
广西地区常见棕榈科植物光合特性研究
2016-10-25 01:02高传友
水土保持研究 2016年2期
高传友
(广西职业技术学院, 南宁 530226)
广西地区常见棕榈科植物光合特性研究
高传友
(广西职业技术学院, 南宁 530226)
以广西南宁市不同棕榈科植物(棕竹、蒲葵、槟榔)为试材,研究和比较了其光合特性。结果表明:不同棕榈科植物株高、茎粗和冠幅均表现为槟榔>蒲葵>棕竹,叶面积指数和比叶重均表现为蒲葵>槟榔>棕竹,根区土壤pH表现为棕竹>槟榔>蒲葵,根区土壤养分大致表现为蒲葵>槟榔>棕竹,局部有所波动,其中不同棕榈科植物根区有机碳含量差异不显著(p>0.05)。不同棕榈科植物光饱和点(LSP)、光补偿点(LCP)、表观量子产额(AQY)、羧化效率(CE)、气孔限制值(Ls)、水分利用效率(WUE)、光合氮利用效率(NUE)和光合磷利用效率(PUE)基本表现为蒲葵>槟榔>棕竹,局部有所波动,与叶面积指数和比叶重以及其根区土壤养分变化趋势相一致,其中不同棕榈科植物Ls差异不显著(p>0.05)。不同棕榈科植物一天各时期净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)和胞间CO2浓度(Ci)基本表现为蒲葵>槟榔>棕竹规律,净光合速率(Pn)日变化均呈双峰曲线,气孔限制导致了光合“午休”现象,胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(EVAP)日变化随时间呈“W”变化规律。通过SPSS最佳曲线拟合不同棕榈科植物的Pn与PAR,CO2浓度和温度变化呈一致的二次曲线关系。相关性分析表明,不同棕榈科植物光合特性由多种因素共同决定,这些因素之间也会相互影响,其中Gs的相关系数较大。
广西; 棕榈科植物; 光合特性
棕榈科(Palmae)植物属于种子植物门被子植物亚门单子叶植物纲初生目,是热带地区重要的代表植物[1]。目前全世界共约210余属,2 800~3 000种左右,主要分布在热带美洲、热带亚洲和太平洋及附近岛屿,热带非洲也有少量分布[2]。棕榈科植物在我国主要分布于海南、云南、广东、广西、福建、台湾等省,据不完全统计,我国棕榈科植物种质资源至今约有18属100多种[3-5]。棕榈科植物因其种类繁多、形态迥异等优点,被世界各国广泛种植和利用,常作为行道绿化树、庭园观赏树、园林景观树等广泛应用于热带和亚热带地区,是绿化、美化、净化、优化人类生活环境不可多得的优良树种[3]。自20世纪80年代,我国南方纷纷引种和栽培,前后出现了“王棕热”、“海枣热”等棕榈热[6]。迄今为止,国内已有很多学者对其进行了大量的研究,主要集中在园林应用、引种及逆境生理方面。黄志华等[7]根据棕榈植物的特点和景观功能,探讨了棕榈植物如何应用于园林造景;潘辉等[8]通过试验研究了棕榈科植物生长的主要环境影响因子;杨清等[9]通过棕榈植物的生物学和美学特征探讨其造景功能、景观构成等。在众多关于棕榈科植物的研究中,光照是植物生命活动中起重大作用的生存因子,棕榈植物需要在一定的光照条件下完成生长发育过程,但不同棕榈植物对光照强度的适应范围亦有所差别,棕榈科植物光合特性的研究尚不多见[3-5]。因此,本文以南宁市不同棕榈科植物(棕竹、蒲葵、槟榔)为试材,研究和比较了其光合特性及生理特性,并探讨了二者之间的相关性,为棕榈科植物在城市生态景观功能性植物的配置方面提供科学依据,使其在园林植物配植中能得到更好的应用。
1 材料与方法
1.1研究区概况
研究区(南宁市)属湿润的南亚热带季风气候,阳光充足,雨量充沛,霜少无雪,春秋两季气候温和,夏季炎热多雨,夏长冬短,多年平均日照时数为1827 h,年平均气温21.6℃,极端最高温度40.4℃,极端最低温度-2.4℃,1月份最冷,平均12.8℃,7月,8月份最热,平均28.2℃,近50年平均降水量1304 mm左右,多集中在夏季(6—8月),占全年降水量的52.3%,土壤类型较多,分布有棕壤土、红黏土、褐土、潮褐土、潮土等,全年无霜期高达345—360 d。
1.2测定方法
2015年7月,对南宁市建成区内主要的公园绿地(人民公园、狮山公园、金花茶公园和滨江公园)采集和调查不同棕榈科植物叶片(棕竹、蒲葵、槟榔),每个公园分别选取3处棕竹、蒲葵、槟榔,每一处分别随机选取3株(尽量包括大、中、小植株)棕榈科植物作为采样及调查的对象,将4个公园的棕榈科植物作为重复取样,并且在同一时间采集叶片样本和测定生长指标(现测株高和茎粗),采集叶片时带上聚乙烯塑料手套,分别从植株不同方向均匀采集上、中、下足够多的成熟叶片带回实验室处理,一部分置于5℃保温冰箱中用于扫描叶面积及烘干后测定其干重(叶片为10个),另一部分叶片粉碎后用于测量叶片N和P含量,同时四分法取每种植物根区土壤(0—20 cm)混合后除去大颗粒物质和根系,自然风干20 d后,过2 mm筛用于测定土壤养分含量。土壤pH值采用电极电位法(5∶1水土比浸提液)测定;土壤电导率采用电导法测定;有机碳含量采用重铬酸钾氧化外加热法测定;全磷含量采用NaOH碱溶—钼锑抗比色法测定;全氮含量采用半微量凯氏定氮法测定;碱解氮含量采用NaOH-H3BO3法测定;全钾含量采用乙酸铵浸提—火焰光度计法测。
(1) 光合气体交换测定。选择在生长旺盛、高温的7月份进行光合作用测定(晴天无风)。采用英国PP-Systems公司生产的CIRAS-2便携式光合仪,时间设定为6:00—18:00(晴天无风),每隔2 h测量1次,连续重复测定3 d,每个处理选择3株健康不同棕榈科植物重复测定,每株选取3年生的成熟叶片(从幼苗顶部数第3~6片完全展开的成熟叶),轮流测定以消除测定时间上的误差。红光光源设定为1 000 mol/(m2·s),温度25~30℃,CO2流量为300~500 mol/L,温度变化在19.1~32.4℃,辐射强度(PAR)在103~1 125 mol/(m2·s),空气相对湿度在17.6%~39.7%,仪器同时记录净光合速率[Pn,μmol/(m2·s)]、蒸腾速率[EVAP,mmol/(m2·s)]、气孔导度[Gs,mol/(m2·s)]、胞间CO2浓度(Ci,ppm)、环境CO2浓度(Ca)和光照强度(PAR),其中PAR和CO2浓度以外界条件为准。气孔限制值(Ls)和叶片瞬时水分利用效率(WUE)由公式计算得到Ls=1-Ci/Ca,WUE=Pn/EVAP。与此同时进行光合—光强响应特性的测定,CO2摩尔分数控制为350 μmol/mol,温度设定25℃左右,湿度为大气湿度的95%。设定光照度梯度为1 800,1 600,1 400,1 200,1 000,800,600,400,200,100,0 μmol/(m2·s),每个光照度下5 min后测定Pn值。根据Pn-PAR回归方程求出不同处理间的光补偿点(LCP)、光饱合点(LSP);表观量子效率(AQY)根据低于200 μmol/(m2·s)光照度下测定的叶片PFD-Pn曲线计算求得;通过测定CO2浓度下[250 μmol/(m2·s)]的光合速率制作出CO2-光合速率响应曲线,进而计算出叶片CO2补偿点和CO2羧化效率[10-11];光合—温度响应特性的测定,设定叶室PAR为900 μmol/(m2·s),CO2浓度为350 μmol/mol,湿度为大气湿度的99%,温度梯度为15,20,25,30,35℃。
(2) 叶面积及叶比重。应用数字扫描仪和图像处理技术测定叶面积(包括叶柄),叶比重(SLW)=单位面积叶干重/单位叶面积
(3) 株高。每个公园选取有代表性的3~5株,测定其绝对高度(从地面至植株最高部位的高度)
(4) 叶片养分测定。将采集的植物叶片,洗净65℃烘箱烘干,粉碎后过1.5 mm筛,凯氏定氮法测定叶片全氮含量,钒钼黄吸光光度法测定叶片全磷含量,光合作用氮、磷利用效率(NUE,PUE),如下公式求得:
光合作用氮、磷利用效率=净光合速率(Pn)/单位叶面积氮、磷含量[12]
1.3数据处理
数据处理采用Excel 2007软件,回归分析和统计检验利用SPSS 18软件。对数据进行方差齐性检验,满足方差齐性的情况下,采用LSD进行多重比较和单因素方差分析(one-way ANOVA)。利用光合速率和温度、CO2,PAR数据进行最佳回归函数的建立,由原始数据拟合得到的回归关系经统计学检验得到拟合度参数R2和AdjustR2,筛选最佳拟合曲线,分别在0.05,0.01水平检验相关系数的显著性。
2 结果与分析
2.1不同棕榈科植物生长特性及土壤特性
由表1可知,不同棕榈科植物株高、茎粗和冠幅均表现为槟榔>蒲葵>棕竹,叶面积指数和比叶重均表现为蒲葵>槟榔>棕竹,其中株高、叶面积指数和冠幅三种棕榈科植物差异均显著(p<0.05);槟榔茎粗显著高于蒲葵和棕竹(p<0.05),蒲葵和棕竹差异不显著(p>0.05);蒲葵比叶重显著高于棕竹和槟榔(p<0.05),棕竹和槟榔比叶重差异不显著(p>0.05)。三种棕榈科植物株高变化范围在58.15~523.41 cm,茎粗变化范围在15.74~25.81 cm,叶面积指数变化范围在13.78~18.94,比叶重变化范围在6.03~7.52,冠幅变化范围为156.45~254.38 cm。通过测定和比较不同棕榈科植物生长根区土壤特性,由表2可知,不同棕榈科植物根区土壤pH表现为棕竹>槟榔>蒲葵,蒲葵显著低于棕竹和槟榔(p<0.05);土壤电导率表现为蒲葵>槟榔>棕竹,三者之间差异均显著(p<0.05);土壤含水量表现为蒲葵>槟榔>棕竹,蒲葵显著高于棕竹和槟榔(p<0.05);土壤总孔隙度表现为蒲葵>槟榔>棕竹,三者之间差异不显著(p>0.05);土壤有机碳表现为蒲葵>槟榔>棕竹,三者之间差异均显著(p<0.05);土壤全氮表现为蒲葵>棕竹>槟榔,蒲葵显著高于棕竹和槟榔(p<0.05);土壤全钾表现为蒲葵>槟榔>棕竹,蒲葵显著高于棕竹和槟榔(p<0.05)。

表1 不同棕榈科植物生长特性
注:同列不同小写字母表示差异显著(p<0.05),下同。
2.2不同棕榈科植物叶片生理特性
由图1可知,不同棕榈科植物光饱和点(LSP)变化范围为865~1 135 μmol/(m2·s),依次表现为蒲葵>槟榔>棕竹,蒲葵显著高于棕竹和槟榔(p<0.05);光补偿点(LCP)变化范围为68~97 μmol/(m2·s),依次表现为蒲葵>槟榔>棕竹,三者之间差异均显著(p<0.05);表观量子产额(AQY)在0.019 7~0.025 3,依次表现为蒲葵>槟榔>棕竹,蒲葵和槟榔之间差异不显著(p>0.05);羧化效率(CE)变化范围为0.049~0.053,依次表现为蒲葵>槟榔>棕竹,蒲葵显著高于棕竹和槟榔(p<0.05);气孔限制值(Ls)在0.85~0.94,依次表现为蒲葵>槟榔>棕竹,三者之间差异并不显著(p>0.05);水分利用效率(WUE)在5.13~6.01,依次表现为蒲葵>棕竹>槟榔;光合氮利用效率(NUE)在5.13~8.56,依次表现为蒲葵>棕竹>槟榔,三者之间差异均显著(p<0.05);光合磷利用效率(PUE)在2.86~3.67,依次表现为蒲葵>槟榔>棕竹,蒲葵和槟榔之间差异并不显著(p>0.05)。
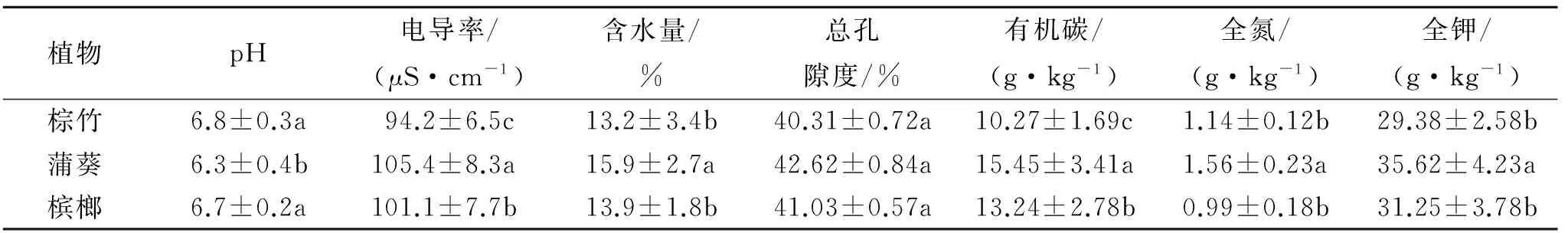
表2 不同棕榈科植物根区土壤特性(0-20 cm)
注:不同小写字母表示差异显著(p<0.05),下同。
图1不同棕榈科植物叶片生理特性
2.3不同棕榈科植物叶片光合特性
不同棕榈科植物叶片光合特性如图2所示,三种植物叶片光合指标在一天之中的变化趋势基本保持一致,一天中各时期净光合速率(Pn)、蒸腾速率(E)、气孔导度(Gs)和胞间CO2浓度(Ci)基本表现为蒲葵>槟榔>棕竹规律,局部有所波动;不同棕榈科植物叶片光合速率(Pn)日变化均表现为典型的双峰型曲线,第一峰出现在上午10:00左右,13:00光合速率出现峰谷,第二高峰出现在下午16:00前后,16:00以后光合速率明显下降,表现出明显的光合“午休”现象,在光合速率日变化曲线中;胞间CO2浓度(Ci)日变化随时间呈“W”字型变化规律,在上午9:00左右出现第一个波谷,13:00左右达到最大(波峰),13:00以后急剧下降,16:00左右出现峰谷,而后随时间的变化缓慢上升;气孔导度(Gs)日变化随时间呈“W”字型变化规律,其第一波峰出现在早上8:00左右,第二波峰出现在中午13:00左右,16:00左右出现第二次峰谷,而后随时间的变化缓慢上升;蒸腾速率(EVAP)日变化规律基本与光合速率(Pn)保持一致,均呈双峰曲线,局部峰值推迟或者提前。
2.4不同棕榈科植物叶片光合特性与生理响应
由表3可知,通过SPSS最佳曲线拟合(分别拟合光合速率Pn与PAR,CO2浓度和温度之间的曲线关系),筛选出R2最大(最佳拟合度及校准以后的最大R2)和p值(显著性)最小的曲线组合,并在此基础上检验其F值和差异显著性(p<0.001)。由表3可知,不同棕榈科植物的Pn-PAR变化呈一致的二次曲线关系。
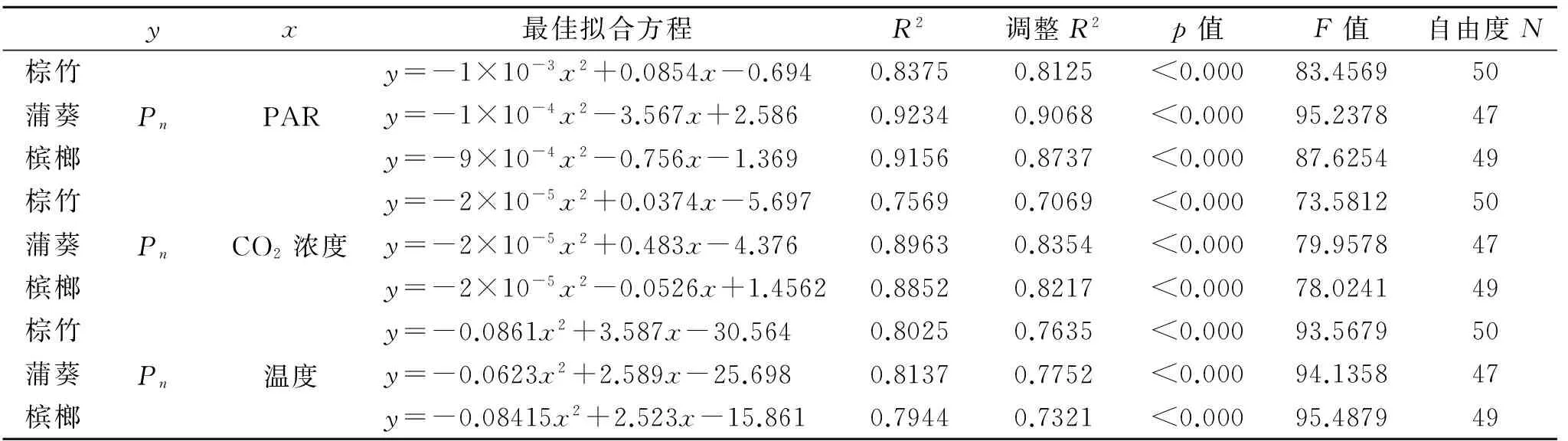
表3 不同棕榈科植物叶片光合特性与生理响应
3 讨论与结论
本研究中不同棕榈科植物株高、茎粗和冠幅均表现为槟榔>蒲葵>棕竹,叶面积指数和比叶重均表现为蒲葵>槟榔>棕竹(表1),不同棕榈科植物的生理代谢和形态建成的变化,导致叶面积指数和比叶重呈现出不同的变化规律[13-14]。叶面积指数是植物冠层生长状况的指标,叶面积指数较大有利于捕获更多的光能[15]。综合比较可知,蒲葵通过增加叶面积指数和光合作用进而影响其自身的生长;不同棕榈科植物根区土壤养分基本表现为蒲葵>槟榔>棕竹(表1),与叶面积指数、比叶重和光合等指标变化趋势相一致,而土壤pH则表现为棕竹>槟榔>蒲葵,由此表明不同棕榈科植物通过降低其根区土壤pH增加土壤养分含量,进而影响其光合等生理特性。由图1可知,不同棕榈科植物光饱和点(LSP)、光补偿点(LCP)、表观资产额(AQY)、羧化效率(CE)、气孔限制值(Ls)、水分利用效率(WUE)、光合氮利用效率(NUE)和光合磷利用效率(PUE)基本表现为蒲葵>槟榔>棕竹,局部有所波动,与叶面积指数和比叶重以及其根区土壤养分变化趋势相一致。综合比较可知蒲葵植物叶片的光能利用效率最高,表明蒲葵叶片对低CO2浓度和低光强的利用率最强,具有较强的再生速率。
一天各时期净光合速率(Pn)、蒸腾速率(EVAP)、气孔导度(Gs)和胞间CO2浓度(Ci)基本表现为蒲葵>槟榔>棕竹规律,局部有所波动,这与不同棕榈科植物光合生理等指标的变化趋势相一致,也间接表明了棕榈科植物光合作用受到各生理指标等影响较为显著。气孔是植物水分与叶片进行气体交换的通道和CO2进入细胞的门户,控制着叶片和大气之间的CO2和水分交换,气孔的闭合程度对光合与蒸腾作用会产生直接的影响[16]。由图2可知,不同棕榈科植物气孔导度(Gs)日变化与蒸腾速率(EVAP)曲线相似,说明不同棕榈科植物气孔的开闭程度直接影响蒸腾作用,主要是由水蒸气浓度差及水蒸汽扩散阻力所决定[17]。不同棕榈科植物EVAP与Gs变化趋势一致,主要是由于叶内外水蒸气压差的增大和水蒸气浓度减少,加剧了植物体蒸腾扩散阻力。纵观全天,气孔阻力和叶内外水蒸气压差交互主导或共同影响EVAP,不同时段以气孔运动因素的影响占优势,即植物自身的生理活动对EVAP起着主要的影响[17]。植物的光合特性反映了对外界环境的响应和适应,受到众多环境因素的影响,其中影响光合速率的因素主要是气孔限制和非气孔限制因素,即气孔的开张和叶肉细胞的光合活性[16,18]。Pn是光合作用强弱的重要指标,对不同棕榈科植物而言,Pn日变化表现为双峰曲线,有明显的“午休”现象,主要是由于中午持续的强光使气温、蒸腾速率急剧增加,导致光合速率与蒸腾速率日变化保持一致的变化趋势,而中午光合速率的最低值对应着光照度、温度的最高值和大气相对湿度的最低值,不同棕榈科植物叶片水分代谢失调,叶面温度过高而抑制了光合酶的合成及活性,Gs,Ci和空气中CO2浓度导致Pn下降,从而造成了不同棕榈科植物光合“午休”现象[14,16,18]。大量研究认为,中午Pn下降可能既有气孔因素也有非气孔因素,如果是气孔因素居主导地位,那么早在Pn和Gs下降的同时,Ci也应该下降,本研究的结果正好与此结论相符。由此说明,不同棕榈科植物光合午休现象主要是由气孔限制所决定的。
通过SPSS最佳曲线拟合(分别拟合光合速率Pn与PAR,CO2浓度和温度之间的曲线关系),不同棕榈科植物的Pn与PAR,CO2浓度和温度变化呈一致的二次曲线关系,由曲线计算可知不同棕榈科植物光合作用最适温度在23~25℃,这与前人关于园林树种的研究结论一致[19-20]。研究还发现,虽然不同棕榈科植物光合适温相差不大,但在最适温时的光合速率却相差较大,说明温度的变化对棕榈科植物光合作用有较大的影响。不同植物的光合作用对温度的响应不同,可能与物种的温度适应性(原产地的气候、环境)有关[14,16,18]。而相关性分析表明,不同棕榈科植物光合特性由多种因素共同决定,这些因素之间也会相互影响,其中Gs的相关系数较其他因子的相关系数绝对值大,这也间接说明了不同棕榈科植物净光合速率日变化(Pn)主要与气孔导度相关,午间净光合速率下降的原因既有气孔限制的调节,也有非气孔限制的调节,而影响棕榈科植物光合特性的主要因子仍然是生理指标,这与光合特性和生理特性的变化趋势相吻合。本研究中不同棕榈科植物光合生理指标属于瞬间测定的结果,受环境影响较大,所测定的结果有一定的偏差,而后续还需要进行反复大量的野外监测来验证本试验的研究结果。
[1]Corpas F J, Leterrier M, Valderrama R, et al. Nitric oxide imbalance provokes a nitrosative response in plants under abiotic stress[J]. Plant Science, 2011,181(5):604-611.
[2]Pell A, Márquez A, López-Sánchez J F, et al. Occurrence of arsenic species in algae and freshwater plants of an extreme arid region in northern Chile, the Loa River Basin[J]. Chemosphere, 2013,90(2):556-564.
[3]陈恒彬,周新月.常见棕榈科植物在园林绿化中的应用[J].亚热带植物通讯,1995,24(2):46-50.
[4]林秀香,陈振东.我国棕榈科植物的研究进展[J].热带作物学报,2008,28(3):115-119.
[5]卫兆芬.棕榈科植物的地理分布[J].热带亚热带植物学报,1995,3(2):1-18.
[6]谢国干,黄世满.海南岛棕榈科植物研究[J].热带地理,1994,14(1):63-72.
[7]黄志华,詹福麟.浅谈棕榈植物在园林景观设计中的应用[J].三明学院学报,2005,22(2):178-181.
[8]潘辉,李永莉,黄石德,等.棕榈科植物群落空气负离子密度影响因素[J].东北林业大学学报,2010,38(3):69-70.
[9]杨清,易国南,苏光荣,等.棕榈植物在园林中的应用及其造景艺术[J].西南林学院学报,2004,23(4):30-34.
[10]Craven D, Dent D, Braden D, et al. Seasonal variability of photosynthetic characteristics influences growth of eight tropical tree species at two sites with contrasting precipitation in Panama[J]. Forest Ecology and Management, 2011,261(10):1643-1653.
[11]Xu D, Li Y, Fan X, et al. Influence of earthworm Eisenia fetida on Iris pseudacorus's photosynthetic characteristics, evapotranspiration losses and purifying capacity in constructed wetland systems[J]. Water Science & Technology, 2013,68(2):335-341.
[12]展小云,于贵瑞,盛文萍,等.中国东部南北样带森林优势植物叶片的水分利用效率和氮素利用效率[J].应用生态学报,2012,23(3):587-594.
[13]王建华,任士福,史宝胜,等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报,2011,31(7):1811-1817.
[14]武辉,戴海芳,张巨松,等.棉花幼苗叶片光合特性对低温胁迫及恢复处理的响应[J].植物生态学报,2014,38(10):1124-1134.
[15]杨富军,赵长星,闫萌萌,等.栽培方式对夏直播花生叶片光合特性及产量的影响[J].应用生态学报,2013,24(3):747-752.
[16]Boccalandro H E, Giordano C V, Ploschuk E L, et al. Phototropins but not cryptochromes mediate the blue light-specific promotion of stomatal conductance, while both enhance photosynthesis and transpiration under full sunlight[J]. Plant Physiology, 2012,158(3):1475-1484.
[17]Lombardozzi D, Levis S, Bonan G, et al. Predicting photosynthesis and transpiration responses to ozone:decoupling modeled photosynthesis and stomatal conductance[J]. Biogeosciences, 2012,9(8):31-38.
[18]Schurgers G, Lagergren F, Mölder M, et al. The importance of micrometeorological variations for photosynthesis and transpiration in a boreal coniferous forest[J]. Biogeosciences, 2015,12(1):237-256.
[19]郎莹,汪明.春,夏季土壤水分对连翘光合作用的影响[J].生态学报,2015,35(9):3043-3051.
[20]梁文斌,聂东伶,吴思政,等.遮荫对短梗大参苗木光合作用及生长的影响[J].生态学杂志,2015,34(2):413-419.
Research on Photosynthetic Characteristics of Common Palmae Plants in Nanning City
GAO Chuanyou
(Guangxi Vocational and Technical College, Nanning 530226, China)
Photosynthetic characteristics, physiological property and the main influence factors of common Palmae plants (RhapisExcelsa,LivistonachinensisandArecacatechu) were investigated comparatively in Nanning City. The results showed that: (1) the plant heights, stem diameters and crown breadths of different Palmae plants decreased in the order:Arecacatechux>Livistonachinensis>Arecacatechu, leaf area indexes and specific leaf weights decreased in the order:Livistonachinensis>Arecacatechu>Rhapisexcelsa, pH levels of the root zone soil decreased in the order:Rhapisexcelsa>Arecacatechu>Rhapisexcelsa, and the nutrient contents of the root zone soil decreased in the order:Livistonachinensis>Arecacatechu>Rhapisexcelsawith local fluctuation, and there was no significant difference of soil organic carbon in common Palmae plants (p>0.05); (2) the light saturation point (LSP), light compensation point (LCP), apparent assets (AQY), carboxylation efficiency (CE), stomatal limitation (Ls), water use efficiency (WUE), nitrogen use efficiency (NUE) and phosphorus use efficiency (PUE) of three common Palmae plants decreased in the order:Livistonachinensis>Arecacatechu>Rhapisexcelsawith local fluctuation, which there was no significant difference of Ls of common Palmae plants (p>0.05), and this variation tendency was the same as the root zone soil nutrients; (3) the responsive curves of photosynthesis in three kinds of Palmae plants to the change of photosynthetic rate (Pn), transpiration rate (EVAP), stomatal conductance (Gs), intercellular CO2concentration (Ci) with theLivistonachinensiswas the highest and theRhapisexcelsawas the lowest, while stomatal limitation resulted in different levels of photosynthetic ‘lunch break’ phenomenon, and transpiration rate (EVAP), stomatal conductance (Gs), intercellular CO2concentration (Ci) of three kinds of Palmae plants showed the W-shape; (4) the change of photosynthetic active radiation (PAR), CO2concentration and air temperature of three kinds of Palmae plants could be described with quadratic equations (P<0.001); (5) correlation analysis showed that photosynthetic characteristics and physiological properties of Palmae plants were determined by a variety of environmental factors, and these factors could be influenced each other, and among these environmental factors, the absolute value of correlation coefficient ofGswas the greatest.
Nanning City; common Palmae plants; photosynthetic characteristics
2015-0-20
2015-09-25
广西高等学校科学技术研究“棕榈科植物园林景观营造及耐寒耐荫性研究”(2013LX183)
高传友(1984—),男,河南开封人,硕士,讲师,工程师,研究方向:园林植物生态、园林设计。Email:gao_chuanyou@126.com
S792
A
1005-3409(2016)02-0183-06
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
基层中医药(2021年11期)2021-06-05
新疆农业科学(2020年1期)2020-02-14
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
读书文摘(2017年10期)2017-10-16
民族音乐(2016年1期)2016-08-28
中国麻业科学(2015年5期)2015-12-28
中国铸造装备与技术(2015年5期)2015-12-10
