
Functions of PARylation in DNA Damage Repair Pathways
2016-09-28 08:02:04HuitingWeiXiaochunYu
Huiting WeiXiaochun Yu*b
1Department of Immunology,Tianjin Key Laboratory of Cellular and Molecular Immunology,MOE Key Laboratory of Immune Microenvironment and Disease,School of Basic Medical Sciences,Tianjin Medical University,Tianjin 300070,China
2Department of Cancer Genetics and Epigenetics,Beckman Research Institute,City of Hope Medical Center,Duarte,CA 91010,USA
REVIEW
Functions of PARylation in DNA Damage Repair Pathways
Huiting Wei1,a,Xiaochun Yu2,*,b
1Department of Immunology,Tianjin Key Laboratory of Cellular and Molecular Immunology,MOE Key Laboratory of Immune Microenvironment and Disease,School of Basic Medical Sciences,Tianjin Medical University,Tianjin 300070,China
2Department of Cancer Genetics and Epigenetics,Beckman Research Institute,City of Hope Medical Center,Duarte,CA 91010,USA
Available online 27 May 2016
Handled by Zhao-Qi Wang
KEYWORDS
Poly ADP-ribosylation;
PARPs;
DNA damage response;
PAR-binding modules;
Ubiquitination
AbstractProtein poly ADP-ribosylation(PARylation)is a widespread post-translational modification at DNA lesions,which is catalyzed by poly(ADP-ribose)polymerases(PARPs).This modification regulates a number of biological processes including chromatin reorganization,DNA damage response(DDR),transcriptional regulation,apoptosis,and mitosis.PARP1,functioning as a DNA damage sensor,can be activated by DNA lesions,forming PAR chains that serve as a docking platform for DNA repair factors with high biochemical complexity.Here,we highlight molecular insights into PARylation recognition,the expanding role of PARylation in DDR pathways,and the functional interaction between PARylation and ubiquitination,which will offer us a better understanding of the biological roles of this unique post-translational modification.
Introduction
Throughout the biological life,genomic stability of the organisms is always challenged by both endogenous and exogenous toxic stresses[1,2].It has been estimated that every cell could experience up to 105spontaneous DNA lesions per day[3]. To maintain genomic integrity,the organisms have evolved a series of sophisticated and precise mechanisms to protect their genome against the deleterious lesions,including cell cycle checkpoint,diverse DNA repair signaling pathways,chromatin reorganization,and protein modifications[4].Among these responses,poly ADP-ribosylation(PARylation)is a pivotal post-translational protein modification(PTM)that appears rapidly at DNA damage sites[5,6].
In human,ADP-ribosylation is catalyzed by poly(ADP-ribose)polymerases(PARPs),which consists of 17 members[7-10].PARPs primarily covalently attach the ADP-ribose(ADPR)unit via an ester bond to the carboxyl group of acidic residues such as glutamate or aspartate residues on the target proteins[11,12],but cysteine(Cys)and lysine(Lys)residues could also act as acceptors[13,14].However,most of them are only able to transfer single mono(ADP-ribose)(MAR)group onto their target proteins[5,15].To date,PARP1,2,and 3 have been identified to catalyze PARylation during DNA damage response(DDR)[5,15].In addition,tankyrases including tankyrase-1(PARP5a)and tankyrase-2(PARP5b)have also been shown to contribute to genomic stability[15,16].Among these PARPs,PARP1 is the founding member of PARP family for the synthesis of PAR chains.The mechanism of PARP1 activation by single-strand and double-strand DNA breaks(SSBs and DSBs)is well established[17].Using NAD+as substrate,PARPs repeatedly catalyze the transfer of successive units of ADPR moieties via a unique 2′,1′′-O-glycosidic ribose-ribose bond to target proteins,finally producing PAR chain[5].Several reports have demonstrated that PAR chains can comprise up to 200 ADPR units in length[5,11,17].In addition,PARP1 can introduce branching into PAR chains through the 2′′,1′′′-glycosidic bond[18,19].
In cells,PAR polymers are primarily degraded by PAR glycohydrolase(PARG),which possesses both exoglycosidic activity and endoglycosidic activity[20-22].PARG efficientlycleavestheunique2′,1′′-glycosidicribose-ribose bonds of the PAR chains and releases the free ADPR moieties[22,23].In addition,ADP-ribosylhydrolase 3(ARH3)also exhibits the PAR-degrading activity,although ARH3 has only exoglycosidase activity[24,25].Neither PARG norARH3canhydrolyzetheproximalprotein-bound ADPR unit from a PAR chain,possibly due to steric hindrance,thus leaving a MARylated protein.MARylated proteins can be recognized by different protein domain and thus serve as scaffolds for recruitment of proteins during diverse biological processes[22,24].Interestingly,a set of Macro domain-containing(MacroD)proteins have been found to exhibit hydrolase activities.These include the terminal ADPR protein glycohydrolase(TARG1/C6orf130)[26],as well as MacroD1 and MacroD2[27-30].Earlier studies showed that these three enzymes can hydrolyze O-acetyl-ADPR,a metabolite derived from NAD during sirtuin2-catalyzed protein deacetylation,regulating diverse biological processes[31].Recently,TARG,MacroD1,and MacroD2 were identified for their action in removal of glutamatespecific ADPR[26,28,29].The hydrolysis of last ADPR from modified protein is the final and rate-limiting step of PAR chain degradation[32].Like many other PTMs,synthesis and degradation of PAR chains is tightly and dynamically controlled in vivo with the half-life of only several minutes[4,27].If PAR chains cannot be hydrolyzed in a timely manner,excessive protein-free PAR chains can induce the apoptosis-like cell death,termed parthanatos[4,27].Parthanatos is another form of programed cell death which is distinct from necrosis and apoptosis.As a distinct death pathway,parthanatos is associated with PARP-1.The synthesis and accumulation of PAR chain will result in mitochondrial depolarization and nuclear apoptosis inducing factor(AIF)translocation,thus inducing cell death[33].
PARylation and DNA repair pathway
PARylation in base excision repair/SSB repair process
The base excision repair/SSB repair process(BER/SSBR)is a pivotal DNA repair signal pathway to repair oxidized bases,apurinic/apyrimidinic sites(AP sites,also known as abasic sites)or SSBs[1].In cells,many chemical alterations such as oxidation,methylation,deamination,and hydroxylation can induce base damage and SSBs[1].In the BER process,damaged bases are cleaved by DNA glycosylases,producing abasic sites,which are next processed by AP endonuclease(APE)into SSBs[2,34].These sites are further repaired through two different pathways termed short-patch repair and long-patch repair,which are distinct in terms of patch sizes and DNA repair factors involved[35].
PARP1 can physically and functionally interact with SSBR factor X-ray repair cross-complementing protein 1(XRCC1),which plays a major role in SSBR signal pathway,facilitating the recruitment and assembly of the SSBR machinery[35]. OurrecentstudyindicatesthattheBRCA1CTerminus(BRCT)domain of XRCC1 directly binds to PAR chain and mediates early recruitment of XRCC1 to DNA lesions[36].Several reports have also demonstrated that PARP1 is able to interact with key factors of the BER/SSBR process including the DNA glycosylase 8-oxoguanine glycosylase 1(OGG1),XRCC1,DNA polymerase(DNAP)β,DNA ligase III,proliferating cell nuclear antigen(PCNA),aprataxin,and condensin I[37-40]. Many of these factors can undergo PARylation by PARP1(Figure 1).Additionally,PARP2 has also been identified to interact with BER/SSBR proteins such as XRCC1,DNAP β,andDNAligaseIII[41].Thesefindingssupport thatPARchain could provide a landing platform for the recruitment of DNA repair complexes as proposed by Masson et al.in 1998[42].
PARylation in DSB repair
DNADSBsareregardedasthemostdetrimentalDNAdamage,whichseriouslyanddirectlythreatengenomicstabilityviainterrupting the physical continuity of the chromosome[1].The failure to repair DSBs will lead to catastrophic consequences such as oncogenesis,cell death,and developmental disorders[1].To deal with DSBs,organisms have employed three major DNA repair mechanisms including classical non-homologous end joining(C-NHEJ),alternative non-homologous end joining(alt-NHEJ),and homologous recombination(HR).The choice ofDNArepairpathwaydependsonwhetherthedamagedDNA end is resected,which is likely mediated by the Mre11/Rad50/ Nbs1(MRN)complexandC-terminal-bindingprotein(CtBP)-interacting protein(CtIP).Once DNA resection is impeded,repair by C-NHEJ is invoked.However,if resection has occurred,HR and alt-NHEJ may compete with each other torepairthedamagedDNA.RAD51formsafilamentatthesite of SSB that drives strand exchange and facilities HR,whereas PARP1 may serve as a platform for recruiting alt-NHEJ repair factors such as DNAP θ[43].
PARylation in C-NHEJ
Eukaryocytes mainly employ C-NHEJ to repair damaged DNA.The process is DNA end resection-independent,and is also unrelated to sequence homology.Therefore,C-NHEJ occurs throughout the cell cycle,but predominantly in G0/G1 and G2 phase[44,45].In the process of C-NHEJ,the Ku70/Ku80 heterodimer is recruited to DNA damage sites followed by loading of DNA-dependent protein kinase catalytic subunit(DNA-PKcs).Meanwhile,Ku70/Ku80 heterodimer facilitates the activation of the DNA ligase IV/XRCC4 complex.AccessoryfactorssuchasnucleaseArtemis,aprataxin-polynucleotidekinase-likefactor(APLF),or polynucleotide kinase/phosphatase(PNKP)process the damaged DNA end to be compatible for ligation.At the final step,the activated DNA ligase IV and its cofactor XRCC4,or Cernunnos/XRCC4-like factor(XLF),rejoin the DNA ends[46].
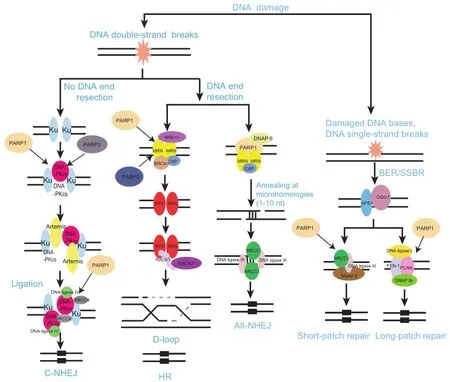
Figure 1 PARylation mediates DNA damage repair
Several studies support an important role of PARP1 in CNHEJ.Interaction between PARP1 and DNA-PKcs facilitates genomic integrity during V(D)J recombination and prevents tumor development[47].It is of note that PARP can stimulate DNA-PKcs activity via PARylation in vitro[48].This interaction is further supported by in vivo evidence as reported recently.A structural PARP1/DNA-PKcs/Ku molecular complex has been identified in which PARP1 elicits a major architectural rearrangement of the DNA-PKcs-mediated synapsis[49].Moreover,previous studies from our lab have shown that the BRCT domain of DNA ligase IV directly recognizes the ADP-ribose of PAR chains,which mediates the early recruitment of the ligase to DNA lesions.Such efficient recruitment may facilitate C-NHEJ[50].
PARylation in alt-NHEJ
As a new DSB repair signal pathway,alt-NHEJ has attracted much attention recently[46].When classical C-NHEJ is deficient,alt-NHEJ can be initiated by resected DNA end.Compared with C-NHEJ,alt-NHEJ is characterized by the following features:initiated by damaged DNA end resection;independent of the Ku70/Ku80 heterodimer,XRCC4,and DNA ligase IV;using complementary microhomologies—short stretches(1-10 nucleotides)that can anneal,to guide DNA repair and much less faithful than C-NHEJ[51].PARP-1,XRCC1,DNA ligase III,PNKP,WRN,CtIP,NBS1,and ERCC1 have all been implicated in alt-NHEJ[46].PARPs play pivotal roles in this process.PARP1 can recognize the broken DNA ends and create a scaffold for the recruitment of other DNA damage factors involved in alt-NHEJ.Finally,end-rejoining is carried out by the DNA ligase III/XRCC1 complex in coordination with PARP1[52](Figure 1).In addition,both XRCC1 and PNKP can be recruited to the DNA damage sites through PAR binding,which could occur at the early steps of alt-NHEJ[50].
PARylation in HR
HR can be activated by single-stranded DNA(ssDNA)resection.The process produces a lagging strand gap or 3′overhang,which is the key step for HR[53].Owing to its requirement for a sister chromatid,HR predominates in S and G2 phases,when the amount of DNA replication is highest and the sister template is available[45,54].HR is typically characterized by error-free[1,55].Using homologous sequence to repair damaged DNA,HR requires strand invasion mediated by the recombinase RAD51.Earlier findings show that PARP1 is dispensable for HR.PARylation appears to have little direct effect on HR since HR is normal in PARP-depleted cells[53].However,PARP1 has been associated with HR-mediated repair and reactivation of stalled replication forks,therefore promoting faithful DNA replication[56].Moreover,PARP1 facilitates recruitment of MRE11 and RAD51,which restart stalled replication BRCA1/2-dependent early DDR[57].The BRCTs of BARD1,theoligonucleotide/oligosaccharidebinding-fold(OB-fold)of BRCA2,and the protein incorporated later into tight junctions(PilT)N terminus(PIN)domain of exonuclease 1(EXO1)are the PAR-binding modules that target these HR repair machineries to DSBs for damaged DNA repair[58].
PAR-binding modules
To regulate numerous biological functions,PAR chains must be recognized by diverse proteins such as DDR factors.To date,several distinct classes of PAR-binding modules have been identified.These include the PAR-binding zinc finger(PBZ),the Macro domain,the WWE domain,the BRCT domain,the forkhead-associated(FHA)domain,the OB-fold domain,the PIN domain,and the RNA recognition motif(RRM)domain[9,59].
PBZ domain
The recently-identified PBZ domains possess the consensus sequence[K/R]xxCx[F/Y]GxxCxbbxxxxHxxx[F/Y]xH[60]. PBZs are less common in mammalian proteins involved in DNA repair and cell cycle checkpoint,although PBZs are much more widespread in some other eukaryotes[48,60-62].Up till now,PBZ domains are only found in three human proteins,including histone chaperone APLF,checkpoint with FHA and RING finger(CHFR),and sensitive to nitrogen mustard 1A(SNM1A)[48,60-62].Crystal structures of APLF and CHFR show that PBZs are essential for their functions.Initial analysis of CHFR primary sequence has identified a zing finger called C2H2,which binds to PAR efficiently.Therefore,this motif is defined as a new PAR binding module termed PBZ[60].APLF containstwotandemPBZdomainstermedF1andF2.Although F1andF2canrecognizethePARchainindependently,presence of both domains remarkably increase the affinity of PAR chain binding,whichisover1000timesmoreefficientthantheisolated PBZ domain[63].Structural analysis demonstrates that PBZ module contains a central zinc ion coordinated by two cysteine and two histidine residues,which can recognize adenines in two neighboring ADP-ribose units of the PAR chain.This type of recognition renders the PBZ motifs to be the truly specific PAR binding modules[63](Figure 2).
The WWE domain
The WWE domain is the most recently discovered PAR-binding domain,named after the three strictly conserved amino acid residues,tryptophan-tryptophan-glutamate(WWE)[64]. The WWE domains,which can recognize iso-ADPR of PAR chain with high affinity,tightly links ubiquitination and PARylation signal pathways.The iso-ADPR which contains a characteristic bond,2′,1′′-O-glycosidic ribose-ribose is the signature of PAR chains[64].The negatively-charged phosphate groups of the iso-ADPR can bind the positively-charged WWE domain[64].The WWE domain is primarily found in two distinct protein families,including the E3 ubiquitin ligases(RNF146,deltex1,and TRIP12)and the PARPs(PARP8 and PARP11-14)[63].So far,the function of WWE domain has been well described for RNF146/Iduna.RNF146 recognizes PAR chain and ubiquitinates DNA repair proteins such as XRCC1,PARP1,DNA ligase III,and Ku70.The PARylated proteins are targeted to proteosome for degradation[64,65]. Taken together,the WWE domain-containing proteins are tightly linked with and influence each other(Figure 2).
The Macro domain
The Macro domain,which consists of 130-190 amino acid residues,is evolutionarily conserved and widely spread throughout all kingdoms of organisms.This is distinct from the PBZ and WWE domains.It is estimated that more than 300 proteins,including 11 human proteins,with a diverse set of biological functions possess the Macro domain[66].Macro domains can bind to the terminal ADPR of PAR,MAR,as well as O-acetyl-ADPR[66-68].Some proteins such as amplified in liver cancer 1(ALC1,also known as CHD1L),can interact with PAR chains through Macro domains and catalyze PARP1-stimulated nucleosome sliding,thus participating in DDR and chromatin remodeling[69,70].Some other Macro domain-containing proteins,in addition to their binding ability,also exhibit catalytic activity on the hydrolysis of PAR chains,making the Macro domains unique among the other PAR-binding modules.These include PARG[22],TARG1[26,71],and MacroD1/2[28,31](Figure 2).PARG enzyme uses Macro domain for the binding and hydrolysis of PAR chains,as we outlined above.
Additional domains
ItiswellknownthatFHAandBRCTdomainscanbindtophosphorylated proteins and modify protein-protein interactions[72].Recently,it was reported that the phosphate-binding pocket in the central BRCT domain of BARD1 is required forselective binding to PAR chain[50,73].Meanwhile,BRCT domainpromotestheinteractionbetweenBARD1andPARP1. Moreover,the FHA domains of aprataxin(APTX)and PNKP confer affinity to iso-ADPR of PAR chain[50,73].

Figure 2 PAR-binding modules
The OB-fold is an ssDNA or ssRNA binding domain that has been found in proteins from all three kingdoms.OB-fold comprises 70-150 AA residues forming five-stranded beta-barrel with a terminating alpha-helix[57].Interestingly,it is reportedrecentlythattheOB-foldcanbindtothePAR-specific iso-ADPR and such binding is required to bring the ssDNA-binding protein 1(SSB1)to sites of DNA damage[58].
The PIN domain-containing proteins serve as nucleases that cleave ssDNA/ssRNA in a sequence-specific manner[74].The PIN domain consists of~130 amino acid residues characterized by a group of three strictly conserved acidic amino acid residues[75].Our recent study found that the PIN domain of EXO1 recognizes PAR in DDR[58].
The RRM is one of the most abundant protein domains in eukaryotes,which can serve as a plastic RNA-binding platform to regulate post-transcriptional gene expression[76].Several RRM-containing proteins have been reported to assemble at sites of PAR formation to promote DDR[77,78].
It is reported that some RNA and DNA binding motifs can recognize PAR chains.Motifs enriched in arginines and glycines,which are termed glycine-arginine-rich(GAR)domains and/or RGG boxes,were identified several decades ago. RGG boxes are found in more than 1000 human proteins that are involved in numerous biological processes including transcription and DDR[79].RGG boxes in the RNA-binding proteins such as FUS/TLS,EWS/EWSR1,TAF15,SAFB1,SAFA,and hnRNPUL1/2,have been identified,and these proteins can be recruited to DNA damage sites via binding to PAR chain through RGG boxes[80-86].
PARylation and ubiquitination
Ubiquitin is a small regulatory protein consisting of 76 amino acid residues,which has been found in almost all tissues of eukaryotic organisms.It can be covalently transferred to a Lys residue of an acceptor protein.This process is termed ubiquitination[87].The ubiquitination pathway in cells is an elaborate system for targeting unwanted proteins for degradation,carried out by three classes of enzymes,E1,E2,andE3.Ubiquitinisfirstactivatedbyubiquitinactivating enzyme(E1)before being transferred to the active site of E1 in an ATP-dependent manner.Then the ubiquitin molecule is passed on to the second enzyme,ubiquitinconjugating enzyme(E2),whereubiquitin is linkedby another thioester bond to the Cys active site of E2.Finally,with the help of a third enzyme,ubiquitin protein ligase(E3),ubiquitin is transferred from E2 to a Lys residue on a substrate protein.Additional ubiquitin molecules can be linked to the first one to form a poly-ubiquitin chain usually targeting the protein to the proteasome[87].
Recent studies have demonstrated that PARylation can serve as a signal for the ubiquitination and promote the degradation of PARsylated proteins[88-90].Some E3 ligases bind PAR via either a WWE(RNF146,also known as Iduna)domain or a PBZ(CHFR)domain[43,44,46].The relationship betweenPARylationandubiquitinationhasbeenwell described in the RING-type E3 ubiquitin ligase,RNF146. The RNF146 WWE domain recognizes the PAR chain via interacting with iso-ADPR(Figure 2),functioning as an allosteric signal that changes the RING domain conformation from a catalytically-inactive state to an active one.RNF146 can polyubiquitylate many repair factors in a PAR-dependent manner,such as PARP-2,XRCC1,DNA ligase III,and Ku70[62].The discovery of a direct connection between PARylation and ubiquitination provides us with a new interpretation of the signaling function of PAR—degradation of proteins in a timely and orchestrated manner.
Dysregulation of PARylation and human diseases
PARP1 is a key facilitator of DDR and is implicated in tumorigenesis of several malignancies,particularly those associated with dysfunctional DNA repair pathways[37].Recent studies further demonstrate that transcript,protein,and enzyme activity of PARP1were increased in several tumor types with the most striking differences noticed in ovarian cancer,hepatocellular cancer,colorectal cancer,and leukemia[76-78].Given that PARP1 has an important role in DDR,a novel therapeutic targeting PARP1 has been developed to treat cancers through increasing tumor sensitivity to chemotherapeutic agents and also through inducing‘‘synthetic lethality”in cells[78].Now PARP inhibitors have demonstrated efficacy in a number of tumor types,including platinum-sensitive epithelial ovarian cancer[50],breast cancer with mutation in BRCA1 or BRCA2[91],and prostate cancer[92].Olaparib is a PARP inhibitor that blocks enzymes involved in repairing damaged DNA[92].Recently olaparib has been licensed as monotherapy for the treatment of patients with hereditary BRCA1 or BRCA2 mutations[91].
Perspectives and conclusions
Over the last decades,PARylation has been proved to be involved in numerous cellular functions including DDR. PAR serves as an initial sensor and mediates the early recruitment of DNA damage repair machineries.As a kind of protein modification,PARylation is tightly and dynamically regulated. PAR chain synthesis is mediated by several PARPs,whereas PARG mainly takes charge of PAR chain degradation.Great strides have been made in the past few decades to decipher the PARylation regulatory processes and the underlying molecular mechanisms.However,many questions remain to be answered.First,other NAD+-consuming enzymes,such as sirtuin 1,are thought to compete for NAD+with PARPs[9]. What is the reciprocal influence of these enzymes?Moreover,how these DNA damage factors are assembled at the DNA damage sites via PAR chains is still unclear exactly,as PAR chain does not have any sequence specificity.In addition,new molecular or chemical methods need to be developed to better achieve cell-permeable PARG or/and ARH inhibitors.More investigations are needed to address these questions in the future.In this regard,a better understanding of the biochemical and functional properties of PARylation in DNA repair may provide new clues to answer these fundamental questions.
Competing interests
The authors declare that they have no competing financial interests.
Acknowledgments
References
[1]Ciccia A,Elledge SJ.The DNA damage response:making it safe to play with knives.Mol Cell 2010;40:179-204.
[2]Lindahl T,Barnes DE.Repair of endogenous DNA damage.Cold Spring Harb Symp Quant Biol 2000;65:127-33.
[3]Hoeijmakers JH.DNA damage,aging,and cancer.N Engl J Med 2009;361:1475-85.
[4]Wang Y,Kim NS,Haince JF,Kang HC,David KK,Andrabi SA,et al.Poly(ADP-ribose)(PAR)binding to apoptosis-inducing factor is critical for PAR polymerase-1-dependent cell death(parthanatos).Sci Signal 2011;4:ra20.
[5]Gibson BA,Kraus WL.New insights into the molecular and cellular functions of poly(ADP-ribose)and PARPs.Nat Rev Mol Cell Biol 2012;13:411-24.
[6]Perina D,Mikoc A,Ahel J,Cetkovic H,Zaja R,Ahel I. Distribution of protein poly(ADP-ribosyl)ation systems across all domains of life.DNA Repair(Amst)2014;23:4-16.
[7]Luo X,Kraus WL.On PAR with PARP:cellular stress signaling throughpoly(ADP-ribose)andPARP-1.GenesDev 2012;26:417-32.
十八届四中全会通过的《中共中央关于全面推进依法治国若干重大问题的决定》,首次提出实行国家机关“谁执法谁普法”普法责任制。2017年5月,中共中央办公厅、国务院办公厅印发《关于实行国家机关“谁执法谁普法”普法责任制的意见》(下文简称《意见》),《意见》是对党的十八届四中全会决定明确提出的实行国家机关“谁执法谁普法”普法责任制这一重要要求的具体化、规范化。
[8]Hakme A,Wong HK,Dantzer F,Schreiber V.The expanding field of poly(ADP-ribosyl)ation reactions.’Protein Modifications: BeyondtheUsualSuspects’ReviewSeries.EMBORep 2008;9:1094-100.
[9]Hottiger MO.Nuclear ADP-Ribosylation and Its Role in Chromatin Plasticity,Cell Differentiation,and Epigenetics.Annu Rev Biochem 2015;84:227-63.
[10]Rouleau M,Patel A,Hendzel MJ,Kaufmann SH,Poirier GG. PARPinhibition:PARP1andbeyond.NatRevCancer 2010;10:293-301.
[11]D’Amours D,Desnoyers S,D’Silva I,Poirier GG.Poly(ADP-ribosyl)ation reactions in the regulation of nuclear functions. Biochem J 1999;342:249-68.
[12]Tallis M,Morra R,Barkauskaite E,Ahel I.Poly(ADP-ribosyl)ation in regulation of chromatin structure and the DNA damage response.Chromosoma 2014;123:79-90.
[13]Altmeyer M,Hottiger MO.Poly(ADP-ribose)polymerase 1 at the crossroad of metabolic stress and inflammation in aging.Aging(Albany NY)2009;1:458-69.
[14]Vyas S,Chang P.New PARP targets for cancer therapy.Nat Rev Cancer 2014;14:502-9.
[15]De Vos M,Schreiber V,Dantzer F.The diverse roles and clinical relevance of PARPs in DNA damage repair:current state of the art.Biochem Pharmacol 2012;84:137-46.
[16]Dregalla RC,Zhou J,Idate RR,Battaglia CL,Liber HL,Bailey SM.Regulatory roles of tankyrase 1 at telomeres and in DNA repair:suppression of T-SCE and stabilization of DNA-PKcs. Aging(Albany NY)2010;2:691-708.
[17]Juarez-Salinas H,Levi V,Jacobson EL,Jacobson MK.Poly(ADP-ribose)has a branched structure in vivo.J Biol Chem 1982;257:607-9.
[18]Tanuma S,Kanai Y.Poly(ADP-ribosyl)ation of chromosomal proteinsintheHeLaS3cellcycle.JBiolChem 1982;257:6565-70.
[19]Miwa M,Saikawa N,Yamaizumi Z,Nishimura S,Sugimura T. Structure of poly(adenosine diphosphate ribose):identification of 2’-[1”-ribosyl-2”-(or3”-)(1”’-ribosyl)]adenosine-5’,5”,5”’-tris(phosphate)as a branch linkage.Proc Natl Acad Sci U S A 1979;76:595-9.
[20]Dunstan MS,Barkauskaite E,Lafite P,Knezevic CE,Brassington A,Ahel M,et al.Structure and mechanism of a canonical poly(ADP-ribose)glycohydrolase.Nat Commun 2012;3:878.
[21]Kim IK,Kiefer JR,Ho CM,Stegeman RA,Classen S,Tainer JA,et al.Structure of mammalian poly(ADP-ribose)glycohydrolase reveals a flexible tyrosine clasp as a substrate-binding element. Nat Struct Mol Biol 2012;19:653-6.
[22]Slade D,Dunstan MS,Barkauskaite E,Weston R,Lafite P,Dixon N,et al.The structure and catalytic mechanism of a poly(ADP-ribose)glycohydrolase.Nature 2011;477:616-20.
[23]Ueda K,Oka J,Naruniya S,Miyakawa N,Hayaishi O.Poly ADP-ribose glycohydrolase from rat liver nuclei,a novel enzyme degradingthepolymer.BiochemBiophysResCommun 1972;46:516-23.
[24]Mueller-Dieckmann C,Kernstock S,Lisurek M,von Kries JP,Haag F,Weiss MS,et al.The structure of human ADP-ribosylhydrolase 3(ARH3)provides insights into the reversibility of protein ADP-ribosylation.Proc Natl Acad Sci U S A 2006;103:15026-31.
[25]Niere M,Mashimo M,Agledal L,Dolle C,Kasamatsu A,Kato J,et al.ADP-ribosylhydrolase 3(ARH3),not poly(ADP-ribose)glycohydrolase(PARG)isoforms,is responsible for degradation of mitochondrial matrix-associated poly(ADP-ribose).J Biol Chem 2012;287:16088-102.
[26]Sharifi R,Morra R,Appel CD,Tallis M,Chioza B,Jankevicius G,et al.Deficiency of terminal ADP-ribose protein glycohydrolase TARG1/C6orf130 in neurodegenerative disease.EMBO J 2013;32:1225-37.
[27]Barkauskaite E,Brassington A,Tan ES,Warwicker J,Dunstan MS,Banos B,et al.Visualization of poly(ADP-ribose)bound to PARG reveals inherent balance between exo-and endo-glycohydrolase activities.Nat Commun 2013;4:2164.
[28]Jankevicius G,Hassler M,Golia B,Rybin V,Zacharias M,Timinszky G,et al.A family of macrodomain proteins reverses cellularmono-ADP-ribosylation.NatStructMolBiol 2013;20:508-14.
[29]Rosenthal F,Feijs KL,Frugier E,Bonalli M,Forst AH,Imhof R,et al.Macrodomain-containing proteins are new mono-ADP-ribosylhydrolases.Nat Struct Mol Biol 2013;20:502-7.
[30]Neuvonen M,Ahola T.Differential activities of cellular and viral macro domain proteins in binding of ADP-ribose metabolites.J Mol Biol 2009;385:212-25.
[31]Chen D,Vollmar M,Rossi MN,Phillips C,Kraehenbuehl R,Slade D,et al.Identification of macrodomain proteins as novel O-acetyl-ADP-ribose deacetylases.J Biol Chem 2011;286:13261-71.
[32]Wielckens K,Schmidt A,George E,Bredehorst R,Hilz H.DNA fragmentation and NAD depletion.Their relation to the turnover of endogenous mono(ADP-ribosyl)and poly(ADP-ribosyl)proteins.J Biol Chem 1982;257:12872-7.
[33]Fatokun AA,Dawson VL,Dawson TM.Parthanatos:mitochondrial-linked mechanisms and therapeutic opportunities.Br J Pharmacol 2014;171:2000-16.
[34]Jiricny J.The multifaceted mismatch-repair system.Nat Rev Mol Cell Biol 2006;7:335-46.
[35]Caldecott KW.Single-strand break repair and genetic disease. Nat Rev Genet 2008;9:619-31.
[36]Li M,Yu X.Function of BRCA1 in the DNA damage response is mediated by ADP-ribosylation.Cancer Cell 2013;23:693-704.
[37]Dantzer F,de La Rubia G,Menissier-De Murcia J,Hostomsky Z,de Murcia G,Schreiber V.Base excision repair is impaired in mammalian cells lacking Poly(ADP-ribose)polymerase-1.Biochemistry 2000;39:7559-69.
[38]Noren Hooten N,Kompaniez K,Barnes J,Lohani A,Evans MK. Poly(ADP-ribose)polymerase 1(PARP-1)binds to 8-oxoguanine-DNA glycosylase(OGG1).J Biol Chem 2011;286:44679-90.
[39]Frouin I,Maga G,Denegri M,Riva F,Savio M,Spadari S,et al. Human proliferating cell nuclear antigen,poly(ADP-ribose)polymerase-1,and p21waf1/cip1.A dynamic exchange of partners.J Biol Chem 2003;278:39265-8.
[40]Harris JL,Jakob B,Taucher-Scholz G,Dianov GL,Becherel OJ,Lavin MF.Aprataxin,poly-ADP ribose polymerase 1(PARP-1)and apurinic endonuclease 1(APE1)function together to protect thegenomeagainstoxidativedamage.HumMolGenet 2009;18:4102-17.
[41]Schreiber V,Ame JC,Dolle P,Schultz I,Rinaldi B,Fraulob V,et al.Poly(ADP-ribose)polymerase-2(PARP-2)is required for efficient base excision DNA repair in association with PARP-1 and XRCC1.J Biol Chem 2002;277:23028-36.
[42]Masson M,Niedergang C,Schreiber V,Muller S,Menissier-de Murcia J,de Murcia G.XRCC1 is specifically associated with poly(ADP-ribose)polymerase and negatively regulates its activity following DNA damage.Mol Cell Biol 1998;18:3563-71.
[43]Ceccaldi R,Rondinelli B,D’Andrea AD.Repair Pathway Choices and Consequences at the Double-Strand Break.Trends Cell Biol 2016;26:52-64.
[44]Chiruvella KK,Liang Z,Wilson TE.Repair of double-strand breaks by end joining.Cold Spring Harb Perspect Biol 2013;5: a012757.
[45]Karanam K,Kafri R,Loewer A,Lahav G.Quantitative live cell imaging reveals a gradual shift between DNA repair mechanisms and a maximal use of HR in mid S phase.Mol Cell 2012;47:320-9.
[46]Lieber MR.The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway.Annu Rev Biochem 2010;79:181-211.
[47]Morrison C,Smith GC,Stingl L,Jackson SP,Wagner EF,Wang ZQ.Genetic interaction between PARP and DNA-PK in V(D)J recombination and tumorigenesis.Nat Genet 1997;17:479-82.
[48]Ruscetti T,Lehnert BE,Halbrook J,Le Trong H,Hoekstra MF,Chen DJ,et al.Stimulation of the DNA-dependent protein kinase by poly(ADP-ribose)polymerase.J Biol Chem 1998;273:14461-7.[49]Spagnolo L,Barbeau J,Curtin NJ,Morris EP,Pearl LH. Visualization of a DNA-PK/PARP1 complex.Nucleic Acids Res 2012;40:4168-77.
[50]Li M,Lu LY,Yang CY,Wang S,Yu X.The FHA and BRCT domainsrecognizeADP-ribosylationduringDNAdamage response.Genes Dev 2013;27:1752-68.
[51]Deriano L,Roth DB.Modernizing the nonhomologous endjoining repertoire:alternative and classical NHEJ share the stage. Annu Rev Genet 2013;47:433-55.
[52]Iliakis G.Backup pathways of NHEJ in cells of higher eukaryotes: cell cycle dependence.Radiother Oncol 2009;92:310-5.
[53]Schultz N,Lopez E,Saleh-Gohari N,Helleday T.Poly(ADP-ribose)polymerase(PARP-1)has a controlling role in homologous recombination.Nucleic Acids Res 2003;31:4959-64.
[54]West SC.Molecular views of recombination proteins and their control.Nat Rev Mol Cell Biol 2003;4:435-45.
[55]Heyer WD,Ehmsen KT,Liu J.Regulation of homologous recombination in eukaryotes.Annu Rev Genet 2010;44:113-39.
[56]Haince JF,McDonald D,Rodrigue A,Dery U,Masson JY,Hendzel MJ,et al.PARP1-dependent kinetics of recruitment of MRE11 and NBS1 proteins to multiple DNA damage sites.J Biol Chem 2008;283:1197-208.
[57]Zhang F,Shi J,Bian C,Yu X.Poly(ADP-Ribose)Mediates the BRCA2-Dependent Early DNA Damage Response.Cell Rep 2015;13:678-89.
[58]Zhang F,Shi J,Chen SH,Bian C,Yu X.The PIN domain of EXO1 recognizes poly(ADP-ribose)in DNA damage response. Nucleic Acids Res 2015;43:10782-94.
[59]Zaja R,Mikoc A,Barkauskaite E,Ahel I.Molecular Insights into Poly(ADP-ribose)Recognition and Processing.Biomolecules 2012;3:1-17.
[60]Ahel I,Ahel D,Matsusaka T,Clark AJ,Pines J,Boulton SJ,et al. Poly(ADP-ribose)-binding zinc finger motifs in DNA repair/ checkpoint proteins.Nature 2008;451:81-5.
[61]Mehrotra PV,Ahel D,Ryan DP,Weston R,Wiechens N,Kraehenbuehl R,et al.DNA repair factor APLF is a histone chaperone.Mol Cell 2011;41:46-55.
[62]Oberoi J,Richards MW,Crumpler S,Brown N,Blagg J,Bayliss R.Structural basis of poly(ADP-ribose)recognition by the multizinc binding domain of checkpoint with forkhead-associated and RING Domains(CHFR).J Biol Chem 2010;285:39348-58.
[63]Li GY,McCulloch RD,Fenton AL,Cheung M,Meng L,Ikura M,et al.Structure and identification of ADP-ribose recognition motifs of APLF and role in the DNA damage response.Proc Natl Acad Sci U S A 2010;107:9129-34.
[64]Wang Z,Michaud GA,Cheng Z,Zhang Y,Hinds TR,Fan E,et al.Recognition of the iso-ADP-ribose moiety in poly(ADP-ribose)by WWE domains suggests a general mechanism for poly(ADP-ribosyl)ation-dependentubiquitination.GenesDev 2012;26:235-40.
[65]Kang HC,Lee YI,Shin JH,Andrabi SA,Chi Z,Gagne JP,et al. Iduna is a poly(ADP-ribose)(PAR)-dependent E3 ubiquitin ligase that regulates DNA damage.Proc Natl Acad Sci U S A 2011;108:14103-8.
[66]Feijs KL,Forst AH,Verheugd P,Luscher B.Macrodomaincontaining proteins:regulating new intracellular functions of mono(ADP-ribosyl)ation.Nat Rev Mol Cell Biol 2013;14:443-51.
[67]Timinszky G,Till S,Hassa PO,Hothorn M,Kustatscher G,Nijmeijer B,et al.A macrodomain-containing histone rearranges chromatin upon sensing PARP1 activation.Nat Struct Mol Biol 2009;16:923-9.
[68]Kustatscher G,Hothorn M,Pugieux C,Scheffzek K,Ladurner AG.Splicing regulates NAD metabolite binding to histone macroH2A.Nat Struct Mol Biol 2005;12:624-5.
[69]Gottschalk AJ,Timinszky G,Kong SE,Jin J,Cai Y,Swanson SK,et al.Poly(ADP-ribosyl)ation directs recruitment and activation of an ATP-dependent chromatin remodeler.Proc Natl Acad Sci U S A 2009;106:13770-4.
[70]Ahel D,Horejsi Z,Wiechens N,Polo SE,Garcia-Wilson E,Ahel I,et al.Poly(ADP-ribose)-dependent regulation of DNA repair by thechromatinremodelingenzymeALC1.Science 2009;325:1240-3.
[71]Peterson FC,Chen D,Lytle BL,Rossi MN,Ahel I,Denu JM,et al.Orphan macrodomain protein(human C6orf130)is an O-acyl-ADP-ribose deacylase:solution structure and catalytic properties.J Biol Chem 2011;286:35955-65.
[72]Reinhardt HC,Yaffe MB.Phospho-Ser/Thr-binding domains: navigating the cell cycle and DNA damage response.Nat Rev Mol Cell Biol 2013;14:563-80.
[73]DaRosa PA,Wang Z,Jiang X,Pruneda JN,Cong F,Klevit RE,et al.Allosteric activation of the RNF146 ubiquitin ligase by a poly(ADP-ribosyl)ation signal.Nature 2015;517:223-6.
[74]Arcus VL,McKenzie JL,Robson J,Cook GM.The PIN-domain ribonucleases and the prokaryotic VapBC toxin-antitoxin array. Protein Eng Des Sel 2011;24:33-40.
[75]Arcus VL,Rainey PB,Turner SJ.The PIN-domain toxinantitoxin array in mycobacteria.Trends Microbiol 2005;13:360-5.
[76]Maris C,Dominguez C,Allain FH.The RNA recognition motif,a plastic RNA-binding platform to regulate post-transcriptional gene expression.FEBS J 2005;272:2118-31.
[77]Gagne JP,Hunter JM,Labrecque B,Chabot B,Poirier GG.A proteomic approach to the identification of heterogeneous nuclear ribonucleoproteins as a new family of poly(ADP-ribose)-binding proteins.Biochem J 2003;371:331-40.
[78]Ji Y,Tulin AV.Poly(ADP-ribosyl)ation of heterogeneous nuclear ribonucleoproteinsmodulatessplicing.NucleicAcidsRes 2009;37:3501-13.
[79]Thandapani P,O’Connor TR,Bailey TL,Richard S.Defining the RGG/RG motif.Mol Cell 2013;50:613-23.
[80]Izhar L,Adamson B,Ciccia A,Lewis J,Pontano-Vaites L,Leng Y,et al.A Systematic Analysis of Factors Localized to Damaged Chromatin Reveals PARP-Dependent Recruitment of Transcription Factors.Cell Rep 2015;11:1486-500.
[81]Altmeyer M,Toledo L,Gudjonsson T,Grofte M,Rask MB,Lukas C,et al.The chromatin scaffold protein SAFB1 renders chromatin permissive for DNA damage signaling.Mol Cell 2013;52:206-20.
[82]Mastrocola AS,Kim SH,Trinh AT,Rodenkirch LA,Tibbetts RS.The RNA-binding protein fused in sarcoma(FUS)functions downstream of poly(ADP-ribose)polymerase(PARP)in response to DNA damage.J Biol Chem 2013;288:24731-41.
[83]Rulten SL,Rotheray A,Green RL,Grundy GJ,Moore DA,Gomez-Herreros F,et al.PARP-1 dependent recruitment of the amyotrophic lateral sclerosis-associated protein FUS/TLS to sitesofoxidativeDNAdamage.NucleicAcidsRes 2014;42:307-14.
[84]Polo SE,Blackford AN,Chapman JR,Baskcomb L,Gravel S,Rusch A,et al.Regulation of DNA-end resection by hnRNPU-like proteins promotes DNA double-strand break signaling and repair.Mol Cell 2012;45:505-16.
[85]Hong Z,Jiang J,Ma J,Dai S,Xu T,Li H,et al.The role of hnRPUL1 involved in DNA damage response is related to PARP1.PLoS One 2013;8:e60208.
[86]Britton S,Dernoncourt E,Delteil C,Froment C,Schiltz O,Salles B,et al.DNA damage triggers SAF-A and RNA biogenesis factors exclusion from chromatin coupled to R-loops removal. Nucleic Acids Res 2014;42:9047-62.
[87]Weissman AM,Shabek N,Ciechanover A.The predator becomes the prey:regulating the ubiquitin system by ubiquitylation and degradation.Nat Rev Mol Cell Biol 2011;12:605-20.
[88]Huang SM,Mishina YM,Liu S,Cheung A,Stegmeier F,Michaud GA,et al.Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling.Nature 2009;461:614-20.
[89]Levaot N,Voytyuk O,Dimitriou I,Sircoulomb F,Chandrakumar A,Deckert M,et al.Loss of Tankyrase-mediated destruction of 3BP2 is the underlying pathogenic mechanism of cherubism.Cell 2011;147:1324-39.
[90]Guettler S,LaRose J,Petsalaki E,Gish G,Scotter A,Pawson T,et al.Structural basis and sequence rules for substrate recognition by Tankyrase explain the basis for cherubism disease.Cell 2011;147:1340-54.
[91]Crafton SM,Bixel K,Hays JL.PARP inhibition and gynecologic malignancies:a review of current literature and on-going trials. Gynecol Oncol 2016.http://dx.doi.org/10.1016/j.ygyno.2016.05.003.
[92]Raison N,Elhage O,Dasgupta P.Getting personal with prostate cancer:DNA-repair defects and olaparib in metastatic prostate cancer.BJU Int 2016.http://dx.doi.org/10.1111/bju.13522.
23 February 2016;revised 29 April 2016;accepted 2 May 2016
*Corresponding author.
E-mail:xyu@coh.org(Yu X).aORCID:0000-0003-2711-3868.bORCID:0000-0002-0751-7390.
Peer review under responsibility of Beijing Institute of Genomics,Chinese Academy of Sciences and Genetics Society of China.
http://dx.doi.org/10.1016/j.gpb.2016.05.001
1672-0229ⓒ2016 The Authors.Production and hosting by Elsevier B.V.on behalf of Beijing Institute of Genomics,Chinese Academy of Sciences and Genetics Society of China.
This is an open access article under the CC BY license(http://creativecommons.org/licenses/by/4.0/).
猜你喜欢
时尚设计与工程(2022年3期)2022-08-10 08:41:06
今日农业(2020年19期)2020-12-14 14:16:52
今日农业(2019年12期)2019-08-13 00:49:58
青海政报(2017年4期)2017-07-24 14:04:48
学生天地(2017年35期)2017-04-19 01:50:08
青海政报(2017年22期)2017-04-09 06:45:55
青海政报(2017年21期)2017-03-16 06:05:09
天津造纸(2016年2期)2017-01-15 14:03:36
天津造纸(2016年2期)2017-01-15 14:03:32
中学生数理化·教与学(2016年2期)2016-11-25 02:21:13
 Genomics,Proteomics & Bioinformatics2016年3期
Genomics,Proteomics & Bioinformatics2016年3期
- Genomics,Proteomics & Bioinformatics的其它文章
- DNA End Resection:Facts and Mechanisms
- UFMylation:A Unique&Fashionable Modification for Life
- New Edges of RNA Adenosine Methylation Modifications
- Topoisomerase I in Human Disease Pathogenesis and Treatments
- Connecting Malfunctioning Glial Cells and Brain Degenerative Disorders
- DNA Damage Response in Hematopoietic Stem Cell Ageing