
一株冬虫夏草相关菌株的生物学特性
2016-10-31 03:19朱玉兰
浙江农业学报 2016年2期
谢 放,张 育,朱玉兰
(兰州交通大学 化学与生物工程学院,甘肃 兰州 730070)
一株冬虫夏草相关菌株的生物学特性
谢放,张育,朱玉兰
(兰州交通大学 化学与生物工程学院,甘肃 兰州 730070)
从冬虫夏草虫体上分离得到一株真菌,对其生物学特性和侵染力,及其与冬虫夏草菌的相互关系进行了初步研究,并对其进行了分子鉴定。结果表明,该菌丝生长的最适温度为20~23 ℃,最适pH为7.50,最适碳氮源分别是蔗糖和麸皮,对蝙蝠蛾幼虫的侵染力为100%,并在与冬虫夏草菌平板对峙生长中发现其对冬虫夏草菌的生长抑制率高达43.37%。PCR结果显示,从菌丝体中扩增获得了560 bp左右的一段ITS序列,系统发育分析表明,该菌是白僵菌,GenBank登录号为KP284435。
白僵菌;冬虫夏草;生物学特性;侵染力
冬虫夏草是冬虫夏草菌侵染蝙蝠蛾幼虫而形成的虫菌复合体。长期以来,冬虫夏草菌的分离鉴定一直备受关注,但迄今仍未实现科赫法则条件下冬虫夏草菌的认定。前人对冬虫夏草菌有以下不同的看法:郭英兰等[1]、魏鑫丽等[2]、刘锡琎等[3]通过组织和子囊孢子多途径、多批次分离试验证明,中国被毛孢就是冬虫夏草菌的无性型;赵锦等[4]、李增智等[5]利用rDNA的ITS序列分析和RAPD-PCR技术也得到了中国被毛孢是冬虫夏草菌的无性型的相关证据;朱佳石等[6-7]、高凌等[8]认为冬虫夏草菌是多种突变基因型菌(一组真菌);姚艺桑等[9]通过分子生物学方法证实天然冬虫夏草子座中存在多个突变基因型菌,并随着冬虫夏草的成熟,其在僵虫体和子座中差异性表达;朱佳石等[6]也发现,蝙蝠蛾拟青霉和中国被毛孢真菌共存于冬虫夏草的虫体和子座中;许多研究认为冬虫夏草菌是多种真菌的总称[10-14],幸兴球[11]指出冬虫夏草的无性型可能由至少8种真菌组成,梁宗琦等[12]则提出冬虫夏草是一个多种菌共存的复杂统一的微生态系统,张永杰等[13]、杨金玲等[14]也认为天然冬虫夏草中除了中国被毛孢外还存在多种其他真菌。以上这些研究均是围绕冬虫夏草菌展开的,但有关冬虫夏草相关菌对冬虫夏草菌的影响,以及相关菌株与冬虫夏草菌之间的对峙研究和回接侵染蝙蝠蛾幼虫方面的研究均相当有限。本试验在冬虫夏草虫体上分离得到一株真菌,分析了该菌的生物学特性,以及其对蝙蝠蛾幼虫的侵染力;并且通过平板对峙试验对该菌株与冬虫夏草菌之间的关系进行了研究,以期为冬虫夏草相关菌株的开发利用和冬虫夏草菌侵染蝙蝠蛾幼虫的研究提供一定的理论依据。
1 材料与方法
1.1材料
1.1.1菌种
1-1菌株分离自甘肃省甘南州夏河县榨油沟乡拉代牧场冬虫夏草的虫部。供试冬虫夏草菌(FN570054,HirsutellasinensisFang. X&J. H. Li Fang. X,2013-01-05)[15],现保存于兰州交通大学化学与生物工程学院生物种苗工程研究所。蝙蝠蛾幼虫采自甘肃省武威市天祝县抓喜秀龙乡代乾牧场。
1.1.2培养基
牛奶培养基:鲜牛奶200 mL,马铃薯200 g,葡萄糖20 g,水解乳蛋白5 g,酵母粉1 g,硫酸镁0.2 g,磷酸二氢钾1 g,复合维生素B 1片,琼脂15 g,蒸馏水1 000 mL,pH 6.5。
PDA培养基:马铃薯200 g,葡萄糖20 g,琼脂15 g,蒸馏水1 000 mL,pH自然。
1.1.3其他
HP Fungal DNA Kit试剂盒、引物、Taq酶、dNTP 等购自上海生物工程公司。
1.2方法
1.2.1形态学观察及鉴定
观察记录菌落、菌丝和孢子形态特征,并用数码相机拍照,用MOTIC B显微镜观察、测量和拍照微观产孢结构。
1.2.2最佳温度和最适pH筛选
据单因子试验设计,将活化好的菌丝体块用9 mm的打孔器打孔分别接种于培养基中,倒置于16,20,23,26 ℃培养箱中恒温避光培养;将活化好的菌丝接种于pH值分别为4.5,5.5,6.5,7.5,8.5的培养基,倒置于23 ℃恒温培养。每个处理均设3组平行,观察记录。
1.2.3最佳碳氮源筛选
以牛奶培养基中20 g葡萄糖的含碳量为标准,分别以相等含碳量的蔗糖、麦芽糖、可溶性淀粉代替基础培养基中的葡萄糖,并以不添加碳源的基础培养基为对照;以牛奶培养基中200 mL牛奶的含氮量为标准,分别以相等含氮量的蚕蛹粉、黄豆粉、麸皮代替基础培养基中的牛奶,并以不添加氮源的基础培养基做对照。
1.2.4平板对峙试验
将活化好的1-1菌株和冬虫夏草菌分别用9 mm的打孔器打孔,放置在牛奶培养基中相距3 cm的位置,16 ℃培养。每个处理设3组平行,观察记录,并按以下方法计算抑菌率和拮抗系数。
抑菌率/%=[(对照菌落半径-处理菌落半径)/对照菌落半径]×100
拮抗系数分级标准[16]:Ⅰ,白僵菌占据平皿100%;Ⅱ,白僵菌占据平皿≥3/4;Ⅲ,白僵菌占据平皿<3/4,≥2/3;Ⅳ,白僵菌占据平皿<2/3,≥1/3;Ⅴ,白僵菌占据平皿<1/3;Ⅵ,冬虫夏草菌占据平皿100%。
1.2.5回接侵染试验
用涂有孢子悬浮液的胡萝卜喂养蝙蝠蛾幼虫,每隔5 d换食1次,每天观察1次,并拍照记录幼虫感染情况。
1.2.6菌丝日均长速的测定
菌丝将近长满平皿而未满平皿时作为测量终点进行测量。
菌丝日均长速(mm·d-1)=[(菌落直径-9)/2]/培养天数,最后取3组平行的均值。
1.2.7菌落干重的测定
采用重量法进行测定[17]。
1.2.8DNA提取和PCR扩增
在PDA培养基上23 ℃培养10 d,取新鲜培养物,用试剂盒提取总DNA。PCR扩增:引物ITS1:5′-TCCGTAGGTGAACCTGCGG-3′;ITS4:5′-TCCTCCGCTTATTGATATGC-3′。PCR反应体系(25 μL):Mix 12.5 μL,引物ITS1和引物ITS4均为0.5 μL,模板1 μL,ddH2O 10.5 μL。PCR条件:95 ℃预变性5 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,30个循环;72 ℃延伸10 min。产物检测:在添加Goldview的1.0%的琼脂糖凝胶中电泳20 min检测。
1.2.9序列分析
PCR产物的纯化和DNA测序:由上海美吉生物科技有限公司完成。
2 结果与分析
2.1形态描述
该菌在PDA培养基上生长迅速,菌落白色,菌丝呈絮状,边缘整齐,背面略显黄色,该菌在生长后期菌落边缘长出基础宽,顶端细的孢梗束,孢梗束为白色,均匀分布在菌落边缘,其长度可达到1.0~1.5 cm。菌丝透明,有隔,表面光滑,粗细均匀,宽2~3 μm,分生孢子梗呈瓶梗状,孢子椭圆形,链状,大小为(3~4 )μm×(1.5~2.5) μm(图1)。
2.2PCR扩增
用试剂盒提取1-1菌株的基因组DNA为模板,选取ITS1和ITS4作为一对通用引物,进行PCR扩增,结果如图2所示,所得的产物片段大小为560 bp左右,条带明亮清晰,无拖尾现象,表明该法提取的基因组DNA质量较好,并满足基于PCR反应的后续分子生物学试验研究。

a. 菌落形态; b. 菌丝结构; c,d. 孢子梗和孢子结构。图1 1-1菌株的菌落、菌丝体和孢子形态Fig.1 The morphology of colony, mycelium and spores of strain 1-1

M: 分子量标准。图2 菌丝体基因组DNA PCR扩增的ITS片段Fig.2 ITS-PCR product amplificated from genomic DNA of mycelium
2.3系统发育分析
将测序结果提交至GenBank,获得登录号为KP284435。在NCBI网站上用BLAST程序进行序列比对,选取相似性较高的菌株,用ClustalX 1.83软件对1-1菌株和一些已知种进行了多序列比对,用MEGA 4.1软件,用邻接法自展值为1 000次的计算构建系统发育树(图3)。结果表明,1-1菌株与白僵菌的遗传关系最近,相似度高达100%,并以100%的支持率处于一个分支中,因此鉴定该菌属于白僵菌属。

图3 依据ITS基因序列构建的系统发育树Fig.3 Neighbour-joining phylogenetic tree based on ITS gene sequences
2.4最适温度筛选
1-1菌株在不同温度下的生长情况见表1,该菌在16~26 ℃范围内都能生长,但长速和干重存在差异:在20 ℃时菌丝长速最快,而在23 ℃时,积累的干重最多;温度超过23 ℃时,菌落干重积累量随之减少,因此,该菌菌丝生长的最适温度为20~23 ℃,而在刘召等[18]的研究中白僵菌菌丝最适生长温度为28 ℃,可能不同地理来源的菌株,其菌丝生长的温度范围不同[19]。
表1不同温度对菌丝生长的影响
Table 1Effect of different temperatures on the growth of mycelia
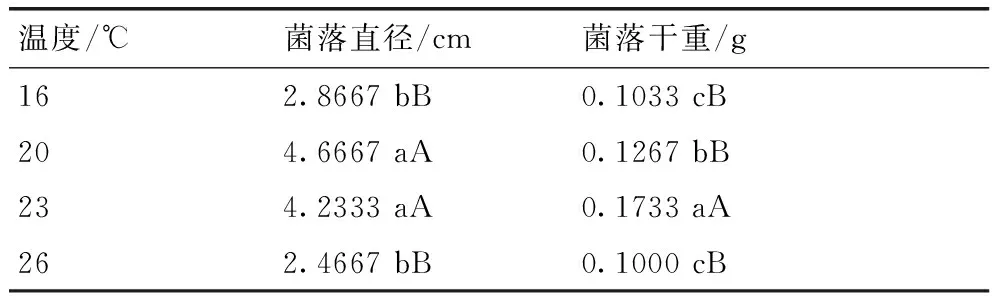
温度/℃菌落直径/cm菌落干重/g162.8667bB0.1033cB204.6667aA0.1267bB234.2333aA0.1733aA262.4667bB0.1000cB
注:同一列不同行数据后没有相同小写字母表示差异显著(P<0.05),没有相同大写字母表示差异极显著(P<0.01)。下同。
2.5最佳pH筛选
pH试验结果表明(表2),1-1菌株在pH值为4.50~8.50的范围内均能生长,且在不同pH值下菌株生长状况差异不大,pH值为4.50时,菌丝生长缓慢,菌丝稀疏;随着pH值的增大菌丝长速和菌落干重均表现出先增加后减弱的趋势。其中pH值为7.50时菌丝的生长速率和干重均达最大,因此该菌丝最适生长pH为7.50。这与张晓敏等[20]对白僵菌的研究结果一致。
表2不同pH值对菌丝生长的影响
Table 2Effect of different pH values on the growth of mycelia

pH值菌落直径/cm菌落干重/g4.504.25000.0867bAB5.504.68330.1000bAB6.504.86670.1400aAB7.504.99330.1467aA8.504.56000.0800bB
2.6最适碳源筛选
由表3可知,1-1菌株在没有添加额外碳源的情况下也能正常生长,说明菌株能利用多种碳源,但长速和干重存在较大差异。其中以葡萄糖为碳源时菌丝长速最快,蔗糖次之,但总体而言,不同碳源对菌丝生长的影响差异性不大。就干重来说,以蔗糖为碳源时菌落质地紧密,长势好,积累的干重最多。因此,该菌的最适碳源为蔗糖,说明该菌能较好地利用双糖。试验中发现不同的碳源对菌株干重积累的影响与对菌丝长速的影响不同,可能是丰富的营养物质促使菌落密度的加厚和分枝的增多有关,如Suzuki等[21]研究表明;合适的碳氮营养素能够增加真菌中部分生长因子,更有利于菌丝生长。
表3不同碳源对菌丝生长的影响
Table 3Effect of different carbon sources on the growth of mycelia

碳源菌落直径/cm菌落干重/g葡萄糖4.7750aA0.1467aA麦芽糖3.7900bB0.1300aA蔗糖4.7000aA0.1767aA淀粉4.4333aA0.1367aA无碳源3.4500bB0.0800bB
2.7最佳氮源筛选
在没有添加额外氮源的情况下,1-1菌株依然生长良好,说明其能利用多种氮源,但长速和干重存在较大差异。其中不同的动植物蛋白对菌丝生长差异性不显著,而以麸皮为氮源的培养基中的菌落直径和干重积累量达到最大,牛奶次之(表4)。因此,该菌的最适氮源为麸皮,因为麸皮除能提供氮源外,还提供了有利于该菌生长的碳素营养和生长因子,从而促使营养平衡和物质转化。
表4不同氮源对菌丝生长的影响
Table 4Effect of different nitrogen sources on the growth of mycelia

氮源菌落直径/cm菌落干重/g牛奶3.19000.0667abA蚕蛹粉3.09000.0467cAB黄豆粉2.71000.0533bcAB麸皮3.26670.0733aA无氮源2.89330.0367cB
2.81-1菌株与冬虫夏草菌之间的关系
1-1菌株与冬虫夏草菌之间的相互关系如图4所示,对照组中的冬虫夏草菌在20 d时菌丝生长进入对数期,并随着时间的推移菌落半径也逐步增大,到30 d时菌落半径达1.9000 cm;而处理组中冬虫夏草菌的生长速度很慢,在第30天冬虫夏草菌菌落直径只有1.1663 cm。说明白僵菌对冬虫夏草菌的生长产生了明显的抑制作用,30 d时白僵菌对冬虫夏草菌的抑制率为43.37%,拮抗系数达到Ⅲ级。在培养后期发现白僵菌与冬虫夏草菌菌落发生重叠,重叠部分白僵菌将冬虫夏草菌覆盖,显微观察发现该区域中白僵菌菌丝与冬虫夏草菌菌丝并列生长。该菌与冬虫夏草菌之间的关系到底是空间竞争还是营养竞争,还需做进一步的试验。
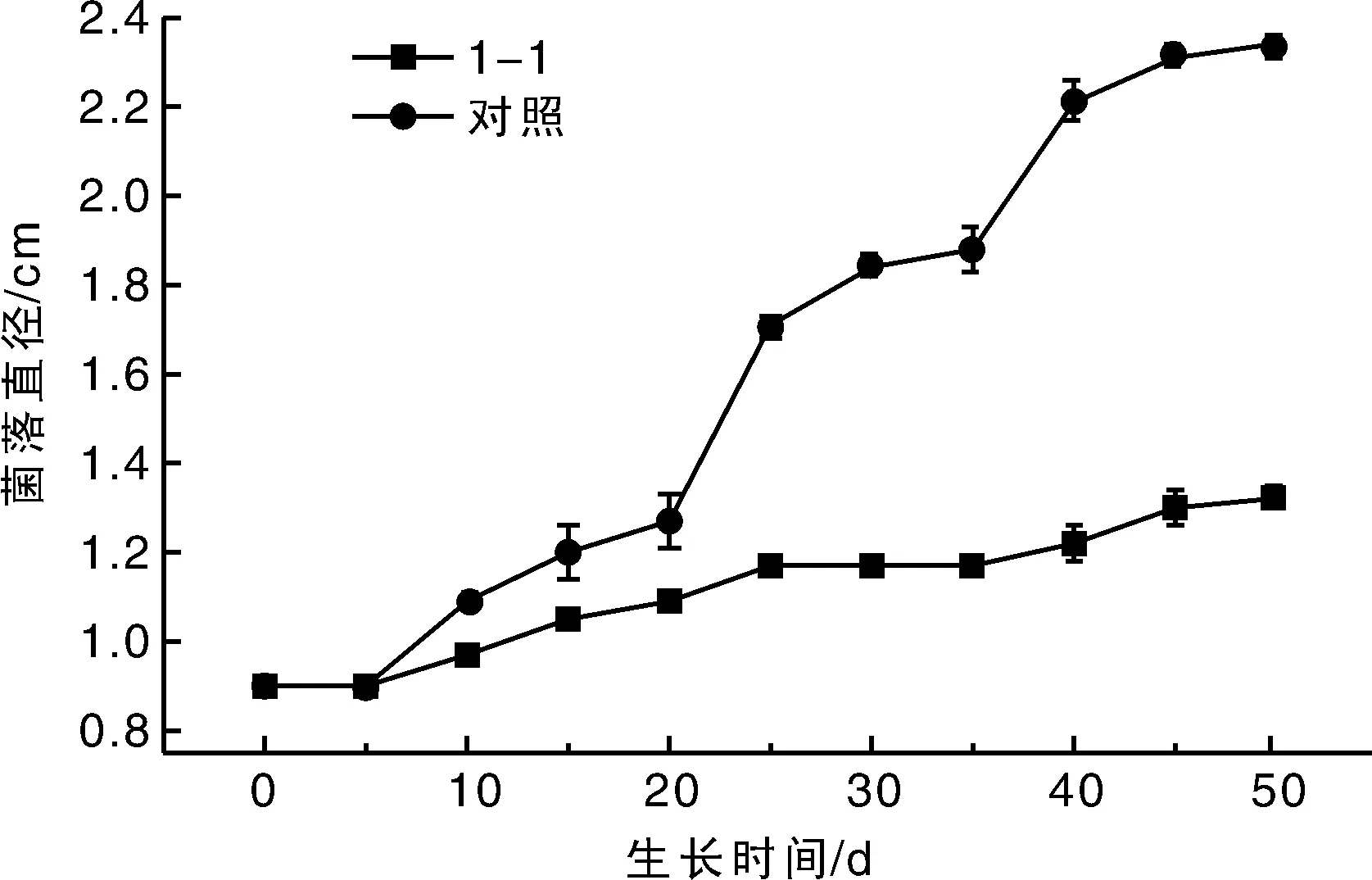
图4 平板对峙实验中该菌株对冬虫夏草菌的影响Fig.4 Effect of the strain 1-1 on Ophiocordyceps sinensis in table confrontation experiment
2.91-1菌株回接侵染蝙蝠蛾幼虫试验
1-1菌株侵染蝙蝠蛾幼虫后第6天,幼虫开始死亡,共死亡20头,菌株对蝙蝠蛾幼虫的侵染率高达100%。死亡的幼虫皮肤呈黄色,身体僵硬,死亡后第2天,幼虫身体开始长出均匀的菌丝,菌丝呈白色,质地疏松。第6天时菌丝布满全身,死亡9 d后菌丝开始大量生长,与培养基上的菌落形态一致,待死亡后第24天,可以看见边缘有孢梗束长出(图5-c箭头所示),说明该菌对蝙蝠蛾幼虫有较高的侵染率,是蝙蝠蛾幼虫的致病菌。

a: 死亡第2天; b: 死亡第15天; c: 死亡第24天; d: 死亡第50天。图5 蝙蝠蛾幼虫侵染试验结果Fig.5 Results of Hepialidae larva infection test
3 讨论
在平板对峙试验中,1-1菌株对冬虫夏草菌抑制效果明显,其具体原因是营养竞争,空间竞争,还是产生了其他的抗生物质?由于两者在对峙中没有明显的抑菌分界线,且冬虫夏草菌落生长没有表现出偏心现象,可判断他们之间没有拮抗作用;其次,在对峙生长后期,该菌继续在冬虫夏草菌的菌落上生长直至全部覆盖,通过显微观察发现,该菌菌丝与冬虫夏草菌菌丝并列生长,对该现象的深入分析还需进一步的细胞学试验验证。如果细胞学试验发现,菌丝被穿透、细胞萎缩等被寄生后的特有现象,方可确定该菌与冬虫夏草菌之间是重寄生关系,否则就是营养竞争或者空间竞争关系。再次,这种抑制作用是否受到培养条件的影响也需进一步研究,作者曾添加蝙蝠蛾幼虫体液培养冬虫夏草菌,表现出生长迅速,暗示在更适宜的条件下,冬虫夏草菌生长表现可能完全不同。如果仅仅是营养竞争关系,则抑制效果就可能不同。
回接侵染试验结果说明,该菌也是寄生蝙蝠蛾幼虫的致病菌,但在冬虫夏草虫体上分离得到意味着它可能有一定作用。马少丽等[23]的研究表明,对蝙蝠蛾幼虫具有侵染力的相关菌株与冬虫夏草菌的复合侵染可将侵染率由原来的4%提高至50%。本供试菌对蝙蝠蛾幼虫具有很高的侵染力,但是否也具有提高冬虫夏草菌对蝙蝠蛾幼虫的侵染率,还需要复合侵染试验加以验证。
[1]郭英兰,肖培根,魏江春.论冬虫夏草生物学与可持续利用[J].中国现代中药,2010,12(11):3-8.
[2]魏鑫丽,印象初,郭英兰,等.冬虫夏草及其相关类群的分子系统学分析[J].菌物学报,2006,25(2):192-202.
[3]刘锡琎,郭英兰,俞永信,等.冬虫夏草菌无性阶段的分离和鉴定[J].真菌学报,1989(8):35-40.
[4]赵锦,王宁,陈月琴,等.冬虫夏草无性型的分子鉴别[J].中山大学学报(自然科学版),1999,38(1):121-123.
[5]李增智,黄勃,李春如,等.确证冬虫夏草无性型的分子生物学证据I.中国被毛孢与冬虫夏草之间的关系[J].菌物系统,2000,19(1): 60-64.
[6]朱佳石,郭英兰,姚艺桑,等.冬虫夏草成熟过程中中国被毛孢和蝙蝠蛾拟青霉DNA共存及竞争增值力、化学成分变化[J].菌物研究, 2007,5(4):214-224.
[7]朱佳石,赵家刚,高凌,等.至少6个突变基因型冬虫夏草菌在冬虫夏草子座中表达的动态变化[J].菌物研究,2012,10(2):100-112.
[8]高凌,李晓红,赵建晴,等.MassARRAY单核苷酸多态性(SNP)MALDI-TOF质谱基因分型法检测未成熟冬虫夏草子座中多个冬虫夏草菌突变基因型[J].北京大学学报(医学科学版),2011,43(2):259-266.
[9]姚艺桑,周妍娇,高凌,等.冬虫夏草菌及其突变基因型在冬虫夏草成熟过程中差异表达的动态变化[J].菌物研究,2011,9(1):37-49.
[10]蒋毅,姚一建.冬虫夏草无性型研究概况[J].菌物系统, 2003, 22(1):161-176.
[11]幸兴球.药用虫草可能不止一种真菌的商讨[J].微生物学通报,1992,19(5): 310-312.
[12]梁宗琦,韩燕峰,梁建东,等.冬虫夏草Ophiocordycepssinensis研究中几个值得关注的问题[J].微生物学通报,2010,37(11): 1692-1697.
[13]张永杰,孙炳达,张姝,等.分离自冬虫夏草可培养真菌的多样性研究[J].微生物学报,2010,29(4):518-527.
[14]杨金玲,肖薇,何惠霞,等.蝙蝠蛾拟青霉与冬虫夏草关系的分子系统学研究[J].药学学报,2008,43(4):421-426.
[15]姚一建.中国菌物名录数据库建设[C]//中国菌物学会. 2012年中国菌物学会学术年会会议摘要.北京:中国菌物学会,2012.
[16]BELL D K, WELLS H D, MARKHAM C R.Invitroantagonism ofTrichodermaspecies against six fungal plant pathogens[J].Phytopatholagy,1982,72(4):379-382.
[17]刘晓鹏,姜宁,向东山,等.金针菇深层发酵条件及水浴提取菌丝体多糖的研究[J].食品科学, 2008, 29(7): 437-440.
[18]刘召,徐丽,雷仲仁.温度和光照对白僵菌加拿大1号菌株菌落及产孢量的影响[J].西南大学学报(自然科学版),2014,36(5):1-6.
[19]VIDAL C, FARGUES J, LACEY L A. Intraspecific variability ofPaecilomycesFumosoroseus: Effect of temperature on vegetative growth[J].JournalofInvertebratePathology, 1997, 70(1):18-26.
[20]张晓敏,杨帆,邢秀霞,等.不同条件对球孢白僵菌分生孢子萌发的影响[J].内蒙古农业大学学报,2010,31(4):43-46.
[21]SUZUKI G T, FLEURI L, MACEDO G A. Influence of nitrogen and carbon sources on riboflavin production by wild strain ofCandidasp[J].FoodandBioprocessTechnology,2012,5(2):466-473.
[22]夏龙荪.白僵菌对几种常见植物病原菌的拮抗作用研究[J].中国生物防治学报,2013,29(2):324-330.
[23]马少丽,张宗豪,徐海峰,等.拉脊蝠蛾幼虫肠道菌群分析及侵染实验[J].食用菌,2014(3):28-30.
(责任编辑张韵)
Biological characteristics of a strain of Ophicordyceps sinensis related fungi
XIE Fang, ZHANG Yu, ZHU Yu-lan
(SchoolofChemicalandBiologicalEngineering,LanzhouJiaotongUniversity,Lanzhou730070,China)
A preliminary study was carried out to investigate the biological characteristics, infection rate of a fungus isolated fromOphicordycepssinensis, as well as its relationship withOphicordycepssinensis, and the molecular identification was analyzed. The results showed that the optimum temperature and pH for culture was 20 to 23 ℃ and 7.50 respectively. The optimum carbon sources and nitrogen sources was sucrose and bran. The infection rate onHepialidaelarva was 100%. The inhibition rate onOphicordycepssinensiswas 43.37% in flat confrontation test. PCR results showed that about 560 bp ITS fragments were amplified from the mycelium. Phylogenetic analysis indicated that the fungi belonged toBeauveria, and GenBank accession number is KP284435.
Beauveria;Ophicordycepssinensis; biological characters; infection
10.3969/j.issn.1004-1524.2016.02.21
2015-06-15
甘肃省科技支撑基金(212256)
谢放(1962—),男,甘肃兰州人,博士,副教授,研究方向为资源与环境微生物。E-mail:1004389243@qq.com
Q935
A
1004-1524(2016)02-0306-06
谢放,张育,朱玉兰. 一株冬虫夏草相关菌株的生物学特性[J].浙江农业学报,2016,28(2): 306-311.
猜你喜欢
发明与创新·小学生(2021年12期)2021-01-06
军事文摘(2020年22期)2021-01-04
湖北林业科技(2019年2期)2019-05-05
中成药(2017年5期)2017-06-13
小溪流(画刊)(2016年12期)2017-02-04
汉语世界(2016年3期)2016-11-16
上海农业学报(2016年2期)2016-10-27
广西林业科学(2016年4期)2016-03-16
微型小说选刊(2015年5期)2015-06-05
小学生·多元智能大王(2014年5期)2014-07-24
