
河南省同名小麦地方品种SSR遗传多样性分析
2016-11-08 11:43李正玲王会伟董海滨李春鑫许为钢
麦类作物学报 2016年5期
李正玲,胡 琳,王会伟,董海滨,李 艳,李春鑫,许为钢
(河南省农业科学院小麦研究中心/小麦国家工程实验室/农业部黄淮中部小麦生物学与遗传育种重点实验室/河南省小麦生物学重点实验室,河南郑州 450002)
河南省同名小麦地方品种SSR遗传多样性分析
李正玲,胡 琳,王会伟,董海滨,李 艳,李春鑫,许为钢
(河南省农业科学院小麦研究中心/小麦国家工程实验室/农业部黄淮中部小麦生物学与遗传育种重点实验室/河南省小麦生物学重点实验室,河南郑州 450002)
为了给河南省小麦地方品种的开发利用提供依据,利用SSR标记对白和尚头、白麦、白芒糙、出山豹等15组名称相同的小麦地方品种共计155份材料进行了组间和组内遗传多样性分析。结果发现,同名品种组内,15组材料的等位变异变化范围为47~79个,平均遗传相似系数变化范围为0.66~0.93,多态性信息含量分布范围为0.856~0.936;同名品种组间,155份材料共产生143个等位变异,遗传相似系数分布范围为0.75~1.00,平均遗传相似系数为0.90,多态性信息含量为0.981。分别对同名品种组内和组间进行聚类分析,结果发现,同名品种组内3组共7份材料遗传相似系数为1.00,而同名品种组间未出现遗传相似系数为1.00的材料。由此可见,河南省小麦地方品种具有丰富的遗传多样性,同名小麦地方品种间存在同名同质和同名异质的现象。
小麦地方品种; SSR;亲缘关系;遗传多样性
小麦是重要的粮食作物,在长期的进化和栽培过程中形成了丰富的遗传变异类型。由于历史上长期的种植,小麦地方品种在生态适应性、抗病性、抗逆性和品质特性等方面更是形成许多优异的基因资源。抗病性方面,河南地方品种齿牙糙携带的 Pm24基因对白粉病具有很强的抗性[1],老芒麦对条锈病具有全生育期抗性[2],长江中下游地区的地方品种望水白是全世界优异的赤霉病抗源[3];抗逆性方面,秃头麦具有抗穗发芽的突出优点[4];在品质方面,人们从地方品种中筛选出的Wx基因缺失种质在改良淀粉特性方面具有特殊的作用[5]。目前,许多研究者对小麦地方品种进行了大量的遗传多样性分析,并一致认为小麦地方品种具有较高的遗传多样性[6-8]。在目前具有突破性的育种材料日趋匮乏、品种遗传基础日益狭窄[9]的情况下,小麦地方品种所具有的遗传多样性及优异基因资源越来越受到广泛重视。
河南省是我国重要的小麦主产区,有着悠久的小麦栽培历史。经过长期广泛的搜集整理,河南省小麦种质资源库现保存有1 048份小麦地方品种。笔者在对河南省小麦种质资源库保存的地方品种资源进行整理与研究工作中,发现许多地方品种是由多种基因型组成的混合群体,且存在同名异质、同质异名的现象。张玲丽等[10]对5份来自不同种植地区的地方品种大青芒进行了材料内和材料间遗传多样性分析,发现在蛋白质和分子水平材料内和材料间都存在变异。这种现象不利于小麦地方品种的开发、利用。本研究是在前期的研究基础上[11],采用SSR分子标记对15组同名的共计155份河南省地方品种遗传多样性进行进一步分析,旨在为河南省小麦地方品种资源的开发利用提供科学依据。
1 材料与方法
1.1材 料
供试材料为155份河南省小麦地方品种,名称相同的小麦品种被分成1组,共15组(表1),均来自河南省农业科学院小麦研究中心资源库。
1.2DNA 提取
利用水培法培养黄化苗,每份材料15粒种子,随机选取新鲜嫩叶混合提取DNA,具体方法参照柴建芳等[12]。
1.3SSR分析
选取分布于不同染色体上的200对SSR引物进行筛选。PCR反应体系为10 μL,其中包括:ddH2O 8.15 μL,10×PCR Buffer 1 μL,dNTPs (5 mmol·L-1)0.2 μL,引物(20 μmol·L-1)0.2 μL,Taq酶(5 U·μL-1)0.05 μL,模板DNA (50 ng·μL-1)0.4 μL。反应在effendorf PCR仪上进行。反应程序为:94 ℃预变性 5 min;94 ℃变性30 s,55 ℃或60 ℃复性 30 s,72 ℃延伸45 s,40个循环,72 ℃延伸10 min。扩增产物用8%聚丙烯酰胺凝胶电泳分离、银染,照相,统计。
1.4数据分析
SSR 扩增带型在相同迁移率位置上有带记为“1”,无带记为“0”。 按Botstein等[13]的方法计算SSR位点和品种的多态性信息含量(Polymorphism information content,PIC):PICi=1-∑(Pi)2,其中Pi为某个SSR位点的第i个等位基因出现的次数占该位点全部等位变异出现次数的百分比。等位变异分布频率:含有该位点的品种数与所有品种的百分比。
按照Nei等[14]的方法计算材料间遗传相似系数:GS=2Nij/(Ni+Nj) ,其中Nij为第i、j材料共有的扩增片段数目,Ni为第i材料中出现的扩增片段数目,Nj为第j材料中出现的扩增片段数目。采用NTSYS-pc2.1软件系统进行数据处理,计算材料间的遗传相似系数,并根据GS值按UPGMA法对供试材料间进行聚类分析。
2 结果与分析
2.1引物筛选及多态性分析
利用材料白麦对200对引物进行筛选,结果得到51对带型清晰、多态性高且重复性好的引物,最后确定17对用于多样性研究,这17对引物分布在7个同源群上。17对引物在155份材料中共产生143个等位变异,等位变异在17对引物上的分布变化范围为2~27个,平均每对引物检测到8.4个。PIC值以Xgwm 344的最高(0.900),Xgwm 428的最低(0.124),其余引物的PIC值分布在这两者之间,平均为0.616(表2)。等位变异分布频率变化范围为0.6%~98.1%,平均为20.0%,其中分布频率为0.6%的等位变异占16.1%,分布频率低于5%的等位变异占45.5%。
*为遗传相似系数为1.00的材料
2.215组同名地方品种的遗传多样性
对15组同名小麦地方品种进行遗传多样性分析,结果(表3)表明,15组同名品种的平均遗传相似系数变化范围为0.66~0.93之间,多态性信息含量变化范围为0.856~0.936。白和尚头、白麦、出山豹、黄瓜先和三月黄组内存在着相似基因型(遗传相似系数>0.95),而大多数具有相同名称的品种实际为不同的基因型(品种间的遗传相似系数≤0.95),这说明同名地方品种内存在同名异质现象。
15组同名地方品种根据遗传相似系数和平均遗传相似系数可以分为四类:第一类遗传相似系数变化范围小,平均遗传相似系数大,有白和尚头、白麦、白芒糙;第二类遗传相似系数变化范围大,平均遗传相似系数小,且遗传相似系数最高为1,这类包含出山豹、三月黄;第三类遗传相似系数变化范围大,平均遗传相似系数小,这类包含二芒麦、黄瓜先、五花头、蚰子头;第四类遗传相似系数变化范围小,平均遗传相似系数小,这类包含和尚头、红和尚头、红芒白、红芒糙、红芒红、红蚰子头。
利用遗传相似系数对15组同名品种组内材料分别进行聚类分析,结果发现白和尚头组1号(洛阳)、2号(新安)、4号(郏县)3份材料,出山豹组35号(嵩县)和36号(嵩县)2份材料,三月黄组120号(温县)和121号(孟县)2份材料出现遗传相似系数为1.00的情况,这说明同名品种组内存在同质品种。
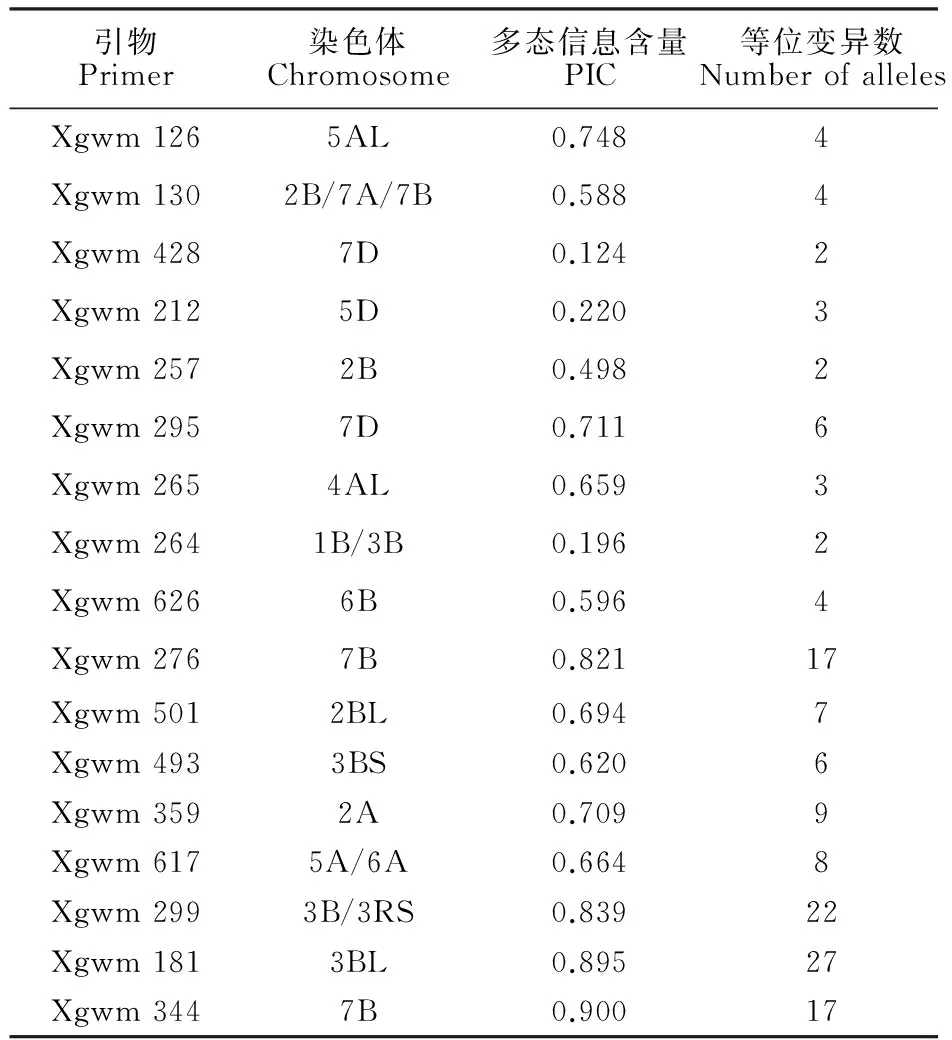
表2 17对SSR引物的染色体位置、PIC及等位变异数Table 2 Chromosome, PIC and polymorphic locus of seventeen SSR primers
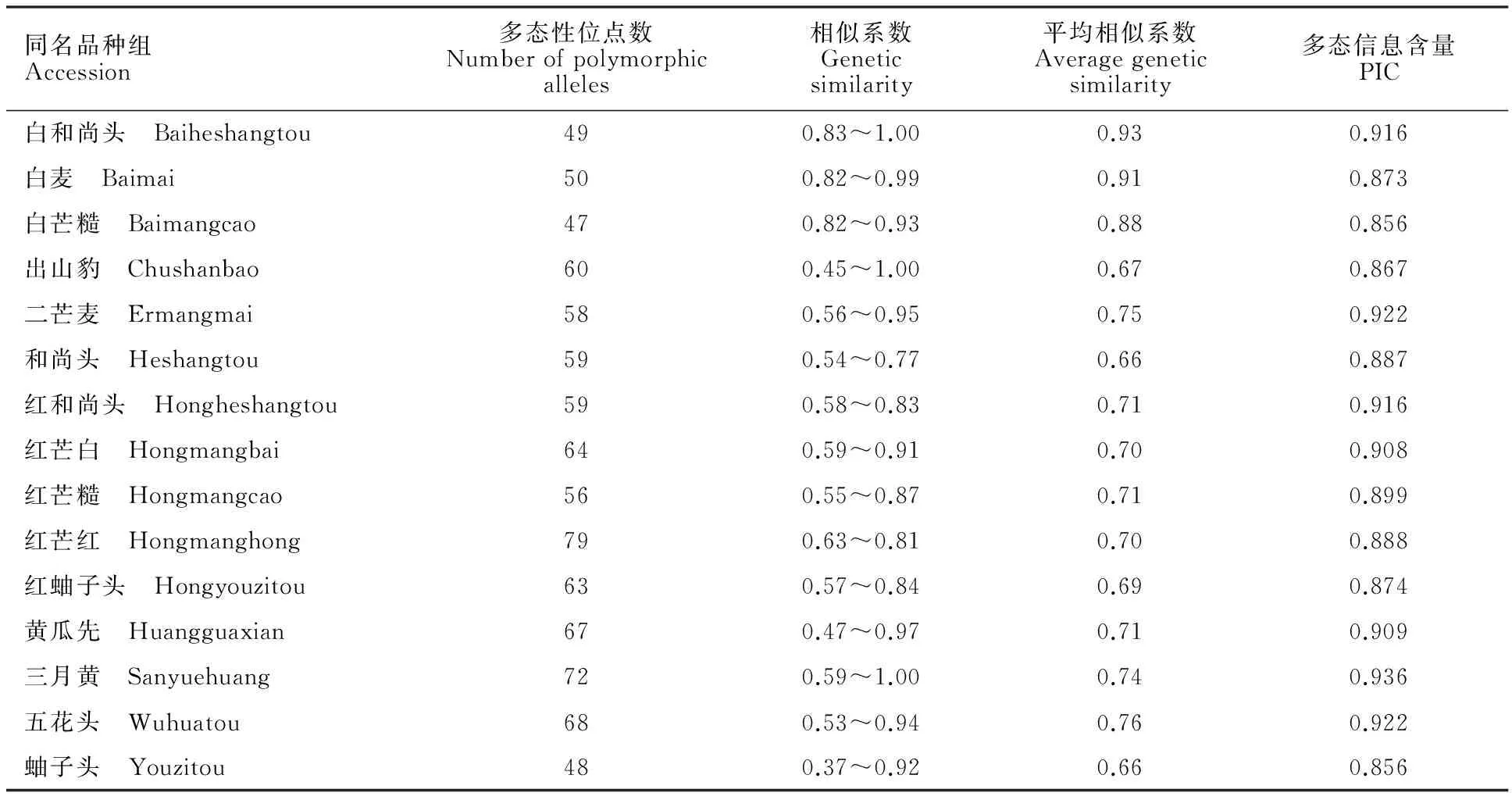
表3 15组地方品种扩增结果遗传多样性分析Table 3 Diversity analysis of fifteen species with SSR results
2.3155份参试材料的遗传多样性
155份材料共产生143条多态性条带。遗传相似系数分布范围为0.75~1.00,平均遗传相似系数为0.90,多态性信息含量为0.981。
对155份材料进行聚类分析,结果未发现不同组间遗传相似系数为1.00的材料(图1)。在遗传相似系数为0.805处 ,将155份材料分为五类:第一类包括2个材料,分别为五花头(144)、黄瓜先(111);第二类包括3个材料,分别为三月黄(130)、红芒红(99)、红芒白(78);第三类包括24个材料,分别为五花头(141、142、146、139、148、145),三月黄(126、128),黄瓜先(118),红蚰子(104),红芒红(92、93、96),红芒糙(82、85、89、91),红芒白(79),红和尚头(69),和尚头(51),出山豹(32),白芒糙(23、24、28);第四类包括29个材料,分别为五花头(138),三月黄(123),黄瓜先(116),红芒红(97),红芒白(72、74),红和尚头(62、66),和尚头(52、58),二芒麦(39、40、41、42、43、44、45、47、49),出山豹(34、35、36),白麦(13、17、19、21),白和尚头(7、10、12);第五类包括97个材料,分别为白和尚头(1、2、3、4、5、6、8、9、11),白麦(14、15、16、18、20),白芒糙(22、25、26、27),出山豹(29、30、31、33),二芒麦(37、38、46、48),和尚头(50、53、54、55、56、57),红和尚头(59、60、61、63、64、65、67、68、70),红芒白(71、73、75、76、77、80、81),红芒糙(83、84、86、87、88、90),红芒红(94、95、98、100),红蚰子头(101、102、103、105、106、107、108),黄瓜先(109、110、112、113、114、115、117、119),三月黄(120、121、122、124、125、127、129、131、132、133、134、135),五花头(136、137、140、143、147),蚰子头(149、150、151、152、153、154、155),具体见图1。从分类结果来看,相同名称的小麦地方品种多聚在一类。

图1 155份材料的聚类分析Fig.1 Dendrogram of 155 landraces based on SSR detection
在聚类图中没有发现相同地理来源的品种被聚在一起,说明这些地方品种的亲缘关系与材料的名称有一定的关系,与地理来源无关。
3 讨 论
小麦地方品种具有植株较高、易倒伏、产量低、结实率低等缺陷,使得长期以来育种家缺乏对它的关注。近年来由于选育品种的遗传基础越来越狭窄,迫切需要开发利用新的育种材料。小麦地方品种所具有的抗病、广适、遗传多样性丰富等优点逐渐被发掘[15]。本研究所涉及到的试验材料中就发现一些具有优异性状的品种,例如:出山豹(栾川,30)穗粒数达63.1粒,为多花多粒品种;红蚰子头(郏县,107)穗粒数为57.0粒,为多花多粒品种;黄瓜先(禹县,109)蛋白质含量17.1%、稳定时间30.5 min为优质品种;三月黄(温县,120)成熟期为5月24日为早熟品种。
本研究对15组相同名称的小麦地方品种进行了遗传多样性分析,虽然15组小麦地方品种的材料数量不同,但经遗传分析发现,这15组小麦地方品种的等位变异数和多态性信息含量都很高,说明这些地方品种具有丰富的遗传多样性。
155份材料中有部分材料含有特有条带,该等位变异的分布频率为0.6%,说明具有这些条带的材料可能具备其他材料所没有的特性,值得进一步研究。
由于本试验所用的小麦地方品种为科研人员自民间采集而来,受当时科研条件的限制,这些地方品种仅依据农艺性状及地理来源加以归类整理。随着科技的进步,利用分子标记技术分析品种间亲缘关系已成为常见的科研手段,本试验就是利用SSR技术对同名的小麦地方品种进行分析,结果发现白和尚头组中有3份材料、出山豹组中有2份材料、三月黄组中有2份材料出现遗传相似系数为1.00的情况,说明这些材料存在同名同质的现象,对于这部分材料,参照其农艺性状等特征可以将其合并,这有利于地方品种的保存、管理和开发利用。虽然大多数小麦地方品种间遗传相似系数小于1.00,但是一些遗传相似系数比较高的品种,也可能是同一个品种或同一个品种的变异,因为试验本身存在一定的局限性,并且小麦地方品种在繁殖更新过程中也会产生一定的变异[16],因此这有待于我们进一步的研究。
155份材料聚类分析结果显示,同名小麦地方品种多聚到一类,以蚰子头组最明显,7份蚰子头组的材料被分为一类,这说明同名小麦地方品种内的亲缘关系较近。这155份材料中,有部分材料来源于同一地方,聚类分析结果没有发现来源地相同的材料被明显聚在一类,说明这些品种的亲缘关系与地理来源无关;同时对155份材料整体聚类分析后发现,名称相同的材料在较近的相似系数内被聚在一起,这说明同名地方品种间的亲缘关系较近,这些同名的品种有可能是同一品种或同一品种的变异种。
[1]Huang X Q,Hsam S L K,Zellet F J.Chromosomal location of genes for resistance to powdery mildew in common wheat(TriticumaestivumL.em.Thcl1.) 4.Gene Pm24 in Chinese landrace Chiyacao [J].TheoreticalandAppliedGenetics,1997,95(5-6):950-953.
[2]韩德俊,张培禹,王琪琳,等.1980份小麦地方品种和国外种质抗条锈性鉴定与评价 [J].中国农业科学 2012,45(24):5013-5023.
Han D J,Zhang P Y,Wang Q L,etal.Identification and evaluation of resistance to stripe rust in 1980 wheat landraces and abroad germplasm [J].ScientiaAgriculturaSinica,2012,45(24):5013-5023.
[3]Lin F,Kong Z X,Zhu H L,etal.Mapping QTL associated with resistance to Fusarium head blight in the Nanda 2419×Wang-shuibai population I.Type II resistance [J].TheoreticalandAppliedGenetics,2004,109(7):1504-1511.
[4]蔡士兵,曹 旸,方先文.江苏省白皮小麦地方品种抗穗发芽性的研究 [J].江苏农业学报,1999,15(1):7-11.
Cai S B,Cao Y,Fang X W.A study on the resistance to preharvest sprouting in white-grained wheat landraces originated from Jiangsu province [J].JiangsuJournalofAgriculturalSciences,1999,15(1):7-11.
[5]王子宁,郭北海,李洪杰,等.小麦(T.aestivum)Waxy-D1基因缺失材料的发现及分析 [J].作物学报,2000,26(3):257-260.
Wang Z N,Guo B H,Li H J,etal.The discovery and analysis of wheat cultivar(T.aestivum) withWx-D1(nul1) Gene [J].ActaAgronomicaSinica,2000,26(3):257-260.
[6]陈华萍,邓 婷,苟璐璐,等.四川小麦地方品种农艺性状与品质性状的多样性分析[J].麦类作物学报,2008,28(6):960-964.
Chen H P,Deng T,Gou L L,etal.Diversity analysis of agronomic and quality characters of wheat landraces in Sichuan[J].JournalofTriticeaeCrops,2008,28(6):960-964.
[7]吴秀亭,张正斌,徐 萍,等.黄淮小麦农艺性状演变趋势 [J].中国农业科学,2013,46(18):3930-3938.
Wu X T,Zhang Z B,Xu P,etal.Agronomic traits development and evolution of winter wheat in Huanghuai basin [J].ScientiaAgriculturaSinica,2013,46(18):3930-3938.
[8]马晓岗,李凤珍,王晓辉,等.青海省小麦种质材料醇溶蛋白的遗传多样性分析 [J].麦类作物学报,2012,32(6):1060-1065.
Ma X G,Li F Z,Wang X H,etal.Gliadin genetic diversity analysis of wheat cultivars and germplasms from Qinghai province [J].JournalofTriticeaeCrops,2012,32(6):1060-1065.
[9]陈雪燕,王亚娟,雒景吾,等.陕西省小麦地方品种主要性状的遗传多样性研究 [J].麦类作物学报,2007,27(3):456-460.
Chen X Y,Wang Y J,Luo J W,etal.Genetic diversity in main characters of wheat landraces in Shaanxi province [J].JournalofTriticeaeCrops,2007,27(3):456-460 .
[10]张玲丽,王 辉,李立会,等.中国小麦地方品种大青芒遗传多样性研究 [J].中国农业科学,2007,40(8):1579-1586.
Zhang L L,Wang H,Li L H,etal.Genetic diversity analysis of common wheat landrace Daqingmang in various growing areas [J].ScientiaAgriculturaSinica,2007,40(8):1579-1586.
[11]李正玲,许为钢,张清珍,等.河南省地方小麦品种醇溶蛋白的遗传多样性分析 [J].麦类作物学报,2008,28(4):582- 587.
Li Z L,Xu W G,Zhang Q Z,etal.Genetic heterogeneity and diversity within and among Henan wheat landraces with the same name as revealed by gliadin composition [J].JournalofTriticeaeCrops,2008,28(4):582- 587.
[12]柴建芳,刘 旭,贾继增.一种适于PCR扩增的小麦基因组DNA快速提取法 [J].植物遗传资源学报,2006,7(2):246-248.
Chai J F,Liu X,Jia J Z.A rapid isolation method of wheat DNA suitable for PCR analysis [J].JournalofPlantGeneticResources,2006,7(2):246-248.
[13]Botstein D R,White R L,Skolnick M,etal.Construction of a genetic linkage map in man using restriction fragment length polymorphisms [J].AmericanJournalofHumanGenetics,1980,32:314-331.
[14]Nei M.Analysis of gene diversity in subdivided populations [J].ProceedingsoftheNationalAcademyofSciences,1973,70(12):3321-3323.
[15]郑殿升.中国作物遗传资源的多样性 [J].中国农业科技导报,2000,2(2):45-49.
Zheng D S.Diversity of crop genetic resources in China [J].ReviewofChinaAgriculturalScienceandTechnology,2000,2(2):45-49.
[16]张舒娜,高爱农,杨欣明,等.小麦地方品种繁殖更新过程中的遗传多样性变化分析 [J].植物遗传资源学报,2012,13(1):57-65.
Zhang S N,Gao A N,Yang X M,etal.Genetic diversity change of wheat landraces in regeneration [J].JournalofPlantGeneticResources,2012,13(1):57-65.
Genetic Diversity of Henan Wheat Landraces with the Same Name Detected by SSR Markers
LI Zhengling,HU Lin,WANG Huiwei,DONG Haibin,LI Yan,LI Chunxin,XU Weigang
(Wheat Research Center, Henan Academy of Agricultural Sciences/National Laboratory of Wheat Engineering/Key Laboratory of Ministry of Agriculture for Wheat Biology, Genetics and Breeding in Central Huang-huai Region/Key Laboratory of Henan Province for Wheat Biology, Zhengzhou,Henan 450002, China)
To analyze genetic diversity of the wheat landraces (OryzasativaL.)with the same name, fifteen groups with the same names including Baiheshangtou, Baimai, Baimangcao and Chushanbao etc., including 155 wheat landraces were analyzed on simple sequence repeat (SSR) polymorphisms. For each group of landraces with the same name, the number of SSR alleles ranged from 47 to 79 across 15 groups, and the average genetic similarity coefficient ranged from 0.66 to 0.93, and the polymorphism information contents (PIC) ranged from 0.856 to 0.936. Among different groups of landraces, there were 143 polymorphism alleles detected among 155 landraces, and the average genetic similarity coefficient was 0.90 ranging from 0.75 to 1.00, and the PIC was 0.981. Cluster analysis identified seven cultivars from three groups have genetic similarity coefficient of 1.00. The identical sample has not been found among the landrace groups. This study indicates that there is abundant genetic diversity of wheat landraces from Henan, even for the landraces with the same name. The study also indicates that SSR is a useful marker system to study genetic relationship among wheat landraces.
Wheat landraces; SSR; Genetic relationship; Genetic diversity
the genetic similarity coefficient is 1.00
时间:2016-05-10
2015-11-23
2015-12-22
国家科技支撑计划项目(2011BAD07B01);国家现代农业产业技术体系专项资金项目(CARS-3-1-9);国家自然科学基金项目(31371707)
E-mail:lxlandlzl@163.com
许为钢(E-mail: xuwg1958@163.com)
S512.1;S330
A
1009-1041(2016)05-0564-07
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160510.1623.008.html
猜你喜欢
区域治理(2022年40期)2022-11-27
作物学报(2022年6期)2022-04-08
西江月(2021年3期)2021-12-21
国际医学放射学杂志(2021年5期)2021-10-22
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
麦类作物学报(2018年4期)2018-05-11
第一财经(2017年36期)2017-09-25
星海音乐学院学报(2017年3期)2017-08-24
