
小麦重组自交系群体苗期抗旱性综合评价
2016-10-10 03:12:58杜欣欣吴允朋秦思思苏荣存耿建芬
水土保持通报 2016年4期
杜欣欣, 张 红, 吴允朋, 秦思思, 苏荣存, 耿建芬
(德州学院 生态与园林建筑学院, 山东 德州 253023)
小麦重组自交系群体苗期抗旱性综合评价
杜欣欣, 张 红, 吴允朋, 秦思思, 苏荣存, 耿建芬
(德州学院 生态与园林建筑学院, 山东 德州 253023)
[目的] 探讨干旱对小麦萌发期幼苗生长的影响,筛选抗旱的优良小麦种质材料,为小麦抗旱新品种的选育提供依据。 [方法] 以洛旱2号与潍麦8号杂交产生的重组自交系(RIL)群体(F9)为材料,基于小麦萌发期幼苗在不同水分条件下的多个农艺性状指标数据,通过主成分分析和K均值聚类的方法,对重组自交系的291个株系及其亲本进行了苗期抗旱性综合评价,并对各株系的苗期抗旱性度量值和其产量的相关性进行了分析。 [结果] (1) 用10%的PEG-6000处理萌动的种子,可促进胚芽鞘长和根数的增加,而对其它农艺性状有明显的抑制作用。 (2) 从该重组自交系的291个株系中筛选出了44个抗旱优良株系,为小麦抗旱育种提供了宝贵的种质材料。 (3) 小麦各株系的苗期抗旱性和其产量之间呈显著的正相关。 [结论] 根系性状对干旱胁迫的反应最为敏感,干旱胁迫下能否形成相对强大的根系系统是衡量幼苗抗旱性的重要条件。
小麦重组自交系; 苗期; 抗旱性评价
文献参数: 杜欣欣, 张红, 吴允朋, 等.小麦重组自交系群体苗期抗旱性综合评价[J].水土保持通报,2016,36(4):286-290.DOI:10.13961/j.cnki.stbctb.2016.04.049
小麦(Triticumaestivum)是世界上第一大粮食作物,中国大部分麦区在其生育期间降水量不足,干旱是小麦生产持续稳定发展的最主要限制因子之一。为了降低干旱对小麦生产的影响,挖掘干旱半干旱地区小麦生产潜力,最大限度地增加小麦产量,通过培育抗旱节水高产新品种实现小麦生物节水的新突破,是当今小麦抗旱研究的重要课题。
小麦重组自交系是不同的小麦品种杂交后,再通过多代自交选育获得的性状稳定、基因型纯合的系统。重组自交系是两个亲本的聚合体,存在着优良的超亲变异。本研究将以不同水分条件下小麦种子苗期的多个农艺性状指标为依据,采用主成分分析和K均值聚类的方法,对由抗旱亲本和不抗旱亲本杂交获得的重组自交系进行苗期抗旱性综合评价,以期从中筛选出优良的抗旱材料,为小麦抗旱新品种的选育提供依据,同时探讨对大批量小麦材料抗旱性评价及筛选的可行途径,研究结果对于小麦的抗旱性育种具有重要的意义。
1 材料与方法
1.1材料
小麦抗旱亲本洛旱2号和非抗旱亲本潍麦8号及其杂交衍生的重组自交系(F9)群体,共291个株系,编号为001-302(中间有空缺)。
洛旱2号:由河南省洛阳市农科所1990年以洛阳78(111)矮做母本、晋麦33做父本杂交,系统选育而成,2003年通过国家审定,是一分蘖力较强,活秆成熟,耐旱,抗倒的半冬性中早熟小麦品种。
1.2试验方法
选100粒充实饱满的亲本和各株系的种子置于培养皿,置25 ℃培养室催芽1 d(24 h)。种子萌动后,各挑出60粒露白一致的种子分别均匀摆放在6个铺有两层滤纸(直径6 cm)的培养皿内,每皿10粒,其中3皿加5 ml 10%的PEG-6 000水溶液,另3皿加5 ml去离子水设为对照,培养皿放在塑料盒中,盖膜,25 ℃条件下培养,第4 d每皿补充5 ml去离子水,并开始补充光照。第8 d每皿选生长一致的5株,分别测定胚芽鞘长度(CL)、苗高(SH)、最大根长(RL)、根数(RN)、株鲜重(PFW)、根鲜重(RFW)、地上部鲜重(SFW)等性状。然后把幼苗放在烘箱里100 ℃杀青20 min,80 ℃烘干至恒重,称量株干重(PDW)、根干重、地上干重,计算根冠鲜重比和根冠干重比。
1.3材料的种植和产量测定
亲本和重组自交系群体分别在济宁市农科院和山东农业大学农学试验站(泰安)2个环境正常水分条件下种植,随机区组设计,顺序排列,3次重复,每个株系每次重复种植两行,每行长1 m,株距5 cm,收获时测1 m行长产量。
1.4数据的统计分析
数据的整理在Microsoft Excel中进行,首先,用SPSS软件进行两亲本和群体在两种水分条件下的方差分析和配对样本t检验,然后,根据抗旱系数计算出各株系性状指标的隶属函数值,再对隶属函数用SAS进行主成分分析,以第1主成分各性状的特征值作为各指标的权数与其隶属函数值相乘,分株系求和后,即得到抗旱性度量值(D值)[1],用SPSS软件对各株系的抗旱性度量值(D值)进行K均值聚类,再进行抗旱度量值和各性状抗旱指数及后期产量之间的相关性分析。性状指标的抗旱系数=(干旱胁迫下性状指标值/非胁迫下性状指标值)×100%[2]
1.4 统计学分析 应用SPSS 22.0统计软件进行统计分析。预后因素的单因素分析采用Kaplan-Meier法,多因素分析采用Cox比例风险模型多因素分析。将单因素分析显示与预后相关的因素再纳入多因素分析中。P<0.05为差异有统计学意义。
各性状指标隶属函数值[3]:
μ(x)=(X-Xmin)/(Xmax-Xmin)式中:μ(x)——某个性状指标抗旱系数的隶属函数值;x——某一性状指标抗旱系数;Xmax,Xmin——各参试材料中某一性状指标抗旱系数中的最大和最小值。
2 结果与分析
2.1干旱对小麦苗期性状指标的影响
t检验结果表明(表1),两种水分条件下,两亲本所有性状平均值都达到了差异极显著水平。在渗透胁迫条件下,潍麦8号和洛旱2号在胚芽鞘长度和根数上都有所增加,以洛旱2号的增加幅度更大;苗高、地上部鲜重、地上部干重洛旱2号和潍麦8号减少幅度基本一致;株鲜重、株干重、根鲜重、根干重潍麦8号的减少幅度大于洛旱2号,而在根长、根冠鲜重比和干重比3个性状上,洛旱2号表现为增加,潍麦8号表现降低。在正常水分条件下,洛旱2号的胚芽鞘长度和最大根长都低于潍麦8号,胁迫条件下,洛旱2号胚芽鞘长度增加幅度较大,与潍麦8号基本一致,而最大根长明显超过潍麦8号。在正常水分条件下,在根冠鲜重和干重比两个性状上,潍麦8号和洛旱2号差异不显著,胁迫条件下洛旱2号在这2个性状上明显增加,而潍麦8号明显降低。表明耐旱品种在干旱胁迫条件下有较强根系形成能力,可以促进对水分的吸收,以减少干旱对生长的影响。
二因素方差分析表明,株系间和不同水分处理间各性状值均差异显著,在两种水分条件下,重组自交系群体所测性状平均值除胚芽鞘达差异显著外(p<0.05),其他性状平均值差异都达极显著水平(p<0.01)。渗透胁迫条件下,除胚芽鞘长度、根数较正常水分条件增加外,其余性状均表现下降,进一步表明渗透胁迫在一定程度上抑制了小麦幼苗的生长,但对小麦苗期根数的增加和胚芽鞘的伸长有一定的促进作用。
正常水分条件下,群体均值除根数高于亲本均值,胚芽鞘低于亲本均值外,其他性状都介于双亲均值之间。胁迫条件下,群体均值除苗高、地上部鲜重高于亲本均值,胚芽鞘、最大根长低于双亲均值外,其他性状介于双亲之间。两种水分条件下,群体所有性状的最大值都大于高值亲本,而最小值都低于低值亲本,表现为超亲分离,变异范围广泛。这表明RIL群体具有比较丰富的遗传变异,选择潜力较大。
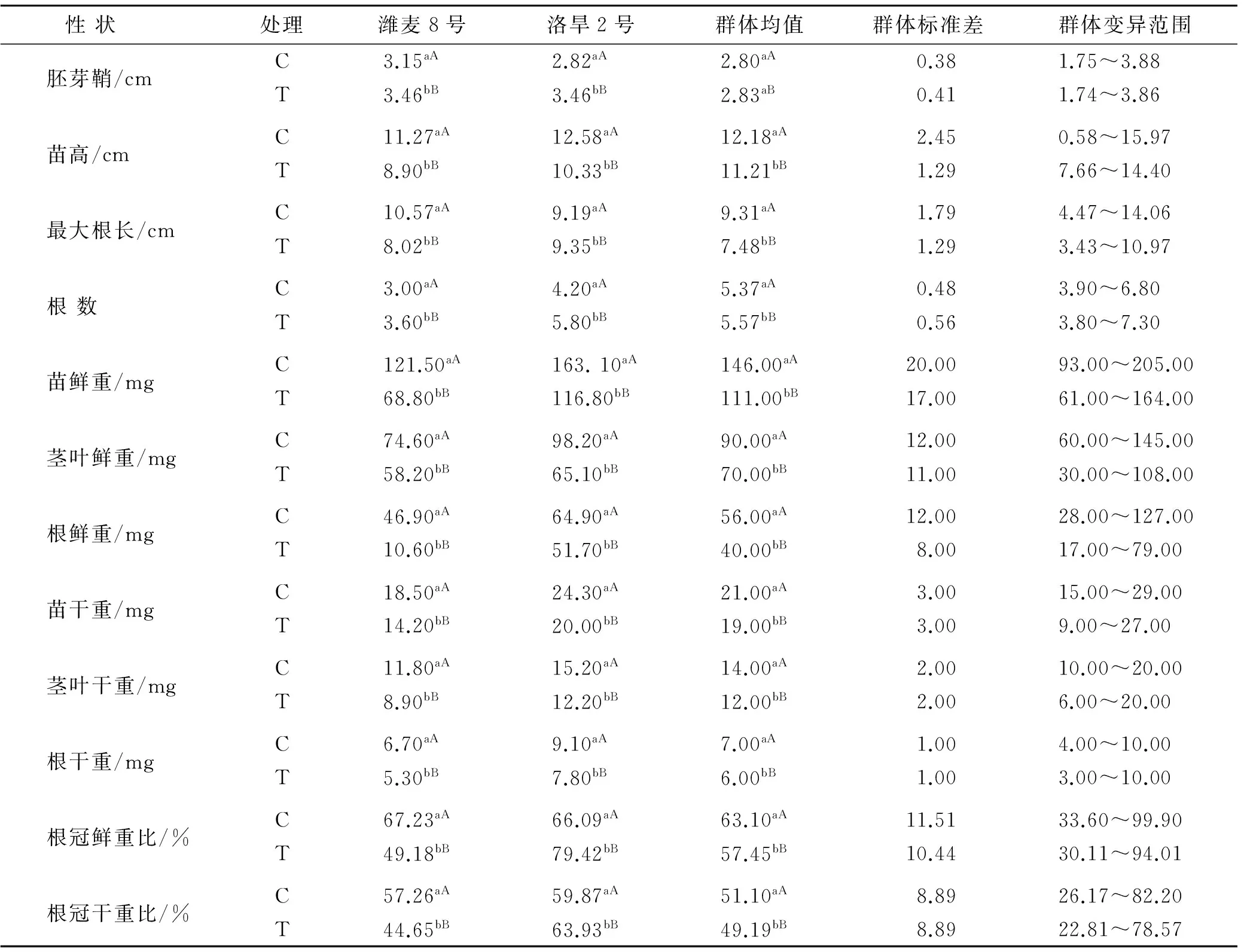
表1 两种水分条件下RIL群体及其亲本的苗期性状值
注:小写字母表示差异显著性水平为p<0.05; 大写字母表示差异显著性水平为p<0.01。
2.2主成分分析
以RIL群体各株系及其亲本苗期抗旱相关性状的抗旱系数为基础,利用SAS软件计算出各主成分的特征向量和贡献率(表2),并根据各向量的绝对值将不同性状指标划分到不同的主成分之中。同一指标在各因子中的最大绝对值所在位置即为其所属主成分。从表2可以看出,主成分分析特征值中4个成分的累计贡献率达到86.19%,而理论上只需85%以上的累计贡献率即可认为其具有较强的信息代表性。因此可以用这几个主成分对其抗旱性进行概括分析,提取的4个主成分基本上代表了12个原始指标的绝大部分信息。
决定第1主成分大小的主要是苗鲜重、苗干重、根鲜重3个性状分量,主成分1相当于6.919个原始指标的作用,它可反映原始数据信息量的40.8%。这几个性状均属于苗的生物产量性状,干旱胁迫下苗生长量越大,说明苗的抗旱性越强。因此,可把第1主成分称为“幼苗生物产量因子”。
决定第2主成分大小的主要是茎叶鲜重、茎叶干重、根冠干重比3个性状分量。主成分2相当于4.875个原始指标的作用,可单独说明整个原始数据标准差异的28.76%。茎叶干、鲜重的抗旱系数越大,根冠干重比的抗旱系数越小,则第2主成分越大,因此,可把第2主成分称为“茎叶生物产量因子”。决定第3主成分大小的主要是根干重1个性状分量,根干重的抗旱系数越大,则第3主成分越大。第3主成分相当于1.652个原始指标的作用,可反映全部信息量的9.75%。根干重反映了植系的生长状况,因此可把第3主成分称为“根系生物产量因子”。
第4主成分的大小主要由胚芽鞘长、苗高、最大根长、根数、根冠鲜重比等决定。第4主成分相当于1.167个原始指标的作用,可反映全部信息量的6.88%。几个性状都与幼苗的生长有关,因此可统称为“生长因子”
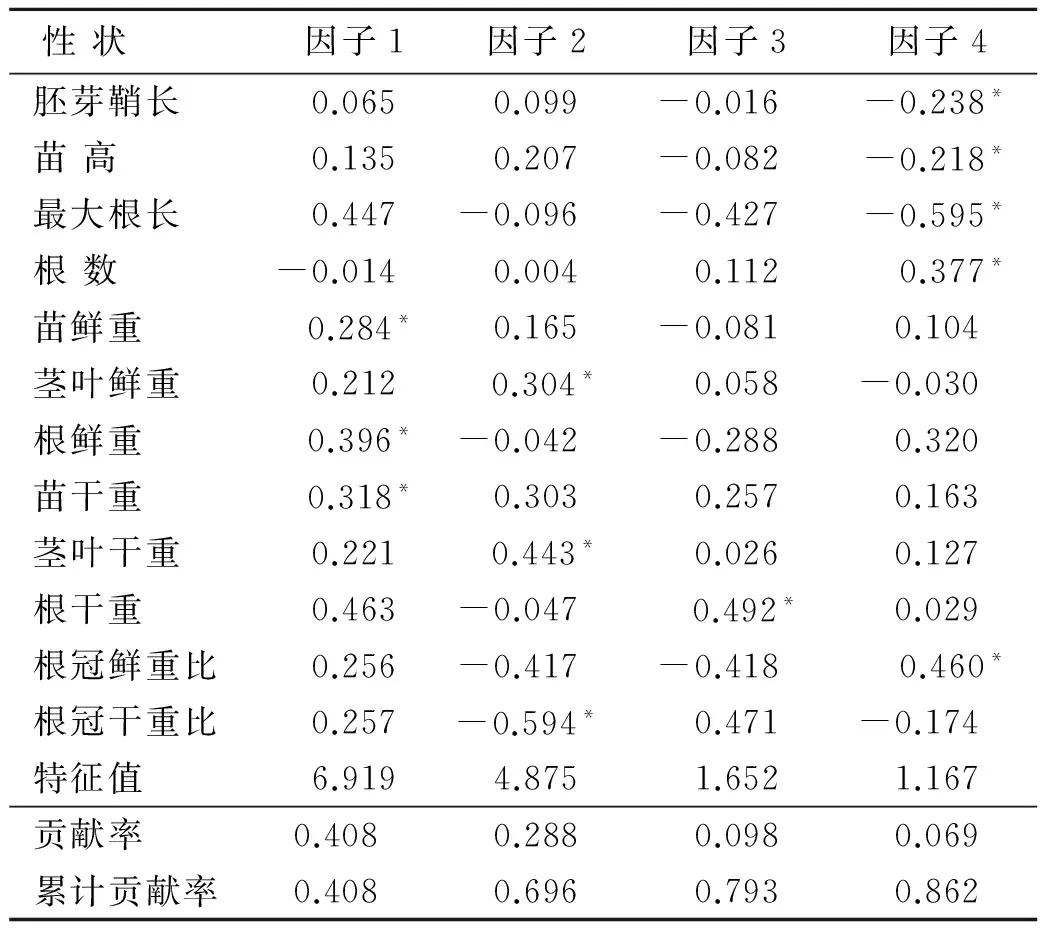
表2 各性状主成分的特征向量及贡献率
注: *表示某指标在各因子中的最大绝对值。
2.3抗旱性评价
根据抗旱性度量值的大小可以推断抗旱性的强弱。本RIL群体和亲本的抗旱度量值的变异范围为0.632~2.220,潍麦8号和洛旱2号的抗旱度量值分别为0.714,1.385。用SPSS软件根据抗旱度量值进行K均值聚类,可将291个株系和两个亲本的抗旱性划分为3类。不抗旱(D值从0.29~1.11)、中度抗旱(D值从1.12~1.48)、高度抗旱(D值从1.49~2.22),亲本潍麦8号和101个株系属于不抗旱类型;洛旱2号和102个株系属于中度抗旱类型;44个株系属于高度抗旱类型。由抗旱性评价结果可以看出,洛旱2号的苗期抗旱性要优于潍麦8号,这与生产实际相符,虽然潍麦8号不抗旱,但两亲本杂交后产生部分超亲变异,有15%的株系的苗期抗旱性优于洛旱2号。这些株系的选出为小麦抗旱育种提供了宝贵的种质材料。
2.4抗旱性度量值和产量的相关性分析
抗旱性度量值和1 m行长产量性状之间的相关性分析表明,抗旱性和2个环境下的平均1 m行长产量之间呈显著正相关,相关系数为0.151。另外,比较抗旱性不同的3类小麦株系,发现所有高度抗旱株系的1 m行长产量的平均值为171.48 g,所有中度抗旱株系的1 m行长产量的平均值为161.60 g,所有不抗旱株系的1 m行长产量的平均值为159.19 g,高度抗旱株系中1 m行长产量大于200 g的占所有抗旱株系的21%,中度抗旱株系中产量大于200 g的占所有中度抗旱株系的8.5%,不抗旱株系中1 m行长产量大于200 g的占所有不抗旱株系的7.2%。由此可见,苗期抗旱性强的株系一般适应环境的能力较强,高产稳产的潜力较大,可以通过苗期抗旱性选择对产量性状进行早期的选择。
3 讨论分析
3.1抗旱性分析方法
抗旱性分析可采用单指标分析和多指标分析。单指标分析是指采用与抗旱性高度相关的性状指标,如抗旱指数[4]、反复干旱后幼苗的存活率[5]等。这类指标虽然简便、快捷,但却只能反映某一性状对干旱胁迫的反应,而多指标评价可以从不同侧面反映作物抗旱性的强弱和特点。抗旱性的多指标分析时,常采用的有平均隶属函数法[6-8]和主成分分析法[9-11],平均隶属函数法是求出各性状的隶属函数以后求平均值,由于各性状受干旱影响的程度是不一样的,在干旱性评价时的重要性也不同,故用此法对小麦抗旱性进行评价存在一定的局限性。常用的主成分分析法是指根据各性状的隶属函数值提取总贡献率大于85%的几个主成分,用各主成分得分值乘以各主成分的加权系数,求和既可得到一个抗逆性的综合评价值。主成分分析可以去粗取精,给每一个性状指标确立一个合理的加权系数,从而可较科学地对各作物品种的抗逆性进行评价。孟生旺[1]、孟庆立[12]指出在主成分分析时应以第1主成份的值作为综合评价值,而不能以综合主成份的值作为综合评价值。本研究对小麦苗期12个性状的隶属函数进行主成分分析,从中提取了4个主成分,总贡献率为86.19%,其中第1主成分的贡献率为40.8%,以第1主成分值作为综合评价值对各株系的抗旱性进行了评价,以K均值聚类方法把291个株系和2个亲本分为3类。整个分析过程都遵循统计学的原理,排除了人为因素的干扰,使分析结果更加客观可靠。
3.2小麦的苗期抗旱性
与小麦苗期抗旱性有关性状的研究主要集中在胚芽鞘和根系性状等。胚芽鞘是小麦种子萌发时的保护组织,在生长初期对幼苗的正常出土和苗全苗壮具有重要的作用[13-15]。王玮等[16]和邹琦等[17]研究发现,在低水势下胚芽鞘长度和叶片渗透能力与产量有密切的关系,在干旱胁迫下小麦胚芽鞘长度与抗旱系数呈极显著正相关;尤其是在20%PEG-6000处理下,抗旱性强的品种胚芽鞘生长得快,从而提出了“低水势下胚芽鞘长度法”用于冬小麦抗旱性鉴定。本研究发现洛旱2号在水分胁迫下胚芽鞘的生长速度远远大于潍麦8号的生长速度,进一步验证了干旱对不同品种胚芽鞘的影响。另外研究发现虽然胚芽鞘的抗旱系数与抗旱性度量值之间呈极显著正相关,但相关系数却只有0.26,远远低于除根数以外的其他性状指标,这也说明只用胚芽鞘作为小麦苗期抗旱性的鉴定指标是否合理还有待探讨。
幼苗根系是最早感受土壤干旱的器官,对小麦苗期耐旱能力有重要作用。关于根系性状与抗旱性的关系,前人已做出很多有益的探索。杨国航等[18]发现玉米在干旱胁迫情况下,有较强生根发芽能力的品种,幼根萌发早,抗旱指数较大,抗旱性较强。郝树荣等[19]对水稻根系做了研究,认为干旱胁迫诱导根系变长,以利于吸收更深层土壤的水分以满足作物生长要求,胁迫越重根系越长。景蕊莲等[20]研究根系形态性状的遗传力及其与抗旱性的关系,结果表明小麦幼苗的抗旱性与根干重呈显著正相关。本研究发现在10%PEG-6000处理下,除了根数相对于正常水分条件下略有增加外,根数、根干重、根鲜重等性状指标都表现为减小,这和郝树荣等的研究结果不甚相同,作者认为干旱胁迫对作物根系的生长有比较明显的抑制作用,只是抗旱品种和不抗旱品种相比受到的抑制较小,因此能形成相对强大的根系系统,来补偿干旱对植物生长造成的影响。本研究还发现根长、根鲜重、根干重和抗旱度量值之间都呈极显著正相关,且相关系数在所有所测的性状指标中是最高的,根系性状是受干旱影响最敏感的性状,也是苗期抗旱性鉴定的重要指标性状,这与前人的研究结果是一致的。
苗期抗旱性研究虽然取得了一些进展。但苗期抗旱性和产量性状之间的关系研究较少,本研究利用RIL群体把苗期抗旱度量值和两个环境正常水分管理条件下的产量性状平均值进行了相关性分析,发现苗期抗旱性和产量之间呈显著正相关,虽然由于条件限制,没有对各株系干旱处理条件下的产量性状与苗期抗旱性间的关系进行研究,但从本研究仍可以看出,各株系的苗期抗旱性在一定程度上反映了该株系对环境的适应性,一般苗期抗旱性强的株系高产稳产的潜力也比较大,因此,在小麦的抗旱高产育种中对大批量的材料可以通过本试验提供的方法进行初步的筛选,从而减少工作量,提高选择效果。
4 结 论
小麦苗期用10%的PEG-6000处理种子,可促进胚芽鞘长和根数的增加,而对其它农艺性状有明显的抑制作用;以第1主成分求得的抗旱度量值为依据,通过K均值聚类,从291个重组自交系群体中筛选出44个抗旱优良株系,为小麦抗旱育种提供了宝贵的种质材料;通过抗旱度量值和各性状的抗旱系数之间的相关性分析,表明根系性状对干旱的反应最为敏感,干旱胁迫下能否形成相对强大的根系系统是衡量苗期抗旱性的重要条件;小麦苗期抗旱性和产量之间呈显著的正相关。
[1]孟生旺.用主成分分析法进行多指标综合评价应注意的问题[J].统计研究,1992(4):67-68.
[2]Bouslama M, Schapaugh W T. Stress tolerance in soybeans(I): Evaluation of three screening techniques for heat and drought tolerance[J]. Crop Science, 1984,24(5):933-937.
[3]何雪银,文仁来,吴翠荣,等.模糊隶属函数法对玉米苗期抗旱性的分析[J].西南农业学报,2008,21(1):52-56.
[4]兰巨生,胡福顺,张景瑞.作物抗旱指数的概念和统计方法[J].华北农学报,1990,5(2):20-25.
[5]李荫梅.谷子(粟)品种资源抗旱性鉴定研究[J].华北农学报,1991,6(3):20-25.
[6]刘莹,盖钧镒,吕慧能,等.大豆耐旱种质鉴定和相关根系性状的遗传与QTL定位[J].遗传学报,2005,32(8):855-863.
[7]王育红,姚宇卿,吕军杰.花生抗旱性与生理生态指标关系的研究[J].杂粮作物,2002,22(3):147-149.
[8]石书兵,徐文修,张强,等.旱作春小麦品种高产抗旱特性的综合评价[J].干旱地区农业研究,2001,19(2):14-20.
[9]白志英,李存东,孙红春,等.小麦代换系抗旱生理指标的主成分分析及综合评价[J].中国农业科学,2008,41(12):4264-4272.
[10]周广生,梅方竹,周竹青,等.小麦不同品种耐湿性生理指标综合评价及其预测[J].中国农业科学,2003,36(11):1378-1382.
[11]王军,周美学,许如根,等.大麦耐湿性鉴定指标和评价方法研究[J].中国农业科学,2007,40(10):2145-2152.
[12]孟庆立,关周博,冯佰利,等.谷子抗旱相关性状的主成分与模糊聚类分析[J].中国农业科学,2009,42(8):2667-2675.
[13]胡颂平,杨华,邹桂花,等. 水稻胚芽鞘长度与抗旱性的关系及QTL定位[J].中国水稻科学,2006,20(1):19-24.
[14]Cosgrove D J, Li ZhenchangC. Role of expansion in cell enlargement of oat coleoptiles[J]. Plant Physiol, 1993,103(4):1321-1328.
[15]Trethowan R M, Singh R P, Espino J H, et al. Coleoptile length variation of near-isogenic Rht lines of modern CIMMYT bread and durum wheats[J]. Field Crops Research, 2001,70 (3):167-176.
[16]王玮,邹琦,杨兴洪,等.低水势下冬小麦胚芽鞘长度与叶片的渗透调节能力及大田产量关系的研究[J].植物学通报,1997,14(S):55-59.
[17]邹琦,王玮,杨兴洪,等.冬小麦抗旱性鉴定的新方法:低水势下胚芽鞘长度法[J].中国农学通报,2000,16(15):23-25.
[18]Yang Guohang, Wang Weihong, Song Huixin, et al. Analysis of drought resistance of maize hybrids in rain-fed farming[J].Crops, 2009(5):78-81.
[19]郝树荣,郭相平,王为木,等.水稻分蘖期水分胁迫及复水对根系生长的影响[J].干旱地区农业研究,2007,25(1):149-152.
[20]景蕊莲,胡荣海,朱志华,等.冬小麦不同基因型幼苗形态性状遗传力和抗旱性的研究[J].西北植物学报,1997,17(2):152-157.
Comprehensive Evaluation of Drought Tolerance at Seedling Stage for Wheat Recombinant Inbred Lines
DU Xinxin, ZHANG Hong, WU Yunpeng, QIN Sisi, SU Rongcun, GENG Jianfen
(CollegeofEcologyandGardenArchitecture,DezhouUniversity,Dezhou,Shandong253023,China)
[Objective] The effects of drought on wheat seedling growth and selecting excellent wheat germplasm materials with drought resistance were studied to provide a basis for the breeding of new drought-resistive varieties. [Methods] Seedlings derived from the cross between Weimai 8 and Luohan 2 were used as experimental materials, which had 291 lines from a recombination inbred line(RIL) population(F9). The seedlings were treated with different water conditions during germination stage, and multiple agronomic trait indices were measured. A comprehensive evaluation of drought tolerance of the RIL population and its parents were conducted by methods of principal component analysis andK-means clustering. And also, correlation analysis between drought tolerance indices and yields of all the lines was conducted. [Results] Those germinated wheat seeds treated with 10% PEG-6000 were observed to be promoted for coleoptile length and root number, while obvious inhibitory effects were observed for other agronomic traits. 44 excellent lines with drought tolerance were selected from the RIL populations, which provided valuable germplasm materials for drought resistive wheat breeding. [Conclusion] The responses of root to drought stress performed the mostly sensitive, because a strong root system under drought tolerance is one of the important conditions that decide the drought tolerance of seedling. Drought tolerance of wheat seedlings and its yield were remarkably positively correlated.
wheat recombination inbred line, comprehensive evaluation; drought tolerance; seedling stage
2015-03-19
2015-10-04
国家高技术研究发展计划(863)子课题“分子染色体工程高效育种技术体系建立和黄淮冬麦区新品种选育”(2011AA100103); 2014德州学院校级科技人才引进计划项目(311885)
杜欣欣(1994—),女(汉族),山东省东营市人,本科生,研究方向为植物遗传育种。E-mail:1172241886@qq.com。
张红(1971—),女(汉族),山东省平原县人,博士,讲师,主要从事作物遗传育种研究。E-mail:zhw718_0@163.com。
A
1000-288X(2016)04-0286-05
S33
猜你喜欢
中学生物学(2022年8期)2022-10-13 09:55:02
亚热带植物科学(2022年1期)2022-05-17 12:39:32
生物学通报(2020年10期)2020-08-13 08:52:34
四川农业科技(2019年5期)2019-07-01 09:46:46
浙江农业学报(2017年1期)2017-05-17 06:13:36
教学考试(高考生物)(2016年2期)2016-08-11 13:10:07
西南农业学报(2016年4期)2016-05-17 05:42:13
浙江柑橘(2016年1期)2016-03-11 20:12:31
山西农经(2015年7期)2015-07-10 15:00:57
中国烟草学报(2012年4期)2012-04-09 07:11:42
