
峡东地区上埃迪卡拉系岩石、生物、层序和碳同位素地层及其年代学意义
2016-09-09 08:15陈孝红张保民
华南地质 2016年2期
陈孝红,周 鹏,张保民,危 凯,张 淼
(中国地质调查局武汉地质调查中心,武汉430205)
·简讯·
峡东地区上埃迪卡拉系岩石、生物、层序和碳同位素地层及其年代学意义
陈孝红,周鹏,张保民,危凯,张淼
(中国地质调查局武汉地质调查中心,武汉430205)
本文系统研究了峡东灯影峡上埃迪卡拉系陡山沱组上部至灯影组石板滩段及其区内与之同期不同相区(或古地理部位)地层岩石、生物、层序和碳同位素地层,结果发现峡东埃迪卡拉系陡山沱组—灯影组界线附近地层横向上存在台地相、高陡边缘台地边缘相和盆地相多种沉积相类型,灯影组的底界和内部划分明显受沉积相的制约,以白云岩出现为标志的灯影组底界是一个明显的穿时面。与灯影组中部存在的两次海进—海退旋回相对应不仅发生了两次δ13C的上升和下降,而且发生了两次重要的生物辐射事件,在石板滩段下部和上部分别出现了具有全球广泛对比意义的叶状埃迪卡拉动物和管状Cloudiniids动物。据此可以将峡东灯影组划分出两个可以进行全球对比的年代地层单位泗溪阶和龙灯溪阶。由于龙灯溪阶上部,继Cloudiniids消失之后,小壳化石出现之前发生了δ13C的震荡下降,并在灯影组白马沱段底部出现了最小δ13C达到-12‰的碳同位素强烈负偏离。因此,生物和碳同位素组成变化特点及其全球对比显示长江三峡地区寒武系的底界应大致与灯影峡剖面上灯影组石板滩段与白马沱段界线一致。
灯影组;稳定碳同位素;埃迪卡拉动物;管状动物;埃迪卡拉系-寒武系界线
CHEN X H,ZHOU P,ZHANG B M,WEI K,ZHANG M.
Geology and Mineral Resources of South China,2016,32(2):87-105.
虽然峡东地区作为中国南方埃迪卡拉系划分对比的标准地区,具有悠久的研究历史[1],但迄今为止,有关区内上埃迪卡拉系的内部划分,特别是灯影组的岩石地层、生物地层和年代地层以及稳定碳、氧同位素地层等研究方面尚有不少值得进一步研究和完善的地方。在岩石地层方面,由于在灯影峡层型剖面上的灯影组下部并未见到陡山沱组层型剖面上陡山沱组顶部发育的黑色炭质页岩层段(第四段)[2]。加之,灯影峡剖面上的陡山沱组出露不佳,地层序列保存不完整,因此,陡山沱组层型剖面上所见的陡山沱组第四段在灯影峡剖面上是否存在,或者相变为含硅质条带的薄层状灰岩、白云岩仍存在不同的看法[3]。在生物地层和年代地层研究方面,虽然灯影组石板滩段内部产丰富的遗迹化石[4-5]、埃迪卡拉型动物化石和文德生物化石[6-8]以及管状动物化石[9-10]等,具有利用生物化石开展全球对比的可能,但这些生物化石的年代地层意义仍未揭示,有关埃迪卡拉系内部分统、建阶的方案和年代地层单位界线层型剖面和点的确定仍然处于研究与探讨之中[11-12]。虽然稳定碳同位素地层在埃迪卡拉系年代地层单位划分对比中的作用日益受到重视,且前人对灯影峡灯影组进行过多次稳定碳同位素地层学研究[13-15]。但早期的稳定碳同位素地层研究样品间距太大,后期研究过程中,由于葛洲坝修建所引起的长江水位上升,影响了剖面的通行条件,不利于样品的连续采集和高分辨率碳同位素地层的研究。近年来,随着三峡旅游公路的修建,灯影峡剖面重新被揭露,从而不仅有利于剖面沉积序列的重新观察,而且为连续采集新鲜的碳同位素样品提供了有利条件。为此,本文拟通过灯影峡新近旅游公路修建而重新揭露的上埃迪卡拉系陡山沱组上部—灯影组下部剖面的重新测量及其岩石、生物、层序和碳同位素化学地层的重新调查与综合研究,重新厘定灯影组底界的含义,分析探讨灯影组内部可能的年代地层单位划分对比界线,进而为我国埃迪卡拉纪年代地层系统的建立提供新的依据。
1 地质概况
长江三峡地区在大地构造位置上位于扬子地台北部。区内埃迪卡拉系与下伏南华系、上覆早古生代地层呈环带状连续出露在黄陵穹窿周缘(图1)。区内埃迪卡拉系露头良好,沉积连续,自下而上划分为陡山沱组和灯影组。其中灯影组的岩石组合在纵向分布上具有明显的三分性特征,可进一步划分为上部白马沱段、下部蛤蟆井段白云岩和中部石板滩段灰岩,横向三段厚度变化明显,灯影峡往东往北,至晓峰、漳村坪一带灯影组下部和上部白云岩的厚度逐步增大,中部灰岩段的厚度变小,往西往南,至秭归青林口、庙河一带则正好相反,灯影组主要相变为薄层状灰岩,上部和下部白云岩,特别是下部蛤蟆井段白云岩厚度明显变薄至不到1 m的厚度[2]。
2 材料和方法
本次研究主要针对上埃迪卡拉系陡山沱组第四段-灯影组石板滩段在岩石地层、生物地层、年代地层和碳同位素地层研究的不足和问题,有目的地选择峡东不同古地理部位灯影组的不同层、段开展目的不同的调查研究工作。主要包括:1)以修订和完善灯影峡灯影组碳、氧同位素记录为目的所开展的灯影峡剖面陡山沱组上部至灯影组石板滩段剖面实测和碳同位素样品的采集与分析。2)针对灯影峡灯影组底界的确定和区域横向对比,对宜昌李家院子和黄牛岩等地灯影组-陡山沱组界线地层进行实测和碳、氧同位素样品的采集和分析。3)针对灯影峡灯影组生物地层和年代地层划分对比问题,重点补充开展宜昌黄牛岩灯影组埃迪卡拉型动物化石和遗迹化石的采集和研究以及李家院子灯影组上部的管状动物化石和碳、氧同位素样品的系统采集、分析和研究(图1)。
灯影峡上埃迪卡拉系剖面的测量采用导线法进行,其余剖面直接用钢卷尺测量。同位素样品尽量采集没有遭受构造影响的新鲜碳酸盐岩,对于单层厚度较大,岩性变化不明显的层段,样品最大间距控制在5 m以内;对于单层岩层较薄,岩性变化明显的层段,样品间隔适当加密。但对本次发现的碳同位素组成具有明显异常的灯影组白马沱段下部第51层和52层下部进行了二次加密采样和测试。所采集岩石样品在室内进行切片观察,选取新鲜和未受方解石脉改造破坏区域采用牙钻碾磨取样,以备送样测试。考虑到灯影峡和黄牛岩灯影组碳氧同位素组成变化以往研究单位取得的进展[13-15],本次将采自黄牛岩陡山沱组第三段至灯影组石板滩段的样品交由中国地质调查局武汉地质调查中心测定,灯影峡灯影组蛤蟆井和石板滩段的样品送中国科学院广州地球化学研究所同位素地球化学重点实验室进行测定,其余剖面的样品,包括在灯影峡白马沱段下部第51层和52层下部加密采集的重复测试样品和李家院子剖面采集的样品交由中国地质大学(武汉)地质过程与矿产资源重点实验室测试完成。上述三个单位的同位素比值测试均在MAT-251进行,中国地质大学(武汉)地质过程与矿产资源重点实验室采用的参考标准为Vienna Pee Dee组的箭石(V-PDB),其余两个单位的参考标准为Pee Dee组的箭石(PDB)。重复分析的结果表明碳酸盐岩13C的分析精度均为±0.1‰。

图1 峡东地区地质简图与剖面分布位置示意图Fig.1 Generalized geological map ofthe eastern Yangtze Gorges
3 结果
3.1宜昌灯影峡剖面
宜昌莲沱灯影峡是灯影组的命名剖面,不仅在中国南方埃迪卡拉系是多重划分对比的标准剖面,同时也是最早和较多地研究灯影组碳同位素地层特征的剖面之一[1,13,16]。本次研究集中在对最近新开旅游公路重新揭露出的灯影组中下部蛤蟆井段、石板滩段。
重新揭露的灯影峡灯影组蛤蟆井段和石板滩段地层出露良好,层序清楚。其中,灯影组下伏陡山沱组顶部为薄层状含硅质结核或条带白云岩、鲕粒白云岩。灯影组蛤蟆井段厚约140 m,下部以灰色厚层-块状白云岩、鲕白云岩砾屑白云岩发育为特征,上部主要为青灰色薄—厚层状泥晶白云岩,中部发育硅质条带,顶部夹角砾状白云岩。石板滩段底部10.3 m为硅质灰岩夹极薄层白云岩,下部90 m为黑色薄层状含硅磷质结核或条带灰岩,产丰富的文德带藻化石。上部为96 m的薄层状硅质白云岩、白云岩,向上夹多层厚层-块状白云岩、硅化白云岩。中部薄层白云岩中发育燧石结核,产管状动物化石[9]。上部上覆灯影组白马沱段为厚层块状粉晶白云岩(图2A)。

图2 峡东地区埃迪卡拉系灯影组岩石地层、层序地层和碳、氧同位素变化曲线及其对比Fig.2 Litho-and sequence stratigraphy,and δ13C(‰,VPDB)and δ18O(‰,VPDB)profiles ofthe Ediacaran DengyingFormation in the eastern Yangtze Gorges and their correlations
灯影组蛤蟆井段和石板滩段内部的碳同位素组成虽然没有出现区内陡山沱组内部那样的强烈振荡现象[4,17-18],但也不像Jianget al.[15]所指出的那样稳定。本次研究在灯影组蛤蟆井段所采集的41件碳氧同位素样品中的δ13C平均值为2.69‰。纵向上,δ13C在蛤蟆井段底部出现其最大值6.24之后震荡下降,至蛤蟆井段顶部出现该段δ13C最小值0.43下。
灯影组石板滩段50个样品的δ13C平均值为2.52‰。在石板滩段内部,伴随石板滩段底部地层沉积环境海平面的快速上升,地层中的δ13C也相应地从该段底部的1.25‰迅速增大至5.97‰。之后δ13C发生轻微下降,并在石板滩段下部薄层灰岩和中部薄层白云岩地层中稳定地变化在2‰~3‰之间,至石板滩段上部,伴随厚层块状白云岩夹层的出现,地层中的δ13C进一步下降至0~1之间。至灯影组白马沱段底部厚层块状粉晶白云岩夹中厚层微晶白云岩地层发生了碳同位素的明显负偏离,δ13C从该层底部1.02‰向上4 m迅速下降至-7‰,甚至更低(-12‰)。但在随后3 m的地层中的δ13C又从-5‰左右迅速上升到2‰附近(表1,图2A)。
3.2宜昌李家院子剖面
宜昌李家院子剖面灯影组下伏陡山沱组第四段厚约0.3~0.7 m,由下部发育水平层理的中—薄层状白云岩和上部发育滑塌构造或包卷层理的薄层白云岩夹薄层泥岩组成。与下伏陡山沱组第三段上部厚层状白云岩夹极薄层泥岩之间存在一个明显的凹凸不平面。灯影组蛤蟆井段厚约30 m,中下部以垮塌角砾白云岩发育为特征,上部为中—薄层状白云岩,发育大量的硅质条带或结核。垮塌角砾白云岩主要由砂屑白云岩和砾屑白云岩岩块和揉皱构造发育的白云岩组成。石板滩段厚约220 m,下部为薄层状夹中层状灰岩,含燧石结核或条带,发育滑塌构造。上部为中—薄层状灰岩夹白云质灰岩、白云岩,向上白云岩含量升高,产Cloudina等管状动物化石。顶部为薄—中厚层状白云岩(未见顶)(图2B,图3)。
剖面上碳同位素地层组成研究主要围绕产管状动物化石的灯影组石板滩段中—上部进行(表2,图2B)。样品采集和分析结果显示,以石板滩段中部白云质灰岩的出现为标志所划分的石板滩段下部靠近上部的 22个样品的的 δ13C平均值为2.827‰,最小 δ13C为 1.49‰,最大 δ13C为3.66‰。石板滩段中部68个样品的的δ13C平均值为2.24‰,最小δ13C为-0.65‰,最大δ13C为3.74‰。石板滩段上部灰岩夹薄层白云岩地层碳同位素组成存在强烈震荡的特点,22个样品的δ13C平均值为1.66‰,最小δ13C为-1.22‰,最大δ13C 为3.22‰。其上石板滩段顶部薄层白云岩中δ13C震荡升高至3‰附近之后趋于稳定,该段地层23个样品的δ13C平均值为2.85‰,最小δ13C为1.01‰,最大δ13C为4.03‰。
3.3宜昌黄牛岩剖面
黄牛岩陡山沱组上部—灯影组下部剖面沿头顶石东侧公路出露,露头良好。实测结果显示,陡山沱组四段为黑色页岩,上部夹灰岩透镜体,厚8.3 m。灯影组下部蛤蟆井段为厚22.1 m的薄—中厚层状白云岩,上部发育分选磨圆较差的同生角砾白云岩。灯影组蛤蟆井段—石板滩段之间接触界面不平整,上覆石板滩段底部为薄层状灰岩,向上薄层灰岩中发育滑塌构造,指示石板滩段下部地层沉积环境有从台地向台地边缘斜坡转变,水体不断加深的趋势。该地石板滩段未见顶,在第一个滑塌层之上,距离石板滩段底部约20 m的黑灰色极薄层泥晶—微晶灰岩间夹亮晶灰岩中产埃迪卡拉型动物化石Yangtziramulus zhangi以及遗迹化石等(图2E,图4C)。碳同位素组成测定结果表明,灯影组下部蛤蟆井段和石板滩段下部的碳同位素组成与灯影峡剖面同期地层的碳同位素组成相似,蛤蟆井段的δ13C有自下而上有先升后降的变化趋势,蛤蟆井段的δ13C从底部的0.92有向上逐步升高,至中部达到其最大值4.4‰,到蛤蟆井段与石板滩段界线附近又下降为1.18段,此后在石板滩段下部又迅速升高至5.66之后保持相对稳定(表3,图2E)。

表1 灯影峡剖面灯影组碳、氧同位素测试结果Table 1 δ13C and δ18O of samples from the Dengying Gorges section,Yichang
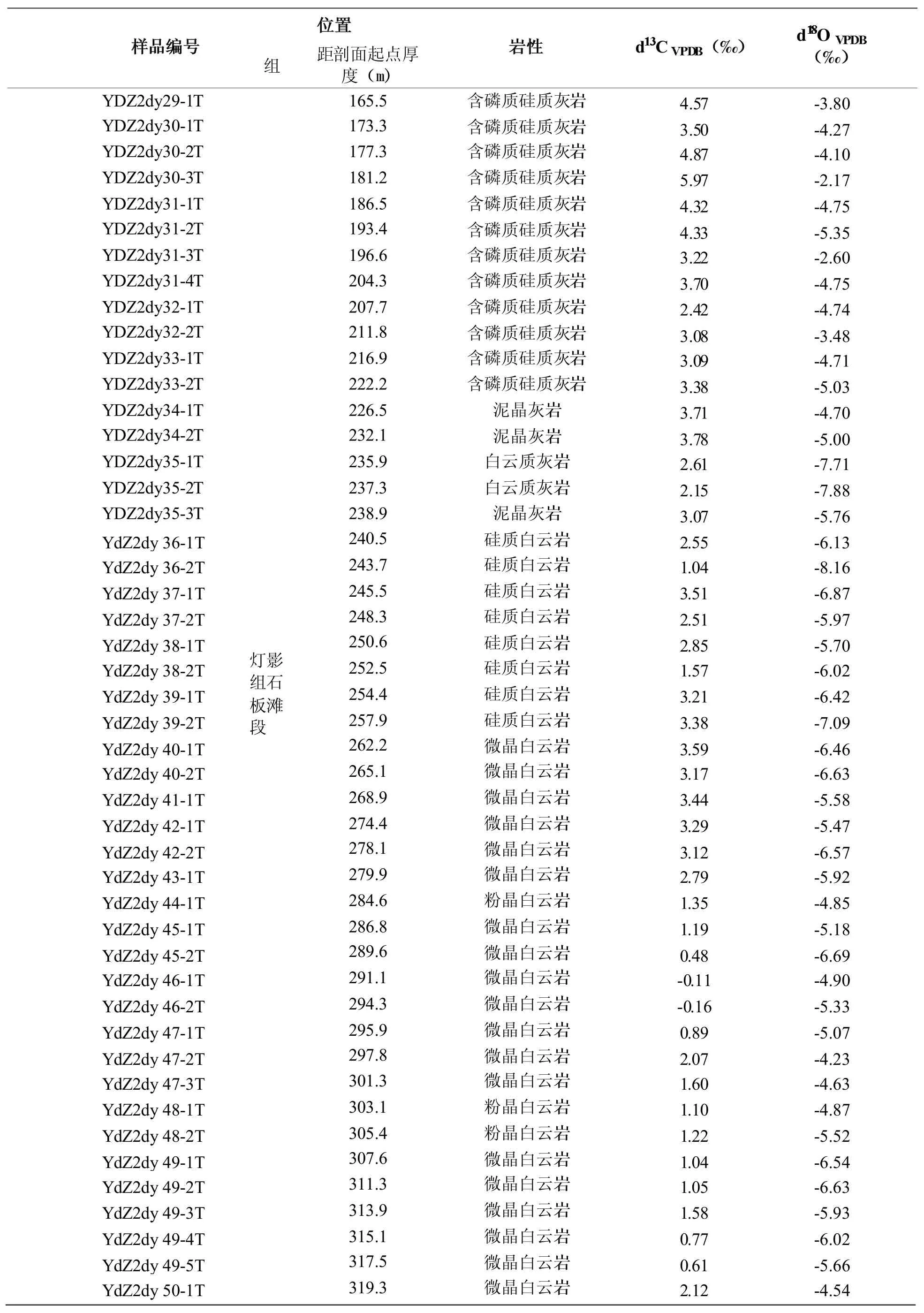
续表

续表
4 比较与讨论
4.1灯影组底界及其内部划分
宜昌黄牛岩、李家院子和灯影峡上埃迪卡拉系陡山沱组上部第四段至灯影组石板滩段地层剖面测量及其岩石、生物、和碳同位素研究结果显示峡东地区埃迪卡拉系陡山沱组三段和四段之间以及灯影组蛤蟆井段和石板滩段、石板滩段中部和下部以及石板滩段与白马沱段下部之间曾经发生过海平面的下降事件(图2)。其中与灯影组底界划分紧密相关的海平面升降旋回开始于陡山沱组第四段,至灯影组蛤蟆井段顶部结束。以此为基础,所获得的上述不同剖面灯影组底界地层的划分对比结果暗示区内陡山沱组—灯影组界线地层沉积时期已经具有明显的岩相古地理分异,灯影组底界划分标志与古地理部位相关,至少存在以下三种不同的类型(图4)。
第一种类型的灯影组底界以宜昌黄牛岩剖面为代表(图4C),主要分布在黄陵背斜南翼宜昌莲沱田家园子、黄牛岩、西翼秭归泗溪、青林口、庙河等地。这种类型的灯影组底界划分标志与陡山沱组标准剖面上灯影组底界划分标志基本一致,以灯影组蛤蟆井段底部薄—中厚层状白云岩的出现为标志与下伏陡山沱组上部第四段黑色炭质页岩相区分。该相带内的灯影组蛤蟆井段下部为薄—中厚层状白云岩,上部白云岩中发育帐篷构造和膏盐角砾白云岩。岩石组合序列指示上述地区灯影组/陡山沱组界线附近的沉积环境在海平面上升时期为深水盆地环境,晚期伴随海平面下降而转化为局限盆地。由于这些地区以盆地相为主,因此所对应的灯影组以石板滩段薄层灰岩发育为特点,灯影组下部蛤蟆井段和上部白马沱段白云岩不发育,厚度为几米至数十米不等。

表2 李家院子剖面灯影组碳、氧同位素测试结果Table 2 δ13C and δ18O of samples from the Lijiayuanzi section,Yichang

续表
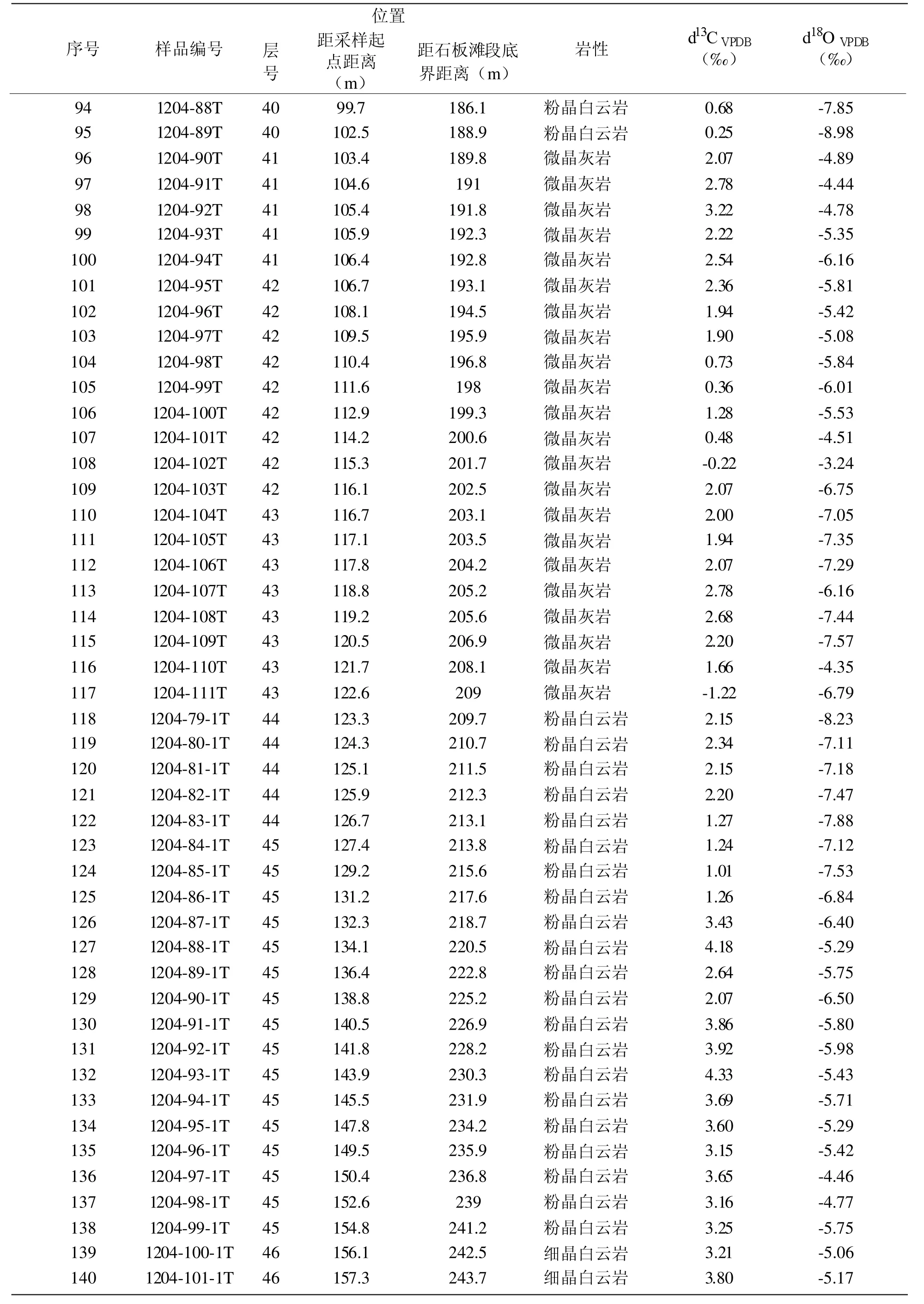
续表
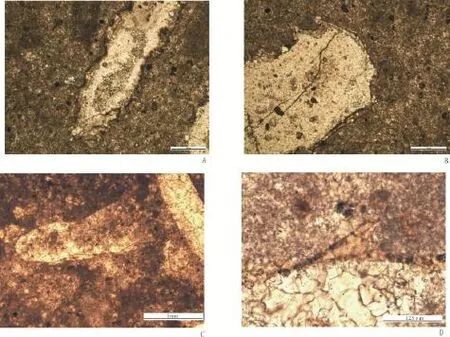
图3 宜昌李家院子灯影组石板滩段中部发育的两种不同的管状动物化石Fig.3 Twokinds ofdifferent tubular fossils in the middle part ofthe Shibantan member ofthe DengyingFormation at Lijiayuanzi,Yichang,Hubei China.

表3 宜昌黄牛岩剖面灯影组下部碳、氧同位素测试结果Table 3 δ13C and δ18O of samples from the Lijiayuanzi section,Yichang
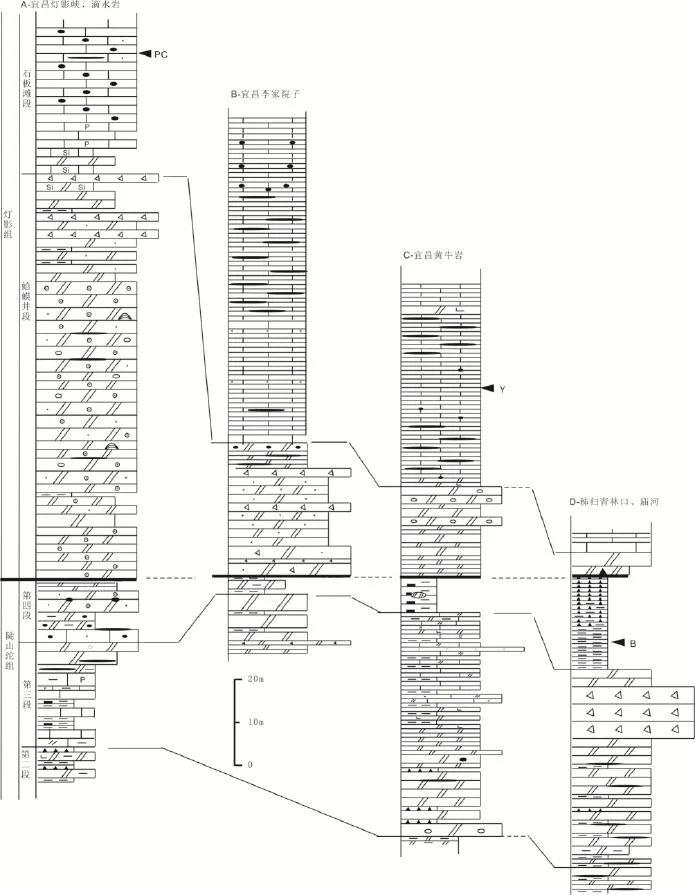
图4 峡东地区埃迪卡拉系陡山沱组-灯影组界线地层划分对比图(图例见图2)Fig.4 The subdivisions and correlations ofthe boundarybetween the Dengyingand DoushantuoFormations in the eastern Yangtze Gorges.(Legends for the logsee Fig.2)
第二种类型的灯影组底界以宜昌李家院子(仙人洞)剖面为代表(图4B)。该种类型的陡山沱组与灯影组之间以一层厚0.03 m的硅质岩、泥岩为标志,陡山沱组上部由发育水平层理的中—薄层状白云岩和发育滑塌构造或包卷层理的薄层白云岩夹薄层泥岩组成(图4B)。灯影组蛤蟆井段以垮塌角砾白云岩发育特征,顶部为中—薄层状白云岩。灯影组/陡山沱组界线附近的这种岩石组合序列特征显示该区在陡山沱组上部沉积时期就开始进入台地边缘斜坡发育阶段,至灯影组下部沉积时期表现得更为突出,转化为高陡的台地边缘。与此对应的灯影组的三分性特点与第一种类型的灯影组相似,以石板滩段薄层灰岩发育为特点,只是下部蛤蟆井段发育垮塌角砾白云岩,中部石板滩段内部滑塌构造相对发育,上部白马沱段白云岩中的颗粒较细,以薄层泥晶白云岩为主,证明这些地区的灯影组以陆棚斜坡—盆地相沉积位置,晚期因海平面下降转化为局限盆地沉积。
第三种类型的灯影组底界以宜昌灯影峡灯影组的标准剖面或宜昌晓峰滴水岩剖面为代表(图4A),主要分布在宜昌棋盘山以东的宜昌莲沱、晓峰滴水岩等地,以灯影组底界灰白色厚层—块状砂屑细晶白云岩的出现与下伏陡山沱组顶部灰黑色中—薄层状细晶白云岩相区分。由于这些地区的陡山沱组第四段主要为中—厚层状灰岩,含硅质、磷质结核的白云岩、鲕粒白云岩,因此,陡山沱组第四段与上伏灯影组蛤蟆井段下部之间沉积相连续,岩性渐变过渡[17],为台地边缘浅滩相沉积,只是至灯影组蛤蟆井段下部时进入浅滩快速增长发育时期,地层的沉积速率增大,地层单层厚度变厚。与此对应的灯影组在岩石组合特征上三分性明显,下部蛤蟆井段和上部白马沱段白云岩的厚度与中部石板滩段薄层灰岩的厚度接近,证明这些地区的灯影组以台地边缘浅滩沉积为主,中期因海平面上升进入陆棚斜坡沉积环境。
4.2海平面变化与碳同位素地层
峡东不同古地理部位灯影组碳同位素组成变化的对比显示,峡东地区埃迪卡拉系灯影组碳同位素组成受古地理(或沉积环境)和海平面变化的多重影响。
岩石组合序列和沉积相标志分析显示,以莲沱灯影峡和晓峰滴水岩剖面为代表的灯影组底部附近地层的海平面升降变化旋回开始于陡山沱组第四段底部,在陡山沱组—灯影组界线附近达到海平面最高,之后逐步下降,至蛤蟆井段顶部达到其最低点,形成了灯影组第一个海平面升降旋回或灯影组下部层序(简称LDYS)。与之对应,陡山沱组第四段至灯影组蛤蟆井段下部的δ13C随海平面的上升而出现轻微正偏离,灯影组蛤蟆井段的δ13C总体上有随海平面下降而出现轻微的负偏离的现象。但与之对应的宜昌黄牛岩和秭归泗溪等地的陡山沱组第四段表现为黑色页岩,在黄牛岩一带所含透镜体灰岩中的δ13C具有强烈负偏离现象,其上覆灯影组底部薄层白云岩的δ13C仍然为负值,但之后δ13C随海平面下降先由负转为正,在蛤蟆井段中部达到其最大值之后逐步下降,在蛤蟆井段—石板滩段界线附近出现一个与莲沱剖面相似的碳同位素组成轻微负偏离。不同相区同期地层中碳同位素组成的上述明显差异,显示与这一时期海水中的碳同位素组成存在一个明显的梯度相关[15],区域内碳同位素组成变化的对比可能仅适用于相同环境或水深条件。
灯影组石板滩段与下伏蛤蟆井段之间明显的岩性转换面是区内灯影组中部最为明显的另一个层序不整合面。开始于石板滩段底部的灯影组第二个海平面变化旋回,结束于石板滩段中部中厚层状白云岩之顶,并由此形成灯影组第二个海平面升降旋回或灯影组中部层序(简称MDYS)(图2A)。在莲沱灯影峡、雾河和黄牛岩一带主要由灯影组石板滩段下部薄层状灰岩和中部薄层状灰岩、白云质灰岩和白云岩组成。但往西至秭归泗溪、庙河一带该海平面变化旋回的顶界面不清,证明这一次的海平面下降的幅度不大。此次海平面升降变化旋回中碳同位素组成变化趋势与灯影组下部旋回碳同位素变化趋势相似,早期伴随海平面上升而出现短暂的轻微正偏离之后,δ13C保持一段时间的稳定,直到海平面下降,白云质灰岩、白云岩的出现,才发生δ13C的轻微下降。此次海平面升降旋回中碳同位素组成在区域上的相对稳定和纵向上变化的相似性,证明峡东地区当时的古海洋环境和古地理条件区域接近,除了海侵时期生物含量的增加以及海退时期可能的陆源碎屑含量增加或生物减少或多或少地影响当时海水碳同位素的组成外,整个峡东地区当时海水中碳同位素组成特点接近。
灯影组中部第三个海平面变化旋回或上部程序(UDYS)在峡东莲沱灯影峡剖面上对应于石板滩段上部,主要由底部薄层状灰岩,中部薄层状硅质白云岩和上部厚层夹薄层白云岩组成(图2A)。往西至黄牛岩、李家院子一带相变为由下部薄层状灰岩,中部含燧石结核或条带厚层状灰岩和上部薄层状白云岩组成。该层序内部碳同位素组成变化特点与前述灯影组第二个海平面变化旋回的碳同位素组成变化特点相似,但伴随海平面上升和下降所发生的δ13C的上升和下降幅度较小,未见明显δ13C负异常。
峡东灯影峡剖面上灯影组第四个海平面变化旋回以灯影组上部白马沱段中厚层状含内碎屑白云岩、鲕粒白云岩的出现为标志,顶部以寒武系水井沱组底部明显的层序转换面为界。同期地层碳同位素组成变化以灯影组白马沱段底部和天柱山段顶部分别出现两次δ13C负偏离为特点。从天柱山段的小壳化石具有梅树村期小壳化石组合1和组合2混生的特点来看,天柱山段沉积时期区内局部地区就存在陆上暴露现象。虽然目前缺乏足够精细的生物地层资料证明黄陵东翼,如宜昌灯影峡至晓峰一带灯影组白马沱段与西翼,如宜昌雾河至秭归庙河一带岩家河组是否同时异相沉积,但从雾河计家坡一带石板滩段与白马沱段连续沉积,岩性渐变,石板滩段地层之上的第一个明显的层序不整合面出现在岩家河组底界面上,界面之下白马沱段的碳同位素组成变化趋势与灯影峡石板滩段上部灯影组第三次海平面变化旋回地层中碳同位素组成变化特点相似(图2C,2D),推测峡东西翼的岩家河组可能不仅仅与天柱山段单独对比,而应与峡东东翼灯影组白马沱段和天柱山段两者之和对比。即在岩家河组标准剖面上,岩家河组下伏白云岩段实际上仅相当于灯影峡剖面上石板滩段顶部白云岩。这一点从海平面变化旋回和碳同位素组成变化特点上也可以得到进一步印证(图2)[19-20]。
4.3化石组合序列及其对比
由于受沉积相、古生物化石化作用或生物演化阶段性特点等方面的制约,目前尚不能像显生宙那样在埃迪卡拉系的一个剖面上准确限定一个或多个古生物化石属、种的最初出现或最后消失的具体层位,进而建立以某一个生物门类化石属、种的地层延限为基础的生物地层延限带,但峡东地区灯影组的部分层段保存有十分丰富的,甚至多门类共生的古生物化石,形成具有划时代意义的生物群或生物组合。基于上述灯影组底界和内部层序划分对比标志,自下而上可以在峡东上埃迪卡拉系陡山沱组第四段至灯影组石板滩段或与之相当的层位中识别出3个生物群,或化石组合。自下而上:
组合1(或Beltanelloides组合):见于秭归庙河陡山沱组第四段中下部厚约4 m的黑色炭质页岩中,层位上与灯影组底部层序的海侵体系沉积对比,化石组合上以小型埃迪卡拉型动物化石Beltanelloides和Eoandromenda octobrachiata等的出现,相伴有有大量底栖多细胞宏体藻类,如Anomalophyton,Baculiphyca,Longifuniculum,Enteromorphites,Doushantuophyton,Konglingphyton,和一些管状炭质印模化石,如 Jiuqunaoella,Cuculus,Calyptrina,Sinospongia等为特点[21-22]。埃迪卡拉动物化石中,Eoandromenda octobrachiata是澳大利亚埃迪卡拉动物群的重要分子之一,主要出现在埃迪卡拉系最上部的Rawsley组石英砂岩下部,且与当前化石的保存方式不同。Beltanelloides最初见于东欧地台文德系列德金统下部。后在加拿大Yukon地区中东部的Wernecke山以及西北部的Mackenzie山以及东欧地台等地的末前寒武系中也广泛出现,被认为是埃迪卡拉动物化石组合2或圆盘状埃迪卡拉动物化石组合的典型代表[23-25]。因此,Beltanelloides在庙河生物群中的出现证明峡东埃迪卡拉系陡山沱组第四段应至少与南澳大利亚Wonoka组上部层序以及加拿大Mackenzie地区Sheepbed组对比(图5)。
组合II(或Yangtziramulus zhangi组合):广泛出现在宜昌雾河、黄牛岩、母猪峡和秭归庙河等地灯影组石板滩段下部,距该段底部约20 m的薄层灰岩中。层位上与灯影组中部石板滩段下部层序的海侵体系对比,化石组合上以大型圆盘状、叶状埃迪卡拉动物,相伴有大量遗迹化石出现为特点。在宜昌灯影峡和秭归泗溪与上述Yangtziramulus产出层位接近的灯影组石板滩段下部还分别发现有Paracharnia dengyingensis和Xenophyophore[5-6,26]。在宜昌雾河还发现大量典型的埃迪卡拉型动物化石Hiemalora,Pteridinium,Rangea,以及 Charniodiscus 和Wutubus annularis等[8]。上述Xenophyophore,特别是遗迹化石和典型埃迪卡拉型动物化石中Charniodiscus的发现,证明上述地区灯影组石板滩段应该与澳大利亚和加拿大等地埃迪卡拉型动物化石组合3分布的层位可以直接对比[23,27]。
组合III(或Sinotubulites组合):广泛出现在宜昌灯影峡、黄牛岩和秭归庙河灯影组石板滩段中部,层位上大致与石板滩段上部层序的海侵体系对比。主要以个体较小的Cloudinasp.出现为特征,共生生物中主要有个体相对粗壮的 Sinotubulites baimatuoensis,和个体相对较小的 Sinotubulites miaoheensis等管状动物化石以及具有枝状分叉的管状化石和表面光滑,呈分裂状态的球状化石等。化石组合面貌与陕西宁强李家沟灯影组顶部的管状动物化石群的组合面貌十分接近[28]。后者与上伏寒武系底部含梅树村期小壳化石的宽川铺组呈平行不整合接触。由于Cloudina是Namibia动物群的代表性分子,因此,它的出现和大量繁盛还指示石板滩段上部与国外同期含Cloudina地层,如Oman地区Ara组下部[29],Namibia地区Urasis组[30-32]以及美国加利福尼亚地区的Striling Quartzite组上部对比[33](图5)。
4.4年代地层意义
传统上,灯影组被简单地冠以“灯影阶”或“灯影峡阶”等年代地层单位名称,归于上震旦统上部或上震旦统[34]。后汪啸风等[35]以石板滩段底部层序界面,灯影组石板滩段中部Cloudiniids的最初出现将灯影组三分,自下而上称为“庙河阶”,“泗溪阶”和“龙灯溪阶”,并将后二者一起归入上震旦统。后Zhu et al.[11]认为以灯影组石板滩段底界层序界面所确定的上震旦统的地质延限不足埃迪卡拉系整体地质延限的10%,于是建议将埃迪卡拉系二分的界线下移至陡山沱组二段内部相当于Glaskier冰期或与之冰期形成,海平面下降相关的层序界面上,同时以石板滩段下部碳同位素最大正偏离出现的位置为标志,将石板滩段及以上埃迪卡拉系地层合并建立第5阶(Stage 5),并将其与国外含Cloudiniids的地层对比。刘鹏举等[12]采纳Zhu et al.[11]关于埃迪卡拉系二统五阶的年代地层划分方案,但与后者不同的是,刘鹏举的第四阶和第五阶底界分别与峡东灯影组底部碳同位素的最大正偏离和Cloudiniids的最初出现相对应,与峡东灯影峡灯影组蛤蟆井段和白马沱段的底界的岩性界面接近或一致。上述不同学者在不同时期针对峡东灯影组之所以提出不同的年代地层单位划分方案,主要原因在于:1)早先在峡东埃迪卡拉系中没有发现典型的埃迪卡拉型动物化石,峡东地区管状动物化石Cloudiniids的分布层位及其与典型埃迪卡拉型动物化石的叠置关系不十分清楚;2)峡东地区埃迪卡拉系灯影组标准剖面上灯影组的碳同位素地层研究不够精细,灯影组碳同位素组成变化与海平面变化的精细对比关系没有很好的建立;3)国际上寒武系底界的划分的标志及其同位素地质年代学研究进展,以及Oman地块上Cloudiniids的消失和与之相伴的碳同位素负偏离的在全球寒武系底界划分对比中的重要意义注意不够[29]。
本次调查研究结果显示,峡东地区埃迪卡拉动物和以Cloudiniids发育为特征的管状动物分别出现在石板滩段下部和上部两次不同的海侵沉积体系中,具有明显的上、下叠置关系。由于这两种生物是世界上广泛分布的埃迪卡拉型动物群和纳米比亚动物群的典型分子,因此,它们的出现和繁盛是埃迪卡拉动物和管状动物辐射的标志,完全具有洲际性地层对比的重要意义,理应分别作为年代地层单位划分对比不可或缺的重要标志。正因为如此,本文仍然坚持作者较早前提出的关于灯影组内部年代地层单位划分的观点,即灯影组中部石板滩段下部层序和石板滩段上部层序对应于两个不同的年代地层单位——泗溪阶和龙灯溪阶的观点。由于石板滩段上部Cloudiniids消失之后,石板滩段顶部中薄层状泥晶白云岩夹(互)块状泥晶白云岩之上厚层块状夹中层状粉晶白云岩之底约10 m厚的地层存在明显的碳同位素负偏离,δ13C在较短地层中从+2‰迅速下降至-10‰,甚至更低,然后又恢复到+2‰之后逐步稳定,显示出与Oman地区埃迪卡拉系—寒武系界线附近生物和碳同位素组成变化关系的明显相似性,暗示峡东地区寒武系的底界应在灯影峡灯影组标准剖面上白马沱段与石板滩段界线附近,在那里大致与厚层块状粉晶白云岩出现所形成的海侵沉积的底界接近,即早先所建立的龙灯溪阶上部的绝大部分地层应该是寒武系,而非埃迪卡拉系。
由于埃迪卡拉型动物化石和Cloudiniids的生物分类位置和起源关系不明,目前尚不具备研究这些生物类型中某个属、种的演化关系,因此,尽管我们可以像显生宙那样粗略地用这些生物的繁盛或大量出现作为区域地层划分对比的依据,但目前尚不具备利用这些门类中的某个化石属种像显生宙那样确定年代地层单位的界线点位,因此,这里借助与上述生物群繁盛相配套的海平面变化旋回和碳同位素地层划分对比结果,建议以灯影峡灯影组石板滩段下部和上部海平面上升时期所对应的碳同位素最大正偏离分别作为对之前所建立的泗溪阶和龙灯溪界的底界划分的新标志。重新厘定的上述两个阶的底界标志较最初以层序界面为标志所确定的底界面略高,但更符合国际地层指南关于年代地层单位底界应选择在单相连续的剖面上的要求。
重新厘定的泗溪阶和龙灯溪阶的两者的地质延限总和虽然不足埃迪卡拉系地质延限的十分之一,但这段地层无论在生物演化阶段上,还是古气候和古环境变化方面均搭架了埃迪卡拉系与寒武系的桥梁,具有地质里程碑的意义,因此,把它们与之前气候冷暖交替所形成的埃迪卡拉系中、下部地层分开,建立一个统级年代地层是合适的。据此,并结合Glasikier冰期的出现和与之相关的碳同位素强烈负偏离是埃迪卡拉系内部另外一个可能的统级年代地层单位划分的界线标志[11,12,17],本文建议将埃迪卡拉系进行三分,自下而上分别称为下、中和上埃迪卡拉统。其中前两者的底界分别位于Mari noan和Glasikier冰期结束之后,海平面快速上升之前地层中发生δ13C强烈负偏离的位置。上埃迪卡拉统的底界则与埃迪卡拉动物辐射发展时期δ13C最大正偏离位置接近(图5)。

图5 厘定的峡东地区埃迪卡拉系年代地层系统及其划分与对比(陡山沱组碳同位素组成曲线来自文献[17],Namibia,Oman,S. Australia以及NWCanada埃迪卡拉系地层、古生物和碳同位素组成曲线分别来自文献[31],[29,36],[37]和[23-25].图例见图2)Fig.5 proposed choronostratigraphic system in eastern Yangtze Gorges and its correlation(δ13C profiles of the Doushantuo Formation modified from[17],The lithological subdivision,fossils assemblages and δ13C profiles ofthe Ediacaran in Namibia,Oman,S.Australia and NWCanada are separatelyafter fromreference[31],[29,36],[37]and[23-25].Legends for the logsee Fig.2)
5 结论
(1)峡东埃迪卡拉系灯影组底界附近地层存在明显的沉积相分异,大致可以划分与盆地相、台地边缘高陡斜坡相、台地边缘浅滩相三种类型,对应有三种不同的岩石组合序列和底界划分标志。
(2)峡东陡山沱组第四段—灯影组石板滩段可以划分为三个海侵—海退旋回,分别形成上埃迪卡拉系下、中和上三个层序。上埃迪卡拉系灯影组碳同位素组成并非以前所认为的那样稳定,而是随海平面变化而变化,以至随海平面上升在灯影组蛤蟆井段底部、石板滩段底部和中部出现了δ13C的轻微正偏离,随海平面的下降在蛤蟆井段顶部、石板滩段顶部和白马沱段底部出现了δ13C的负偏离。其中以白马沱段底部4 m厚地层中δ13C的负异常最为明显,最小δ13C达到-12‰。
(3)峡东陡山沱组第四段—灯影组石板滩段或与之相当地层自下而上可以识别出Beltanelloides,Yangtziramulus zhangi和Sinotubulites等三个分别代表圆盘状、叶状和管状动物辐射发展的化石组合。由于前两者分别与加拿大埃迪卡拉动物化石组合2和组合3对比,Sinotubulites组合中含有埃迪卡拉系最晚期地层管状动物化石的典型分子Cloudina。因此,埃迪卡拉系灯影组的古生物化石组合不仅具备进行远距离对比的重要意义,而且代表埃迪卡拉动物演化发展的三个不同阶段,具有重要年代地层单位划分意义,可以作为早先在三峡地区建立的埃迪卡拉系庙河阶、泗溪阶和龙灯溪阶远距离对比的生物标志。
(4)由于灯影组石板滩段顶部Cloudiniids消失之后,天柱山段小壳化石出现之前发生了δ13C的下降,并在灯影组白马沱段下部海侵早期达到δ13C的最小值,因此,从其与Oman地区等同期地层生物、碳同位素地层和同位素地质年代学对比角度上出发,本文认为峡东地区寒武系底界应大致与灯影峡剖面上灯影组石板滩段和白马沱段的分界线对比。重新厘定的埃迪卡拉系内部可以进一步以Glasikier冰期或冰期之上碳同位素的强烈负偏离的出现,以及埃迪卡拉动物的复杂化或叶状埃迪卡拉动物的辐射为标志进一步三分为埃迪卡拉系下、中和上埃迪卡拉统。其中上埃迪卡拉统的底界与泗溪阶的底界位置一致,与叶状埃迪卡拉动物辐射发展初期δ13C最大正偏离位置一致。
[1]Lee J,Chao Y.Geology of the Gorge District of the Yangtze (from Ichang to Tzekuei),with special reference to the development ofthe Gorges[J].Bulletin ofthe Geological Society of China,1924,3(3-4):351-391.
[2]Wang X F,Erdtemann B D,Chen X H,Mao X D.Integrated sequence-,bio-,and chemostratigraphy ofthe Terminal Proterozoic to Lowermost Cambrian“black rock series”from central south China[J].Episodes,1998,21(3):178-189.
[3]Jiang G Q,Shi X Y,Zhang S H,Wang Y,Xiao S H.Stratigraphy and paleogeography of the Ediacaran Doushantuo Formation(ca.635-551 Ma)in South China[J].Gondwana Research,2011,19(4):831-849.
[4]陈孝红,李华芹,陈立德,王传尚,汪啸风.三峡地区震旦系碳酸盐地层碳、氧同位素特征 [J].地质论评,2003,49(1):66-73.
[5]Weber B,Steiner M,Zhu M Y.Precambrian-Cambrian trace fossils fromthe Yangtze Platform(South China)and the early evolution of bilaterian lifestyles[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,254(s1-2):328-349.
[6]丁启秀,陈忆元.湖北峡东地区震旦纪软躯体后生动物化石的发现及其意义[J].地球科学,1981,(2):53-57.
[7]Shen B,Xiao S H,Zhou C M,Yuan X L.Yangtziramulus zhanginewgenus and species,a carbonate-hosted macrofossil from the Ediacaran Dengying Formation in the Yangtze Gorges area,South China[J].Journal of Paleontology,2009,83 (4):575-587.
[8]ChenZ,ZhouCM,XiaoSH,WangW,GuanCG,HuaH,YuanX L.NewEdiacara fossils preserved in marine limestone and their ecologicalimplications[J].ScientificReports,2014,4(8):4180.
[9]陈孟莪,王义昭.峡东区上震旦统灯影组中段的管状动物化石[J].科学通报,1977,(4-5):219-222.
[10]丁启秀,邢裕盛,王自强,尹崇玉,高林志.湖北庙河—莲沱地区灯影组管状化石及遗迹化石[J].地质论评,1993,39 (2):119-123.
[11]Zhu MY,Zhang J M,Yang A H.Integrated Ediacaran(Sinian)chronostratigraphy of South China[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,254(1-2):7-61.
[12]刘鹏举,尹崇玉,陈寿铭,李猛,高林志,唐烽.华南峡东地区埃迪卡拉(震旦)纪年代地层划分初探[J].地质学报,2012,86(6):849-866.
[13]Lambert I B,Walter MR,ZangWL,Lu S N,Ma G G.Pale-oenvironment and carbon isotope stratigraphy of Upper Proterozoic carbonates of the Yangtze platform[J].Nature,1987,325:140-142.
[14]王自强,尹崇玉,高林志,柳永清.湖北宜昌峡东地区震旦系层型剖面化学地层特征及其国际对比 [J].地质论评,2002,48(4):408-415.
[15]Jiang G Q,Kaufman A J,Christie-Blick N,Zhang S,Wu H.Carbon isotope variability across the Ediacaran Yangtze platform in South China:Implications for a large surface-to-deep ocean δ13C gradient[J].Earth and Planetary Science Letters,2007,261(1-2):303-320.
[16]王自强,高林志,尹崇玉.2001,峡东地区震旦系层型剖面的界定与层序划分[J].地质论评,2001,47(5):449-458.
[17]陈孝红,周鹏,张保民,王传尚.峡东埃迪卡拉系陡山沱组稳定碳同位素记录及其年代地层意义 [J].中国地质,2015,42(1):207-223.
[18]Zhu M Y,Lu M,Zhang J M,Zhao F C,Li G X,Yang A H,Zhao X,Zhao M J.Carbon isotope chemostratigraphy and sedimentary facies evolution of the Ediacaran Doushantuo Formation in western Hubei,South China[J].Precambrian Research,2013,225(1):7-28.
[19]吕苗,朱茂炎,赵美娟.湖北宜昌茅坪泗溪剖面埃迪卡拉系岩石地层和碳同位素地层研究 [J].地层学杂志,2009,33(4):359-372.
[20]Ishikawa T,Ueno Y,Komiya T,Sawaki Y,Han J,Shu D G,Li H,Maruyama S,Yoshida N.Carbon isotope chemostratigraphy of a Precambrian/Cambrian boundary section in the Three Gorge area,South China:Prominent global-scale isotope excursions just before the Cambrian Explosion[J].Gondwana Research,2008,14(1-2):193-208.
[21]陈孟莪,萧宗正.峡东上震旦统陡山沱组宏体化石生物群[J].古生物学报,1992,31(5):513-520.
[22]尹崇玉,唐烽,刘鹏举,高林志,王自强,陈寿铭.华南埃迪卡拉(震旦系)陡山沱组生物地层学研究的新进展[J].地球科学,2009,30(4):421-432.
[23]Narbonne G M,Hofmann H J.Ediacaran biota of the Wernecke Mountains,Yukon,Canada[J].Palaeontology,1987,30 (4):647-676.
[24]Narbonne G M,Kaufman A J,Knoll A H.Integrated chemostratigraphy and biostratigraphy of the Windermere Supergroup,northwestern Canada:Implication for Neoproterozoic correlations and the early evolution of animals[J]. Geological Society of America Bulletin,1994,106(10):1281-1292.
[25]Pyle L J,Narbonne G M,James N P,Dalrymple R W,Kaufman A K.Integrated Ediacaran chronostratigraphy,Wernecke Mountains,northwestern Canada[J].Precambrian Research,2004,132(1-2):1-27.
[26]Sun W G,Late Precambrian Pennatulids(sea pens)from the eastern Yangtze Gorges,China:Paracharnia gen.nov[J].Precambrian research,1986,31(4):361-375.
[27]Seilacher A S,Grazhdankin D,Legouta A.Ediacaran biota:The dawn of animal life in the shadow of giant protists[J]. Paleontological Research,2003,7(1):43-54.
[28]华洪,张录易,张子福,王静平.高家山生物群化石组合面貌及其特征[J].地层学杂志,2001,25(1):13-17.
[29]Amthor J E,Grotzinger J P,SchrōDER S,Bowring S A,Ramezani J,Martin M W,Matter A.Extinction of Cloudina and Namacalathus at the Precambrian-Cambrian boundary in Oman[J].Geology,2003,31(5):431-434.
[30]Grotzinger J P,Bowring S A,Saylor B Z,Kaufman A J.Biostratigraphic and geochronologic constraints on early animal evolution[J].Science,1995,270:598-604.
[31]Saylor B Z,Kaufman A J,Gortzinger J P.Urban F.A composite reference section for terminal Proterozoic strata of Southern Namibia[J].Journal ofSedimentaryResearch,1998,68(6):1223-1235.
[32]Halverson G P,Hoffman P F,Schrag D P,Maloof A C,Rice A H N.Toward a Neoproterozoic composite carbon-isotope record[J].Geological Society of America Bulletin,2005,117,(9):1181-1207.
[33]Corsetti F A,Kaufman A J.Stratigraphic investigations of carbon isotope anomalies and Neoproterozoic ice ages in Death Valley,California[J].Geological Society of America Bulletin,2003,115(8):916-932.
[34]邢裕盛,尹崇玉,高林志.震旦系的范畴、时限及内部划分[J].现代地质——中国地质大学研究生院学报,1999,(2):202-204.
[35]汪啸风,陈孝红,王传尚,陈立德.震旦系底界及内部年代地层单位划分 [J]. 地层学杂志,2001,25(增刊):370-376.
[36]Le Guerrou′e E,Allen P A,Cozzi A.Chemostratigraphic and sedimentological framework of the largest negative carbon isotopic excursion in Earth history:The Neoproterozoic Shuram Formation(Nafun Group,Oman)[J].Precambrian Research,2006,146(s1-2):68-92.
[37]Calver C R.Isotope stratigraphy of the Ediacarian(Neoproterozoic III)of the Adelaide Rift Complex,Australia,and the overprint ofwater column stratification[J].Precambrian Research,2000,100(1-3):121-150.
CHEN Xiao-Hong,ZHOU Peng,ZHANG Bao-Min,WEI Kai,ZHANG Miao
(Wuhan Center of China Geological Survey,Wuhan 430205,Hubei,China)
Lithostratigraphy,Biostratigraphy,Sequencestratigraphy and Carbon Isotope Chemostratigraphy of the Upper Ediacarian in Yangtze Gorges and Their Significance for Choronostratigraphy.
Litho-,bio-,sequence-and carbon isotope chemo-stratigraphy of the Upper Ediacaran ranging from upper part of the Doushantuo Formation to the Shibantan member of the Dengying Formation in eastern Yangtze Gorges were studied in this paper.Conclusions show that there are different kinds of sedimentary facies,which included such as platform,slope with high steep margin and basin facies,in the upper part of the Doushantuo Formation and lower part of the Dengying Formation in the eastern Yangtze Gorges.The subdivision of the basal boundary of the Dengying Formation and the members of the Dengying Formation are obviously restricted by sedimentary facies.The base boundary marked by occurrence of dolostone for the Dengying Formation is not an isochronal boundary.Correlated with two transgression-regression cycles,twice increasing-decreasing trend of δ13C value and twice important biodiversity events marked by occurrence ofvendobiontan and tublar fossils were happened separately in the lower and the upper part of the Shibantan member of the Dengying Formation in Dengying Gorges section.Consequence,two regional chronostratigraphic units(Sixi and Longdengxi stages)would be recognized in the Shibantan member of the Dengying Formtion in the eastern Yangtze Gorges.The decreasing of δ13C value between the disappearance of the Cloudiniids fossils at the upper part of the Shibantan member and the appearance of the small shelly fossils at the upper part of the Baimatuo member led a very negative δ13C value of-12‰at the base of the Baimatuo member of the Dengying Formation.The basal boundary of the Cambrian in the eastern Yangtze Gorges should be correlated with the occurrence of this negative interval,i.e the boundary between the Shibantan and Baimatuo members based on the changes of the fossil assemblages and δ13C and their correlations with other contemporaneity strata in the world.
Dengying Formation;stable carbon isotope;Ediacaran fauna;tubular fauna;Ediacaran-Cambrian boundary
中图分类法:P534.31A
1007-3701(2016)02-087-19
10.3969/j.issn.1007-3701.2016.02.001
2016-03-24;
2016-04-13.
中国地质调查局项目“华南中部地壳演化及其资源制约”(1212011121103)资助.
陈孝红(1964—),男,研究员,主要从事地层和古生物研究,E-mail:yccxiaohong@163.com.
猜你喜欢
广东第二课堂·小学(2022年5期)2022-06-02
知识窗(2021年6期)2021-10-07
趣味(语文)(2018年11期)2019-01-14
——贵州省遵义市播州区石板镇见闻
西部大开发(2018年5期)2018-07-03
小学生作文(低年级适用)(2017年5期)2017-07-07
天然气勘探与开发(2015年3期)2015-12-08
小学生导刊(中年级)(2014年7期)2014-08-04
故事会(2009年5期)2009-05-21
山西文学(2008年12期)2008-12-24
