叶绿素荧光的影响 [J]. 西北植物学报,28(4):752-758.]
TCHERKEZ G, MAHÉ A, GAUTHIER P, et al,2009. In folio respiratory fluxomics revealed by13C isotopic labeling and H/D isotope effects highlight the noncyclic nature of the tricarboxylic acid “cycle” in illuminated leaves [J]. Plant Physiol, 151(2):620-630.
TIAN LY,LANG YC,LIU CQ,et al, 2013. Distribution patterns of organic carbon and its isotope compositions in soil profiles on the slopes in Puding karst areas of Guizhou Province, Southwest China [J]. Chin J Ecol,32(9):2362-2367. [田丽艳,郎赟超,刘丛强,等,2013. 贵州普定喀斯特坡地土壤剖面有机碳及其同位素组成 [J]. 生态学杂志,32(9):2362-2367.]WALTERS MB, KPUGER EL, REICH PB, 1993. Relative growth rate in relation to physiological and morphological traits for northern hardwood tree seedlings: species, light environment and ontogenetic considerations [J]. Oecologia, 96(2):219-231.
WANG HB,LI JH,YANG TL,et al, 2009. The geographical patterns of species diversity ofRubusin Yunnan Province [J]. J Dali Univ,8(8):63-66. [王浩波,李继红,杨堂亮,等,2009. 云南省悬钩子属植物多样性的地理分布格局 [J]. 大理学院学报,8(8):63-66.]
WANG SJ,2002. Concept deduction and its connotation of karst rocky desertification [J]. Carsol Sin,21(2):101-105. [王世杰,2002. 喀斯特石漠化概念演绎及其科学内涵的探讨 [J]. 中国岩溶,21(2):101-105.]WU YY, SHI QQ, WANG K, et al.,2011. An electrochemical approach coupled with Sb microelectrode to determine the activities of carbonic anhydrase in the plant leaves [J]//ZENG D. Future intelligent information systems. LNEE 86: Berlin: Springer: 87-94.
WU YY, XING DK,2012. Effect of bicarbonate treatment on photosynthetic assimilation of inorganic carbon in two plant species of Moraceae [J]. Photosynthetica, 50(4):587-594.
WU YY,XING DK,LIU Y,2011. The characteristics of bicarbonate used by plants [J]. Earth Environ,39(2):273-277. [吴沿友,邢德科,刘莹,2011. 植物利用碳酸氢根离子的特征分析 [J]. 地球与环境,39(2):273-277.]
XING DK, WU YY,2012. Photosynthetic response of three climber plant species to osmotic stress induced by polyethylene glycol (PEG) 6000 [J]. Acta Physiol Plant, 34(5):1659-1668.
YANG C,LIU CQ,SONG ZL,et al, 2007. Characteristics of carbon isotopic compositions in karst mountain plants and their indication to water use efficiency——A case study on Yangzhong catchment in Huaxi, Guizhou [J]. Carsol Sin,26(2):105-120. [杨成,刘丛强,宋照亮,等,2007. 喀斯特山区植物碳同位素组成特征及其对水分利用效率的指示——以贵州花溪杨中小流域为例 [J]. 中国岩溶,26(2):105-120.]
YE B,WU YB,SHAO W,et al, 2014. Effects of combined stress of elevated temperature and drought and of re-watering on the photosynthetic characteristics and chlorophyll fluorescence parameters ofBroussonetiapapyriferaseedlings [J]. Chin J Ecol,33(9):2343-2349. [叶波,吴永波,邵维,等,2014.高温干旱复合胁迫及复水对构树(Broussonetiapapyrifera)幼苗光合特性和叶绿素荧光参数的影响 [J]. 生态学杂志,33(9):2343-2349.]
YE ZP,GAO J,2008. Change of carboxylation efficiency of Salvia miltiorrhiza in the vicinity of CO2compensation point [J]. J Northwest A & F Univ: Nat Sci Ed,36(5):160-164. [叶子飘,高峻,2008. 丹参羧化效率在其CO2补偿点附近的变化 [J]. 西北农林科技大学学报·自然科学版,36(5):160-164.]
YE ZP,YU Q,2008. Comparison of new and several classical models of photosynthesis in response to irradiance [J]. J Plant Ecol,32(6):1356-1361. [叶子飘,于强,2008. 光合作用光响应模型的比较 [J]. 植物生态学报,32(6):1356-1361.]
ZHANG CS,LIU GB,XUE S,et al, 2012. Photosynthetic characteristics ofBothriochloaischaemumunder drought stress and elevated CO2concentration [J]. Chin J Appl Ecol,23(11):3009-3015. [张昌胜,刘国彬,薛萐,等,2012. 干旱胁迫和CO2浓度升高条件下白羊草的光合特征 [J]. 应用生态学报,23(11):3009-3015.]
ZHANG FH,HU D,SUN F,et al, 2014. Ecological effects of short-time increased air temperature in Summer and Autumn periods on dark respirations and typical photosynthetic characteristics forRosachinensisin Beijing urban areas [J]. Acta Ecol Sin,34(24):7385-7392. [张富华,胡聃,孙凡,等,2014. 北京城区夏、秋季节短期增温对月季暗呼吸及光合参数的影响 [J]. 生态学报,34(24):7385-7392.]
ZHU SQ,2003. Karst forest ecosystem research Ⅲ [M]. Guiyang:Guizhou Science and Technology Press:72. [朱守谦,2003. 喀斯特森林生态系统研究Ⅲ [M]. 贵阳:贵州科技出版社:72.]
Photosynthetic physiological response of three plant species to different slope environments in karst forest of Guizhou, China
XING De-Ke1, WU Yan-You2*, FU Wei-Guo1, HANG Hong-Tao2,ZHAO Yu-Guo1, WU Yan-Sheng1
( 1. Key Laboratory of Modern Agricultural Equipment and Technology, Ministry of Education, Institute of Agricultural Engineering, JiangsuUniversity, Zhenjiang 212013, Jiangsu, China;2.ResearchcenterforEnvironmentalBio-ScienceandTechnology,StateKeyLaboratoryofEnvironmentalGeochemistry,InstituteofGeochemistry,ChineseAcademyofSciences, Guiyang 550002, China )
In order to gain a better understanding on the different adaptive characteristics of karst plants and to develop reasonable vegetation we studied the carbonic anhydrase (CA) activity, diurnal variation of photosynthesis, response curves of net photosynthetic rate of CO2concentration and light intensity, chlorophyllafluorescence parameters and stable carbon isotope composition ofBroussonetiapapyrifera,CeltissinensisandRubustsangiiwhich grew in the middle and lower slopes of the karst forest in Puding County, Guizhou Province. Their different photosynthetic physiological response of the three species were also analyzed. The results indicated that in the photosynthesis,B.papyriferacould utilize the atmospheric CO2and the intracellular HCO3-through the catalysis conversion by CA when the stomatal conductance decreased. With higher activity of CA,B.papyriferagrowing in the lower slope had higher HCO3-utilization efficiency. Besides,B.papyriferagrowing in the lower slope exhibited higher light utilization efficiency under low light intensity, which might be attributed to its higher CA activity in the lower slope.B.papyriferashowed better adaptability to lower slope compared to middle slope. The photosynthetic inorganic carbon assimilation efficiency ofC.sinensiswas the lowest, which only used the atmospheric CO2as inorganic carbon resource for the photosynthesis. Since the growth rate ofC.sinensiswas very slow, the requirement for inorganic carbon was the lowest, and it could maintain the stable inorganic carbon assimilation rate.C.sinensisgrowing in the middle slope exhibited higher net photosynthetic rate and light utilization efficiency, and it showed better adaptability to the middle slope compared to lower slope.R.tsangiimainly utilized the atmospheric CO2in the photosynthesis process. The light utilization efficiency and net photosynthetic rate ofR.tsangiiin the middle slope were higher.R.tsangiiexhibited better adaptability to middle slope. The results provide a scienti-fic basis for reasonably selecting and designing plant species during the revegetation process in the karst ecologically vulnerable forest region.
revegetation, carbonic anhydrase, apparent quantum efficiency, carboxylation efficiency, composition of stable carbon isotope
10.11931/guihaia.gxzw201507030
2015-11-27
2015-12-28
国家自然科学基金青年基金(31301243);贵州省社会发展攻关项目(黔科合SY [2010]3043);江苏大学引进人才科研启动基金(13JDG030)[Supported by the National Natural Science Foundation of China (31301243); the Brainstorm Project on Social Development of Guizhou Province (SY [2010]3043); the Research Foundation for Introduce Talents of Jiangsu University (13JDG030)]。
邢德科(1983-),男,河南南阳人,博士,助理研究员,主要从事植物逆境生理生态方面的研究,(E-mail) xingdeke@ujs.edu.cn。
吴沿友,博士,研究员,研究方向为环境地球化学,(E-mail) wuyanyou@mail.gyig.ac.cn。
Q945.1, S945.79
A
1000-3142(2016)10-1147-09
邢德科,吴沿友,付为国,等. 贵州喀斯特森林三种植物对不同坡位环境的光合生理响应 [J]. 广西植物,2016,36(10):1147-1155
XING DK, WU YY, FU WG,et al. Photosynthetic physiological response of three plant species to different slope environments in karst forest of Guizhou, China [J]. Guihaia,2016,36(10):1147-1155
猜你喜欢
中南林业科技大学学报(2020年4期)2020-04-26 13:15:56
做人与处世(2020年8期)2020-04-26 01:38:37
花卉(2020年4期)2020-03-16 08:20:26
绿色中国(2019年16期)2019-11-26 07:12:36
读者(2019年10期)2019-05-08 01:51:40
作文与考试·高中版(2019年14期)2019-04-26 12:37:26
中国林业产业(2019年3期)2019-03-25 02:42:30
江西农业(2018年15期)2018-11-23 08:02:12
海峡姐妹(2017年12期)2018-01-31 02:12:24
中国扶贫(2017年16期)2017-08-16 11:09:57
