
接种摩西管柄囊霉(Funneliformis mosseae)对连作大豆根腐病病原菌菌群结构的影响
2016-09-10 02:00:16崔佳琦范雪露蔡柏岩黑龙江大学生命科学学院哈尔滨50080黑龙江省普通高等学校微生物重点实验室哈尔滨50080
农学学报 2016年7期
崔佳琦,范雪露,蔡柏岩(黑龙江大学生命科学学院,哈尔滨50080;黑龙江省普通高等学校微生物重点实验室,哈尔滨50080)
接种摩西管柄囊霉(Funneliformis mosseae)对连作大豆根腐病病原菌菌群结构的影响
崔佳琦1,2,范雪露1,2,蔡柏岩1,2
(1黑龙江大学生命科学学院,哈尔滨150080;
2黑龙江省普通高等学校微生物重点实验室,哈尔滨150080)
笔者将生物修复方法应用于连作大豆根腐病防治中,旨在降低连作大豆根腐病发生以及提高大豆质量。试验选择3个大豆品种作为试验材料并接种摩西管柄囊霉(Funneliformis mosseae),通过PCRDGGE技术研究F.mosseae对连作大豆根系以及根际土壤根腐病病原菌菌群结构的影响。试验结果表明:3个大豆品种接种F.mosseae后,试验组根样以及根际土样根腐病病原菌丰度以及多样性指数显著低于对照组,并且接种F.mosseae对不同大豆品种防治效果存在差异;聚类分析结果也验证了接种F. mosseae对连作大豆根样以及根际土样根腐病病原菌产生以及分布产生影响。
根腐病;F.mosseae;大豆连作;菌群结构
0 引言
大豆(Glycine max[L.]Merr.)是重要的油料以及经济作物,为人类提供了丰富的脂质以及蛋白质资源[1-2]。黑龙江地区是中国大豆主要生产基地,大豆种植面积以及产量在中国大豆产业中占有重要地位。农业生产中,作物种植方式是影响土壤微生物菌群结构的关键因素[3]。近年来,由于黑龙江地区大豆种植方式的改变,即大豆长期连作,造成了土壤中微生物菌群结构以及土壤理化性质发生了显著的变化[4],大豆田土壤由高肥力的细菌型逐渐转变为低肥力的真菌型[5],最终导致大豆根腐病(Root rot)发生频率显著提高。因此,探索如何缓解以及防治大豆根腐病发生已成为首要课题。
近年来,高效、低耗以及环保型生物防治方法突显出极大的研究价值以及应用潜力。科研人员已将植物-微生物共生体应用于植物抗逆性研究中,并取得了丰硕的研究成果。丛枝菌根(Arbuscular Mycorrhizal)真菌是土壤生态系统中同时具有植物根系以及微生物特性的互惠共生体[6]。AM真菌可以与陆生维管植物根系建立共生关系,形成“菌根”结构。AM真菌通过形成庞大的菌丝体结构,可以扩大植物根系对根际土壤中水分以及矿质元素的吸收面积[7-8],进而加快叶绿体的形成,提高植株光合速率以及生物量[9-10]。AM真菌在调节植物抗逆性过程中,通过对植物相关代谢、防御性基因表达的影响,达到对植物防御性蛋白、防御性酶类以及次生代谢产物的调控[11-12],进而增强植株抗病虫害[13]、抗干旱[14]以及抗重金属污染能力[15-16]。因此,AM真菌在维持生态系统多样性以及微生态系统稳定性方面具有重要作用。
鉴于AM真菌在土壤生态系统中的重要作用以及生物修复方法在防治作物病害方面的研究基础。本试验通过向黑龙江地区种植面积较大、代表性较强的3个非转基因大豆品种不同连作年限盆栽土壤中接种筛选自黑龙江地区连作大豆田优势AM真菌-摩西管柄囊霉(F.mosseae),研究F.mosseae对不同大豆品种根系以及根际土壤中根腐病病原菌菌群结构的影响。旨在为AM菌剂应用于防治大豆根腐病发生做出前期探索以及实践,并为AM菌剂应用于缓解大豆连作障碍提供理论依据。
1 材料与方法
1.1试验样地
本试验于2014年5—8月在哈尔滨工业大学糖业研究所试验站进行(46°40′N,130°10′E)。分别选用未种植过大豆土壤(L0)、连作1年大豆田土壤(已种植过2年大豆田土壤,L1)以及连作2年大豆田土壤(已种植过3年大豆田土壤,L2)进行试验。
试验土壤初始理化指标:有机质26.13 g/kg,全N 1.69g/kg、全K25.4g/kg、全P5.5g/kg、碱解N133.1mg/kg、速效P 13.14 mg/kg,速效K 206 mg/kg、pH 7.0。
1.2试验材料以及供试菌种
试验选用黑龙江地区种植面积较广、代表性较强的3个非转基因大豆品种:‘垦丰16'(KF16,中间型,脂肪含量:20.00%,蛋白质含量:40.17%),‘黑农44' (HN44,高脂肪型,脂肪含量:23.01%,蛋白质含量:36.06%),‘黑农48'(HN48,高蛋白型,脂肪含量:19.50%,蛋白质含量:45.23%)。
试验菌种筛选自黑龙江地区连作大豆田优势AM真菌-摩西管柄囊霉(F.mosseae),保藏于中国科学院微生物研究所,菌种保藏号:CGMCC No.3012。
1.3试验方法
1.3.1试验设计 试验采用盆栽方式。每盆装风干土壤4 kg,土壤分别取自L0、L1以及L2。土壤装好后,向盆体中接种45 g F.mosseae菌剂(试验组,简写:TF),将菌剂与土壤充分混匀,并设置对照组(简写:CF,未接种F.mosseae)。每个试验处理重复6次。2014年5月17日进行播种,每盆留苗3株,生产管理同大豆田。同时在盆体周围摆放种植大豆、同样大小的盆体,作为保护行,防止边际效应。
1.3.2样品采集及处理 在大豆苗期(简写:S,播种后30天)和分枝期(简写:B,播种后60天)进行样品采集。采集土样时,将盆体土壤表面杂物去除。利用铲子挖10~20 cm土壤剖面,将大豆根系取出。用手轻轻抖动大豆植株后,附着在大豆根系表面1~3 cm处土壤,即根际土样。将盆体中3株大豆植株根际土样混合在一起,记为1个样品。采集根样时,利用自来水将大豆根系表面土壤洗净,利用剪刀将盆体中3株大豆植株根系剪下并混合,记为1个样品。随机选择同一处理、不同盆体,重复上述步骤3次,将不同连作年限根样分别标记为RL0、RL1以及RL2,不同连作年限土样分别标记为SL0、SL1以及SL2。样品收集好后,立即带回实验室并保存在4℃冰箱中。
1.3.3PCR-DGGE大豆根样以及根际土样总DNA分别利用十六烷基三甲基溴化铵(CTAB)法和土壤基因组提取试剂盒(E.Z.N.A.Soil DNAKit,Omega,USA)进行提取。大豆根样以及根际土样总基因组提取完毕后,通过真菌通用性引物:NS1(上游引物):5′-GTAGTCATATGCTTGTCTC-3′和FungGC(下游引物):5′-CGCCCGCCGCGCCCCGCGCCCGGCCCGC CGCCCCCGCCCCATTCCCCGTTACCCGTTG-3′对真菌18S rRNA(V1+V2)区进行特异性扩增。PCR反应体系为(50 μL):10×Ex Taq Buffer(含Mg2+)5.0 μL,2.5 mmol/L dNTP Mixture 4.0 μL,5 U Taq酶0.2 μL,DNA模板1.0 μL,1.0 μmol/L引物各5 μL,ddH2O 29.8 μL。反应程序:94℃变性3 min;94℃30 s,57℃1 min,72℃1 min,共35个循环;72℃延伸10 min;4℃保存。PCR扩增产物进行1%琼脂糖凝胶电泳检测,并将扩增产物置于-20℃冰箱保存。
将以不同品种大豆根样以及根际土样DNA为模板扩增的18S Rrna(V1+V2)区PCR扩增产物用于DGGE分析。DGGE反应条件为:8%丙烯酰胺为变性剂,电泳温度为60℃,电压为130 V,电泳时间为10 h,变性梯度范围为40%~65%(100%的变性剂为40%甲酰胺,7M尿素),银染15 min。
1.3.4菌群结构分析 研究菌群结构时,主要通过丰度(S)、优势度(P)以及Shannon-Winener指数(H)来表征菌群动态变化。丰度以及优势度主要通过Gel-Pro Analyzer 4.5(Media Cybernetics Company,USA)软件分析DGGE图谱得到。H指数的计算公式如下:

s为DGGE图谱中,每个泳道中所包含的条带数。Pi为第i个条带的面积占该泳道所有条带总面积的百分数。
1.4基因测序以及数据分析
分析不同大豆品种根样以及根际土样DGGE图谱,选取其共有的和特有的条带进行割胶以及PCR扩增。扩增产物进行DGGE电泳,验证条带单一性。验证成功后,将胶回收扩增后的PCR产物连接至pGMT Easy(Tiangen,GER)载体,并转化到E.coli DH5α (AS,CHN)感受态细胞中,吸取符合DGGE电泳检测要求的阳性克隆菌液1.0 mL,送至上海英骏生物工程有限公司进行序列测定。测序结果提交至GenBank 数 据 库 ,并 通 过 NCBI(NationalCenterfor Biotechnology Information)数据库进行BLAST比对,得到相关种属信息。通过MEGA 5.0(Arizona State University,USA)进行系统发育树的构建。通过Quantity One 4.6.0(BIO-RAD,Hercules,CA,USA)进行聚类分析。通过SPSS 16.0进行统计学分析。
图1 KF16根样根腐病病原菌18S rRNA(V1+V2)区扩增结果
图2 KF16根际土壤根腐病病原菌18S rRNA(V1+V2)区扩增结果
2 结果与分析
2.1不同品种大豆根样以及根际土样真菌18S rRNA (V1+V2)区扩增结果
不同品种大豆根样以及根际土样总DNA通过引物序列(NS1/FungGC)进行18S rRNA(V1+V2)区扩增,扩增产物进行1%琼脂糖凝胶电泳检测。以KF16为例,扩增产物在430 bp处出现目的条带(图1、2)。说明KF16根样以及根际土样真菌DNA提取效果较好,可进行下一步分析。
2.2不同品种大豆根样以及根际土样根腐病病原菌DGGE图谱分析
通过分析3个大豆品种不同连作年限、不同生育时期根样以及根际土样根腐病病原菌18S rRNA(V1+ V2)进行DGGE后图谱(图3、4、5、6、7、8),发现不同试验样品根腐病病原菌丰度以及多样性存在差异(P<0.05),3个大豆品种不同生育时期对照组根腐病病原菌丰度以及多样性存在显著差异(P<0.05),并且对照组根腐病病原菌丰度以及多样性随着连作年限的延长而升高;试验组根腐病病原菌丰度以及多样性显著低于对照组(P<0.05);根样与根际土样根腐病病原菌丰度以及多样性也存在差异(P<0.05);说明接种F. mosseae、大豆品种、连作年限以及病原菌栖息环境(根系和土壤)均对大豆根腐病病原菌菌群结构产生影响。
通过对不同试验样品DGGE图谱进行分析与比较,选取DGGE图谱中13条条带进行割胶测序,测序结果见表1。其中,M3代表终极腐霉菌(Pythium ultimum),相似度 97%;M4代表立枯丝核菌(Rhizoctonia solani),相似度99%;M8代表格孢腔菌属(Pleosporales sp.),相似度99%;M11代表镰孢菌属(Fusarium sp.),相似度99%,M12代表尖孢镰刀菌(Fusarium oxysporum),其他7条带均为未培养真菌,相似度均在96%以上。利用Gel-Pro Analyzer 4.5(Media Cybernetics Company,USA)软件对DGGE图谱中条带进行优势度分析,结果表明大豆根腐病病原菌优势菌群为Pythium ultimum以及Fusarium sp.。
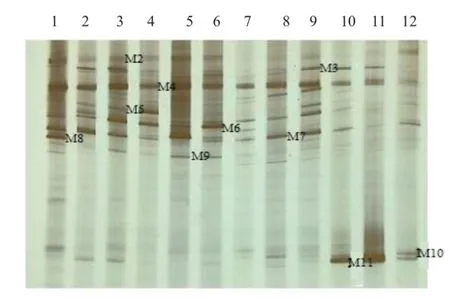
图3 KF16苗期根样以及根际土样根腐病病原菌DGGE图谱
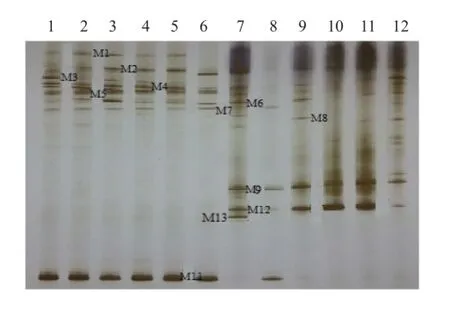
图4 KF16分枝期根样以及根际土样根腐病病原菌DGGE图谱

图5 HN44苗期根样以及根际土样根腐病病原菌DGGE图谱
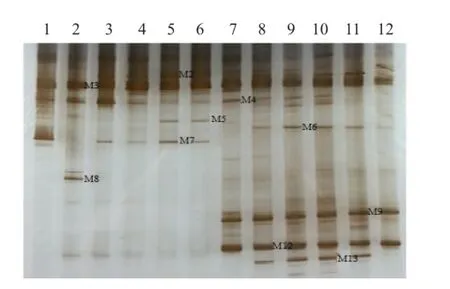
图6 HN44分枝期根样以及根际土样根腐病病原菌DGGE图谱
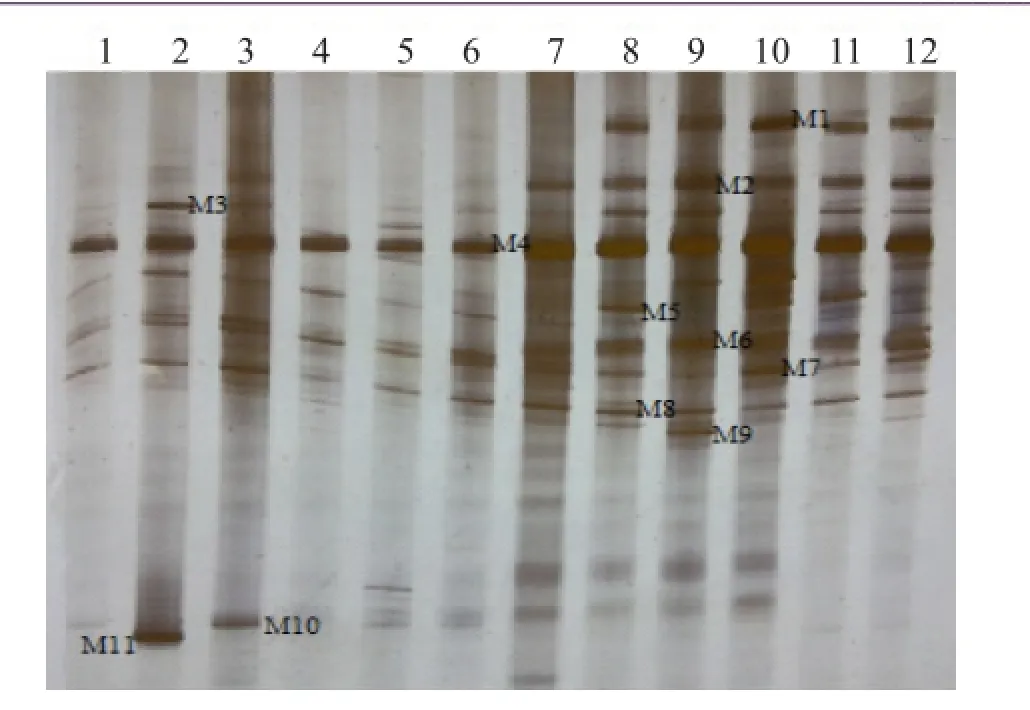
图7 HN48苗期根样以及根际土样根腐病病原菌DGGE图谱
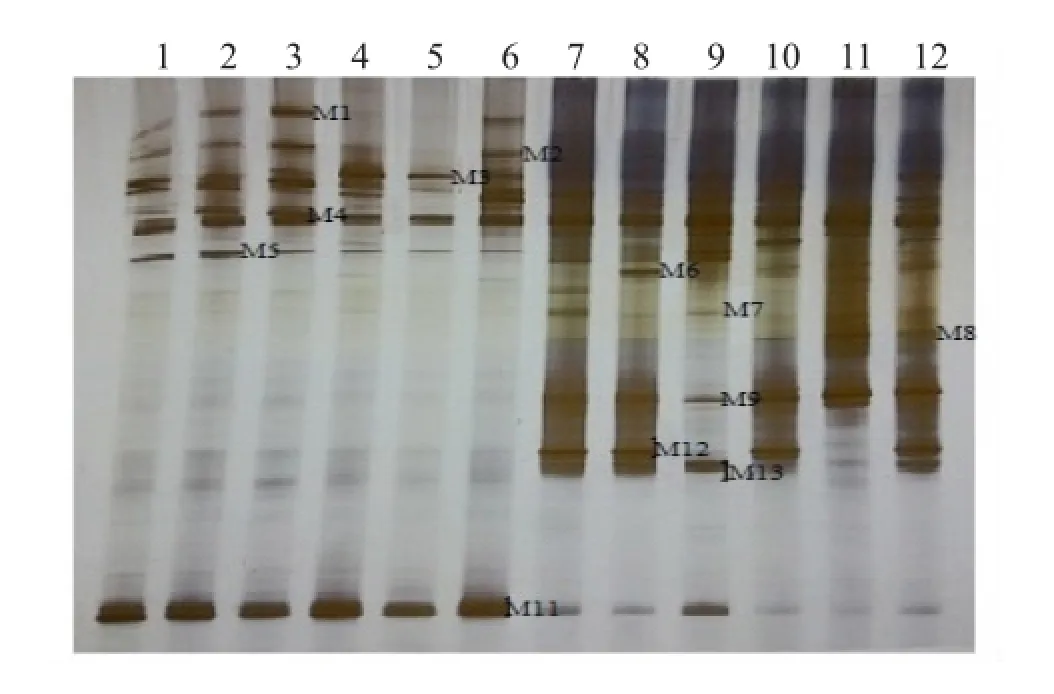
图8 HN48分枝期根样以及根际土样根腐病病原菌DGGE图谱

表1 13条序列的BLAST结果
2.3系统发育树
为了更加直观地显示出13条DGGE序列与GenBank数据库中已知微生物之间的亲缘关系以及系统地位,利用MEGA 5.0(Arizona State University,USA)进行系统发育树构建(图9)。
从系统发育树图可知,13条DGGE序列分为3组菌群:第I组包括尖孢镰刀菌(Fusarium oxysporum),镰孢菌属(Fusarium sp.)、立枯丝核菌(Rhizoctonia solani)以及未培养真菌,第II组包括终极腐霉菌(Pythium ultimum)以及未培养真菌(Uncultured fungus),第III组包括格孢腔菌属(Pleosporales sp.)以及未培养的真菌(Unculturedfungus)。 其 中 ,Pythiumultimum、 Fusarium oxysporum以及Fusarium sp.为大豆根腐病病原优势菌群。
2.4接种F.mosseae对连作大豆根样以及根际土样根腐病病原菌菌群结构的影响
相同大豆品种、不同生育时期根样以及根际土样根腐病病原菌丰度进行统计分析,以HN48为例(其他品种均具有相同规律)。HN48苗期对照组根样以及根际土样根腐病病原菌丰度显著高于分枝期对照组(P<0.05)。同时,根腐病病原菌丰度在根样以及根际土样之间也存在差异(P<0.05),并且对照组根腐病病原菌丰度值随着连作年限的增加而升高。HN48通过接种F.mosseae,试验组根腐病病原菌丰度显著低于对照组(P<0.05),并且随着大豆生育期的延长,分枝期试验组根腐病病原菌丰度显著低于苗期试验组(P<0.05)(表2)。相同生育时期,不同大豆品种根样以及根际土样根腐病病原菌丰度进行统计分析(表2)。KF16苗期以及分枝期对照组根样以及根际土样根腐病病原菌丰度显著高于HN44、HN48(P<0.05)。3个大豆品种接种F.mosseae后,HN44苗期以及分枝期试验组根腐病病原菌丰度显著低于KF16、HN48(P<0.05)。分析上述结果表明:接种F.mosseae对根腐病病原菌生长以及分布产生影响,并且F.mosseae的防治效果具有品种选择性。
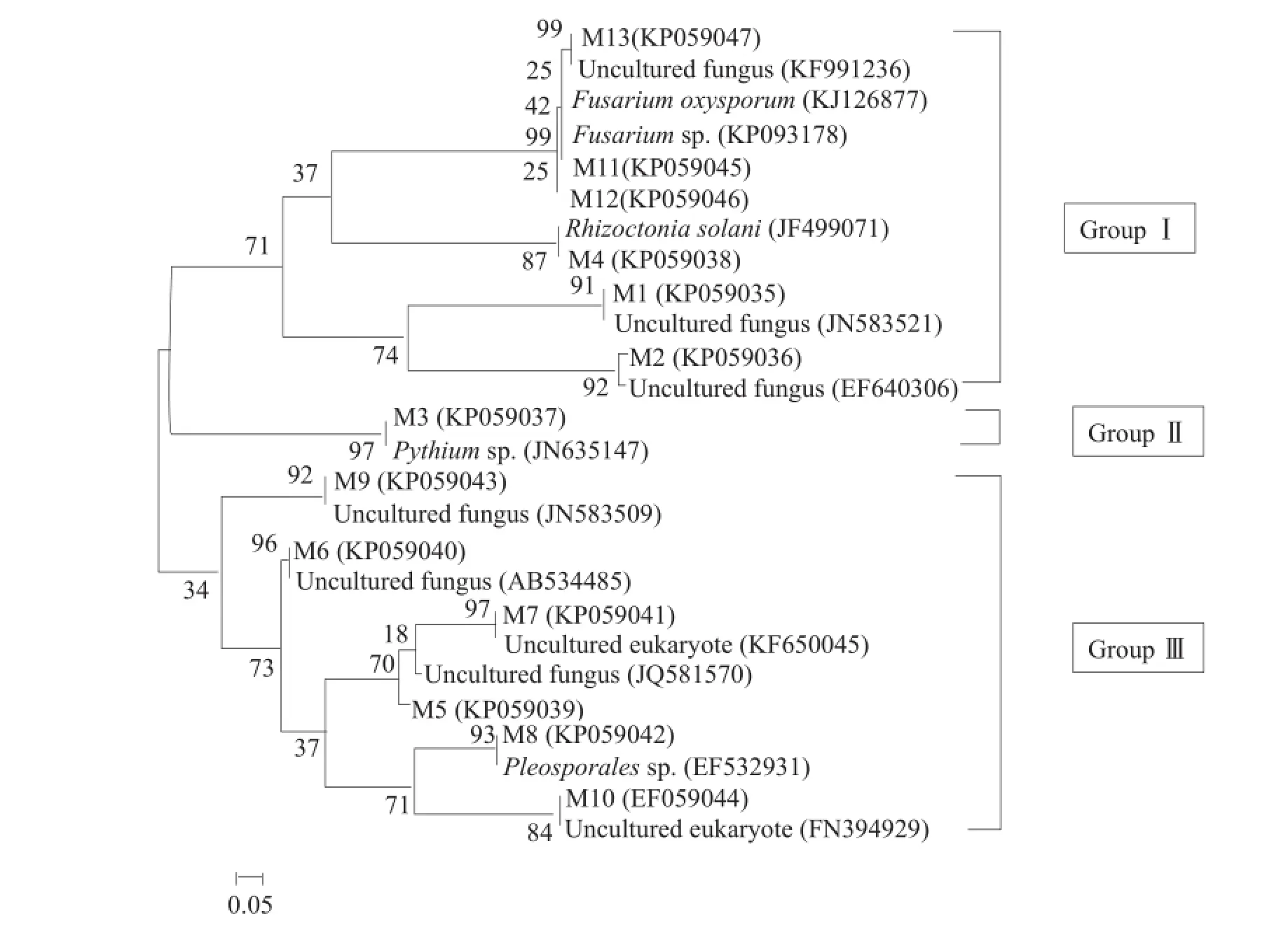
图9 13条DGGE序列与其近缘物种的系统发育树(节点上的数字表示1000自展后的数值)

表2 不同大豆品种不同生育时期根腐病病原菌丰度方差分析
相同大豆品种、不同生育时期根样以及根际土样根腐病病原菌多样性进行统计分析,以KF16为例(其他品种均具有此规律)。KF16苗期对照组根样以及根际土样根腐病病原菌多样性指数显著高于分枝期(P<0.05)。同时,根样与根际土样之间根腐病病原菌多样性指数也存在差异(P<0.05)。并且根腐病病原菌多样性指数随着连作年限的增加而升高。KF16试验组通过接种F.mosseae,试验组根腐病病原菌多样性指数显著低于对照组,并且随着大豆生育期的延长,分枝期试验组根腐病病原菌多样性指数显著低于苗期试验组(P<0.05)(表3)。相同生育时期、不同大豆品种根样以及根际土样根腐病病原菌多样性进行统计分析。苗期时,KF16对照组根样根腐病病原菌多样性指数显著高于HN44、HN48(P<0.05)。HN48对照组根际土样根腐病病原菌多样性指数显著高于KF16、HN44(P<0.05)。同时,根样与根际土样之间根腐病病原菌多样性指数也存在差异(P<0.05),并且根腐病病原菌多样性指数随着连作年限的增加而升高。3个大豆品种接种F.mosseae后,试验组根腐病病原菌多样性指数显著低于对照组(P<0.05)。KF16试验组根腐病病原菌多样性指数显著低于HN44、HN48(P<0.05)。分枝期时,HN44对照组根样以及根际土样根腐病病原菌多样性显著高于KE16、HN48(P<0.05)。同时,根样与根际土样之间根腐病病原菌多样性指数也存在差异(P<0.05)。3个大豆品种接种F.mosseae后,试验组根腐病病原菌多样性指数显著低于对照组(P<0.05)。HN48试验组根腐病病原菌多样性指数显著低于KF16、HN44(P<0.05)。分析上述结果表明:接种F.mosseae对根腐病病原菌多样性产生影响。并且不同品种接种F.mosseae后,根腐病病原菌防治效果具有差异。

表3 不同大豆品种不用生育时期根腐病病原菌多样性方差分析
通过Quantity One 4.6.0(BIO-RAD,Hercules,CA,USA)对3个大豆品种根样以及根际土样根腐病病原菌DGGE图谱进行聚类分析(图10a、b、c)。通过分析KF16根样以及根际土样聚类分析结果(图10a),表明根腐病病原菌菌群被分成15组:组I包括分枝期土样(试验组SL2),组II包括分枝期土样(试验组SL0和SL1),组III包括分枝期土样(对照组SL0和SL2),组IV包括分枝期根样(对照组RL1)和分枝期土样(对照组SL1),组V包括分枝期根样(对照组RL0),组VI包括分枝期根样(试验组RL2),组VII包括分枝期根样(试验组RL0),组VIII包括分枝期根样(试验组RL1和对照组RL2),组IX包括苗期土样(试验组SL1和SL2),组X包括苗期土样(试验组SL0),组XI包括苗期土样(对照组SL2)和苗期根样(对照组RL1),组XII包括苗期根样(对照组RL0和RL2),组XIII包括苗期土样(对照组SL0和SL1),组XIV包括苗期根样(试验组RLI),组XV包括苗期根样(试验组RL0和RL2)。图10b、10c分析方法同图10a,不再进行描述。聚类分析结果表明不同品种、不同样品间根腐病病原菌菌群结构存在差异,并进一步验证接种F.mosseae、大豆品种、连作年限以及病原菌栖息环境(根系以及土壤)均对大豆根腐病病原菌产生以及分布产生影响。

图10 不同大豆品种不同生育时期根腐病病原菌DGGE图谱聚类分析
3 讨论
AM真菌侵染维管植物根系后,通过形成庞大的菌丝结构,促进植物根系对水分、矿质元素吸收以及运输,进而提高植物对营养物质的吸收,促进植株生长。近年来,AM真菌被广泛应用于植物-微生物共生体增强植物抗逆性研究中,并取得了丰硕的研究成果[17-19]。但是,国内外研究人员很少报道接种AM菌剂后,其对植物根系以及根际土壤中病原菌菌群结构的影响[20-21]。因此,本试验通过接种F.mosseae,研究其对3个大豆品种根系以及根际土壤中根腐病病原菌菌群结构的影响。
根腐病是一种世界性作物病害,严重威胁农作物产量以及品质。同时,PCR-DGGE技术是目前研究微生物群落结构的常用技术,并且国内外学者已将其应用于根腐病病原菌菌群结构研究中[22-23]。试验过程中发现随着连作年限的增加,3个大豆品种对照组根样以及根际土壤中根腐病病原菌丰富度以及多样性指数逐年升高,并且每个大豆品种根样与根际土样之间根腐病病原菌丰富度以及多样性指数也存在差异,说明大豆连作造成土壤理化性质改变,导致大豆根腐病病原菌丰富度以及多样性指数逐年升高[24]。同时也说明根腐病病原菌对栖息环境有一定的选择性。大豆苗期根腐病病原菌丰富度以及多样性指数显著高于分枝期,说明随着大豆生育进程的延长,大豆防御性蛋白以及次生代谢产物逐渐表达以及分泌,有利于大豆防御性体系的形成以及建立,进而提高大豆抵御根腐病病原菌侵染的能力[25-26]。
试验通过向3个大豆品种盆栽土壤中接种F.mosseae,研究其对不同大豆品种不同连作年限根腐病病原菌菌群结构的影响。试验结果表明3个大豆品种试验组根样以及根际土样根腐病病原菌丰富度以及多样性指数显著低于对照组。说明接种F.mosseae后,F.mosseae通过侵染大豆根系形成庞大的菌丝体结构,提高大豆根系吸收水分以及矿质营养的面积以及能力,进而促进大豆根系防御性蛋白以及次级代谢产物表达以及分泌,有利于加快大豆防御性体系的形成以及建立,最终提高大豆抵御根腐病病原菌侵染能力[20]。
试验选择黑龙江地区较具代表性3个大豆品种进行研究,研究发现3个大豆品种对照组根腐病病原菌丰度值以及多样性指数存在差异。并且接种F.mosseae后,3个大豆品种根腐病病原菌防治效果也存在差异显著。说明3个大豆品种遗传性质对于根腐病防治具有一定影响,同时也说明F.mosseae对于根腐病防治效果具有一定的品种选择性[27]。聚类分析结果进一步验证了接种F.mosseae、大豆品种、连作年限以及病原菌栖息环境(根系和土壤)均对连作大豆根腐病病原菌产生以及分布产生影响。
4 结论
不同连作年限不同大豆品种接种F.mosseae后,试验组根样以及根际土样根腐病病原菌丰度以及多样性指数显著低于对照组,并且接种F.mosseae对不同大豆品种防治效果存在差异;聚类分析结果进一步验证了接种F.mosseae对连作大豆根样以及根际土样根腐病病原菌产生以及分布产生影响。因此,本项研究对于应用生物防治方法——AM菌剂于连作大豆根腐病防治做出了前期的探索以及实践,并为AM菌剂应用于缓解大豆连作障碍提供理论依据。
[1]Hartman G L,West E D,Herman T K.Crops that feed the world 2. Soybean-worldwide production,use,and constraints caused by pathogens and pests[J].Food Security,2011,3(1):5-17.
[2]Han J,Jie W G,Cai B Y.Effect of phosphorus on the composition and accumulation of 7S and 11S globulin subunits during seed development of three soybean varieties[J].Journal Soil Science and Plant Nutrition,2014,14(4):769-782.
[3]Chen M N,Li X,Yang Q L.Dynamic succession of soil bacterial communityduringcontinuouscroppingofpeanut (Arachishypogaea L.)[J].PLoS One,2014,9(7):e101355.doi: 10.1371/journal.pone.0101355.
[4] 魏巍,许艳丽,朱琳,等.长期连作对大豆根际土壤镰孢菌种群的影响[J].应用生态学报,2014,25(2):497-504.
[5] Xiong W,Zhao Q Y,Zhao J,et al.Different continuous cropping spans significantly affect microbial community membership and structureinavanilla-grownsoilasrevealedbydeep pysosequencing[J].Microbial Ecology,2015,70(1):1-10.
[6]Reinhardt D.Programming good relations development of the arbuscular mycorrhial symbiosis[J].Current Opinion in Plant Biology,2007,10(1):98-105.
[7]Walder F,Niemann H,Natarajan M,et al.Mycorrhizal networks: common goods of plants shared under unequal terms of trade[J]. Plant Physiology,2012,159(2):789-797.
[8]Wei Y,Hou H,Li J N,et al.Molecular diversity of arbuscular mycorrhizalfungiassociatedwithanMnhyperaccumulator-PhytolaccaAmericana,inMnminingarea[J]. Applied Soil Ecology,2014,82:11-17.
[9]Lehmann A,Kathryn-Barto E,Powell J R,et al.Mycorrhizal responsiveness trends in annual crop plants and their wild relativesa meta-analysis on studies from 1981 to 2010[J].Plant Soil,2012,355(1-2):231-250.
[10]Zhang H S,Li G,Qin F F,et al.Castor bean growth and rhizosphere soil property response to different proportions of arbuscularmycorrhizalandphosphate-solubilizingfungi[J]. Ecological Research,2014,29(2):181-190.
[11]Zhang F G,Zhu Z,Yang X M,et al.Trichodermaharzianum T-E5 significantly affects cucumber root exudates and fungal community in the cucumber rhizosphere[J].Applied Soil Ecology,2013,72:41-48.
[12]Huang Y M,Srivastava A K,Zou Y N,et al.Mycorrhizal-induced calmodulin mediated changes in antioxidant enzymes and growth response of drought-stressed trifoliate orange[J].Frontiers in Microbiology,2014,5:682-682.
[13]罗巧玉,王晓娟,林双双,等.AM真菌在植物病虫害防治中的作用机制[J].生态学报,2013,33(19):5997-6005.
[14]Ortiz N,Armada E,Duque E,et al.Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought toleranceundernaturalsoilconditions:effectivenessof autochthonous or allochthonous strains[J].Journal of Plant Physiology,2015,174:87-96.
[15]Gomez-Bellot M J,Ortuno M F,Nortes PA,et al.Protective effects of Glomusiranicum var.tenuihypharum on soil and Viburnum tinus plants irrigated with treated wastewater under field conditions[J]. Mycorrhiza,2014,25(5):1-11.
[16]Ei-Faiz A,Duponnois R,Winterton P,et al.Effect of different amendments on growing of Canna indica L.inoculated with AMF on mining substrate[J].International Journal of Phytoremediation,2015,17(5):503-513.
[17]樊宇红,凌宏文,朴河春.桑树(Morus alba)与丛枝菌根的共生对重金属元素吸收的影响[J].生态环境学报,2014,23(3):477-484.
[18]杨海霞,李士美,郭绍霞.丛枝菌根真菌对紫薇耐盐性的影响[J].植物生理学报,2014,50(9):1379-1386.
[19]周宝利,郑继东,毕晓华,等.丛枝菌根真菌对茄子黄萎病的防治效果和茄子植株生长的影响[J].生态学杂志,2015,34(4):1026-1030.
[20]Qian L,Yu W J,Cui J Q,et al.Funnelifomis mosseae affects the root rot pathogen Fusarium oxysporum in soybeans[J].Acta Agriculturae Scandinavica,Section B-Soil&Plant Science,2015,64 (4):321-321.
[21]Jie W G,Bai L,Yu W J,et al.Analysis of interspecific relationships between Funeliformis mosseae and Fusarium oxysporum in the continuous cropping of soybean rhizosphere soil during the branching period[J].Biocontrol Science and Technology,2015,25 (9):1036-1051.
[22]Yergeau E,Filion M A.PCR-denaturing gradient electrophoresis approach to assess Fusarium diversity in asparagus[J].Journal of Microbiological Methods,2005,60:143-154.
[23]范雪露,崔佳琦,蔡柏岩.接种摩西柄囊霉对连作大豆苗期根腐病病原真菌多样性的影响[J].中国农学通报,2014,30(31):272-279.
[24]魏巍,许艳丽,张思佳,等.大豆根腐病原镰孢菌种群多样性DGGE分析及其致病性研究[J].植物病理学报,2013,43(5):500-508.
[25]鞠会艳,韩丽梅,王树起,等.连作大豆根系分泌物对根腐病病原真菌的化感作用[J].应用生态学报,2002,13(6):723-727.
[26]马丹炜,王亚男,王煜,等.化感胁迫诱导植物细胞损伤研究进展[J].生态学报,2015,35(5):1640-1645.
[27]Jie W G,Liu X R,Cai B Y.Diversity of rhizosphere soil arbuscular mycorrhizal fungi in various soybean cultivars under different continuous cropping regimes[J].PLoS One,2013,8(8):e72898.
Effect of Inoculated Funneliformis mosseae on Root Rot Pathogen Flora Structure in Soybean Continuous Cropping
Cui Jiaqi1,2,Fan Xuelu1,2,Cai Baiyan1,2
(1College of Life Sciences,Heilongjiang University,Harbin 150080,Heilongjiang,China;
2Key Laboratory of Microbiology of General Institutes of Higher Education,Harbin 150080,Heilongjiang,China)
In this study,the bioremediation method was applied to preventing the root rot in soybean continuous cropping in order to reduce the occurrence of root rot and enhance the quality of soybean.Three soybean varieties were selected and inoculated with Funneliformis mosseae in the trial.The root rot pathogen community structure of the root system and rhizosphere soil in soybean continuous cropping were measured by PCR-DGGE.The results showed that the abundance and diversity index of root rot pathogen of the soybean root system and rhizosphere soil samples in the treatment groups were significantly lower than that in the control groups after inoculated with F.mosseae,and F.mosseae had different effect on different soybean cultivars.The results of cluster analysis also verified that F.mosseae affected the occurrence and distribution of root rot pathogen in soybean continuous cropping.
Root Rot;F.mosseae;Soybean Continuous Cropping;Flora Structure
S435.651
A论文编号:cjas15120018
国家自然科学基金面上项目“摩西管柄囊霉(Funneliformis mosseae)抑制连作大豆根腐病发病机理的研究”(31570487)。
崔佳琦,男,1989年出生,黑龙江绥化人,在读硕士,研究方向为微生物生态学。通信地址:150080黑龙江省哈尔滨市南岗区学府路74号黑龙江大学生命科学学院,E-mail:cuijiaqi2015@126.com。
蔡柏岩,男,1968年出生,黑龙江哈尔滨人,教授,博士后,研究方向为微生物生态学。通信地址:150080黑龙江省哈尔滨市南岗区学府路74号黑龙江大学生命科学学院,Tel:0451-86626691,E-mail:caibaiyan@126.com。
2015-12-24,
2016-02-24。
猜你喜欢
河南水利与南水北调(2023年7期)2023-08-29 02:19:54
环境保护与循环经济(2022年7期)2023-01-03 09:31:55
今日农业(2022年15期)2022-09-20 06:54:16
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
今日农业(2021年21期)2021-11-26 05:07:00
现代畜牧科技(2021年9期)2021-10-13 06:38:54
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
治淮(2018年6期)2018-01-30 11:42:44
中国蔬菜(2016年8期)2017-01-15 14:23:38
上海蔬菜(2016年5期)2016-02-28 13:18:10
