
不同发育阶段东、黄海凤鲚的食性差异
2016-08-31 07:21朱文斌王忠明蒋日进周永东
浙江海洋大学学报(自然科学版) 2016年1期
郭 爱,陈 峰,朱文斌,王忠明,蒋日进,周永东
(1.浙江省海洋水产研究所,农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021;2.上海海洋大学海洋科学学院,上海 201306;3.国家海洋局第二海洋研究所,浙江杭州 310012)
不同发育阶段东、黄海凤鲚的食性差异
郭爱1,2,3,陈峰1,朱文斌1,王忠明1,蒋日进1,周永东1
(1.浙江省海洋水产研究所,农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山316021;2.上海海洋大学海洋科学学院,上海201306;3.国家海洋局第二海洋研究所,浙江杭州310012)
摘要:根据东海区(26°-35°00′N,120°00′-126°30′E)大面调查渔获物的凤鲚样本,对东、黄海凤鲚不同发育阶段的食物组成、摄食强度、食物生态位宽度和食物团单个饵料的重量差异进行了研究,并对不同体长组的凤鲚进行了聚类分析。研究结果表明:(1)凤鲚不同阶段的食物组成随着发育的变化有所不同,根据%IRI分析,桡足类为各个发育阶段的最重要的食物,糠虾类、磷虾类为次重要的类群,随着体长的增加小型鱼类在食物组成的%IRI比重越来越高。(2)凤鲚有两个摄食强度的高峰:幼体组70~100 mm和成体组191~250 mm,平均胃饱满系数均超过10‰。空胃率和体长呈现出负相关关系,随着体长的增加,空胃率不断下降。(3)食物团中单个饵料的平均重量随着凤鲚体长的增加而增加,两者呈现正相关关系。体长70~160 mm阶段,单个饵料的平均重量都小于10 mg,体长161~250 mm阶段,单个饵料的平均重量均大于30 mg。(4)聚类分析表明可将的凤鲚6个体长组可以分为3组:70~100 mm、101~130 mm、131~250 mm。体长70~100 mm的凤鲚主要以桡足类和幼体类为食,体长101~130 mm的凤鲚主要摄食桡足类、糠虾类和磷虾类,体长131~250 mm的凤鲚主要摄食桡足类、糠虾类和鱼类。(5)凤鲚不同发育阶段食物生态位宽度也存在差异,体长101~130 mm饵料多样性指数最高为2.37,体长70~100 mm饵料多样性指数最低为0.96,体长131~250 mm之间各体长组饵料多样性指数相差不大。
关键词:东、黄海;凤鲚;发育阶段;食性;%IRI
凤鲚Coilia mystus为沿海中小型鱼类,栖息于港湾、河口附近,常为沿海定置张网所渔获[1]。主要分布于印度洋北部沿海和太平洋西部沿海,东至中国、朝鲜、日本,南至印度尼西亚,中国沿海均有分布,在较大的江河、湖泊均有出产,如珠江、闽江、长江、太湖等,尤以长江中下游最多[2-3]。
鱼类食性研究是阐述海洋生态系统结构和功能的基础,作为渔业研究的重要命题广为国内外所重视[4]。凤鲚为我国湾口和沿岸的重要经济鱼类,关于凤鲚生物学和资源利用有过持续和详细的研究,对凤鲚摄食习性也有过一些研究,主要是对河口和湾口等区域性的样品进行过一些研究和分析。上世纪50年代郑重等[5]对厦门港凤鲚摄食和影响凤鲚摄食的因素进行过研究,结果表明,凤鲚饵料以甲壳动物为主,其中以桡足类最为重要,糠虾类和十足类次之。刘守海等[6-7]对长江口和杭州湾6-8月凤鲚的摄食情况及食物组成与研究区域的海洋浮游动物进行过比较研究,分析表明胃含物和海洋中全部种类的相似值大致在0.363~0.365之间,两者的桡足类的相似值在0.521~0.575之间。此外,关于凤鲚的生物学研究中也提及过凤鲚主要的摄食种类[8-9],但是没有详细的定量分析。目前尚无人对不同发育阶段凤鲚的摄食差异进行过系统的研究。本研究根据2008年5月-2009年2月东海区资源大面调查的渔获物所取得的样本对东、黄海凤鲚不同发育阶段的食性差异进行了分析,包括对不同发育阶段食物组成、摄食强度、食物生态位宽度和食物团单个饵料的重量差异进行了研究,并对不同体长组的凤鲚进行了聚类分析,旨在为全面了解东、黄海凤鲚的摄食生态及摄食行为奠定基础,为凤鲚渔业资源的合理利用和可持续发展提供基础资料。
1材料与方法
1.1样本来源
样本来自2008年5月~2009年2月东海区资源大面调查的渔获物,取样时间分别为春季(3-5月)、夏季(6-8月)、秋季(9-11月)、冬季(12月-次年2月),春、夏季样本的范围为28°-35°00′N,124°E以西海域,秋、冬季样本的范围为27°30′-35°00′N,125°30′E以西海域。总计分析的样品数量为416 ind,其中春季159 ind,夏季91 ind,秋季76 ind,冬季90 ind,样品体长范围为77~241 mm,体重范围为1.6~58.2 g。
1.2样本处理
样品经生物学测定后,取出其胃用10%的福尔马林液固定保存。摄食强度根据海洋调查规范海洋生物调查部分鱼类摄食强度分级标准[10]分为0~4级,消化等级分为1~5级。胃含物分析在Nikon SMZ-2000解剖镜下进行,重量测定采用精度0.001 g的电子天平。分析时先用吸水纸吸去胃表面的水分,测定胃的总重,并记录摄食等级,然后解剖分析食物团,记录消化等级,鉴别饵料生物种类并分别计数,对于可以称重的种类进行称重并记录,被消化的甲壳类根据残留的头胸甲、附肢、尾扇等不同特征进行鉴别,消化程度较高的鱼类则通过比较难消化的器官(例如:耳石、眼珠等)进行鉴定,部分胃含物由于已消化或只剩残骸,采用更正重量计算,种类鉴定时尽可能鉴定到最小的分类单元。
1.3数据处理
用于评价饵料重要性的指标[11]包括出现频率(F%)、重量百分比(W%)、个数百分比(N%)、相对性重要指数(IRI)和相对性重要指数百分比(%IRI),计算公式如下:
(IRI)=(W%+N%)F%
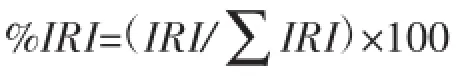
摄食强度[12]用空胃率和胃饱满系数来表示,计算公式如下:

食物多样性指数[13]用shannon-weaver指数H′来表示,计算公式如下:
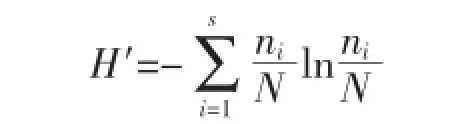
式中,ni为饵料i的个数,N为饵料总个数。
本研究应用SPSS22.0软件进行聚类分析,分析不同体长组凤鲚食物组成的相似性,选择的指标为IRI,分析过程中将数据进行标准化转换到0~1,采用的指标间距离计算方法为欧氏距离(Euclidean distance)[14]。
2结果
2.1凤鲚不同发育阶段的饵料组成
根据样本的体长范围,将样本以30 mm为组距分为6个体长组:70~100、101~130、131~160、161~190、191~220、221~250 mm,各体长组的样本数量分别为41、137、151、62、14、11尾。凤鲚不同发育阶段食物组成%IRI见表1,由表1可得,凤鲚摄食的饵料生物共计有40种(不包括6种不可辨认类型饵料),分属10个类群,其中桡足类种类最多有17种,其次为糠虾类有7种。
凤鲚不同阶段的食物组成随着发育的变化有所不同,根据%IRI分析,桡足类为各个发育阶段的最重要的食物,糠虾类、磷虾类为次重要的类群,随着体长的增加小型鱼类在食物组成的%IRI比重越来越高。体长70~100 mm的凤鲚主要以桡足类和幼体类为食,%IRI占94.36%,摄食的种类有11种,其中代表性的种类为小拟哲水蚤Nannocalanus minor(%IRI为83.62%)和六肢幼体Balanus nauplius(%IRI为7.67%)。体长101~130 mm的凤鲚主要摄食的种类为桡足类、糠虾类和磷虾类,%IRI占99.6%,摄食的种类有28种,代表性的种类为小拟哲水蚤(%IRI为42.55%)、亚强真哲水蚤Eucalanus subcrassus(%IRI为16.34%)、宽尾刺糠虾Acanthomysis latiscauda(%IRI为5.19%)和太平洋磷虾Euphausia pacifica(%IRI为8.07%)。体长131~160 mm的凤鲚主要摄食的种类为桡足类和糠虾类,%IRI占95%,摄食的种类有30种,代表性的种类为亚强真哲水蚤(%IRI为65.19%)、短额刺糠虾Acanthomysis brevirostris(%IRI为10.39%)和宽尾刺糠虾(%IRI为9.32%)。体长161~190 mm的凤鲚主要摄食的种类为桡足类和鱼类,%IRI占91.2%,摄食的种类有23种,代表性的种类为亚强真哲水蚤(%IRI为76.38%)和矛尾虾虎鱼Chaeturichths stigntatias(%IRI为7.04%)。体长191~220 mm的凤鲚主要摄食的种类为桡足类、糠虾类、十足类和鱼类,%IRI占96.1%,摄食的种类有15种,代表性的种类为亚强真哲水蚤(%IRI为33.93%)、宽尾刺糠虾(%IRI为22.58%)、细螯虾Leptochela gracilis(%IRI为18.31%)和七星底灯鱼Benthosema pterotum(%IRI为12.08%)。体长221~250 mm的凤鲚主要摄食的种类为桡足类、鱼类和十足类,%IRI占90.8%,摄食的种类有11种,代表性的种类为亚强真哲水蚤(%IRI为58.72%)、矛尾虾虎鱼(%IRI为10.79%)、七星底灯鱼(% IRI为10.79%)和细螯虾(%IRI为7.84%)。
2.2凤鲚不同发育阶段摄食强度的变化
凤鲚摄食强度随发育的变化明显(图1),幼体组70~100 mm阶段和成体组191~250 mm阶段平均胃饱满系数较高,均超过10‰,体长101~190 mm阶段胃饱满系数相对较低,均在7‰以下。空胃率和体长呈现出负相关关系,随着体长的增加,空胃率不断下降。
2.3凤鲚不同发育阶段单个饵料的质量变化
随着凤鲚体长的增加,食物团中单个饵料的平均重量也在增加,两者呈现正相关关系(图2)。体长70~160 mm阶段,单个饵料的平均重量都小于10 mg,体长161~250 mm阶段,单个饵料的平均重量均大于30 mg,体长131~190 mm阶段为单个饵料平均重量变化最显著阶段,由表1分析可得体长131~190 mm阶段是凤鲚开始摄食小型鱼类的关键阶段,体长131~160 mm阶段鱼类%IRI仅为0.9%,体长161~190 mm阶段鱼类%IRI达到12.02%,因此认为摄食鱼类是导致体长131~190 mm阶段单个饵料重量变化显著的重要原因。
2.4凤鲚不同发育阶段的食物组成聚类分析
聚类分析表明(图3),体长70~250 mm的凤鲚6个体长组可以分为3组,其中70~100 mm为一组,101~130 mm为一组,131~250 mm为一组。体长70~100 mm的凤鲚主要以桡足类和幼体类为食,体长101~130 mm的凤鲚主要摄食的种类为桡足类、糠虾类和磷虾类,体长131~250 mm的凤鲚主要摄食桡足类、糠虾类和鱼类。此外,聚类分析表明131~250 mm的凤鲚也可分为三组:131~160 mm、161~190 mm、191~250 mm,131~160 mm主要摄食桡足类和糠虾类,161~190 mm主要摄食桡足类和鱼类,191~250 mm主要摄食的种类为桡足类、鱼类和十足类。
2.5凤鲚不同发育阶段食物的生态位宽度差异
东、黄海凤鲚不同发育阶段食物生态位宽度也存在差异(表2),Shannon-Weiner多样性指数H′计算结果显示,凤鲚体长101~130 mm饵料多样性指数最高为2.37,体长70~100 mm饵料多样性指数最低为0.96,体长131~250 mm之间各体长组饵料多样性指数相差较小,表明凤鲚幼体阶段70~100 mm摄食的食谱宽度较小,体长101~130 mm阶段摄食的食谱宽度增加到最大,之后体长131~250 mm阶段摄食的食谱宽度相对较稳定。从饵料的种类来分析,幼体组70~100 mm和成体组191~250 mm摄食的饵料种类相对较少,体长101~190 mm饵料生物种类较丰富。
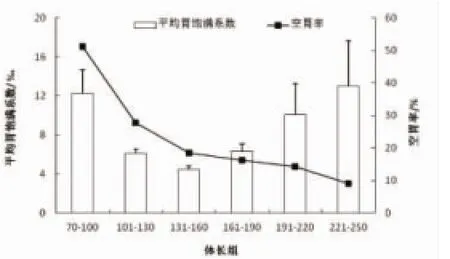
图1 凤鲚不同体长组的平均胃饱满系数和空胃率Fig.1 Mean stomach fullness index and percentage of empty stomachs of C.mystus in different size class
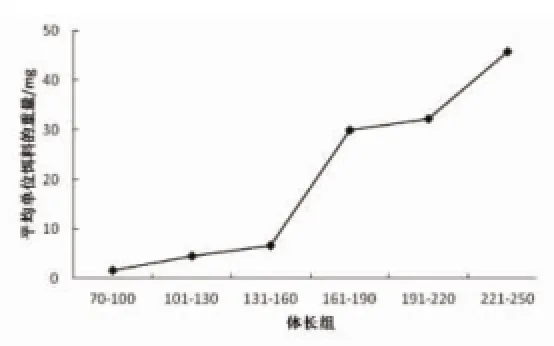
图2 凤鲚各体长组胃中平均单个饵料的质量Fig.2 Mean weight of single prey in the stomach of C.mystus in different size class
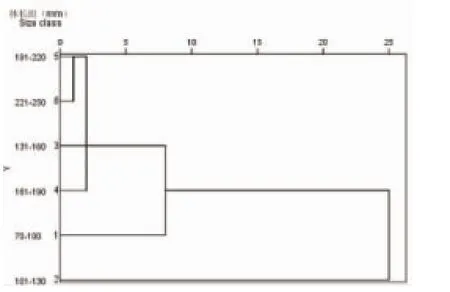
图3 凤鲚各体长组食物组成聚类分析图Fig.3 Dendrogram of cluster analysis for dietary composition of C. mystus in each size class
3讨论
3.1东、黄海凤鲚不同发育阶段摄食类群和种类的转换
摄食的饵料生物发生转换在鱼类生活史中较为普遍,蒋日进等[15]对竹筴鱼Trachurus japonicas、颜云榕等[16]对北部湾带鱼Trichiurus lepturus、张波等[17]对黄海中部高眼鲽Clesithenes herzensteini和薛莹等[18]对黄海中部小黄鱼Pseudosciaena polyactis Bleeker研究表明这些鱼类均出现过明显的食性转换现象。凤鲚
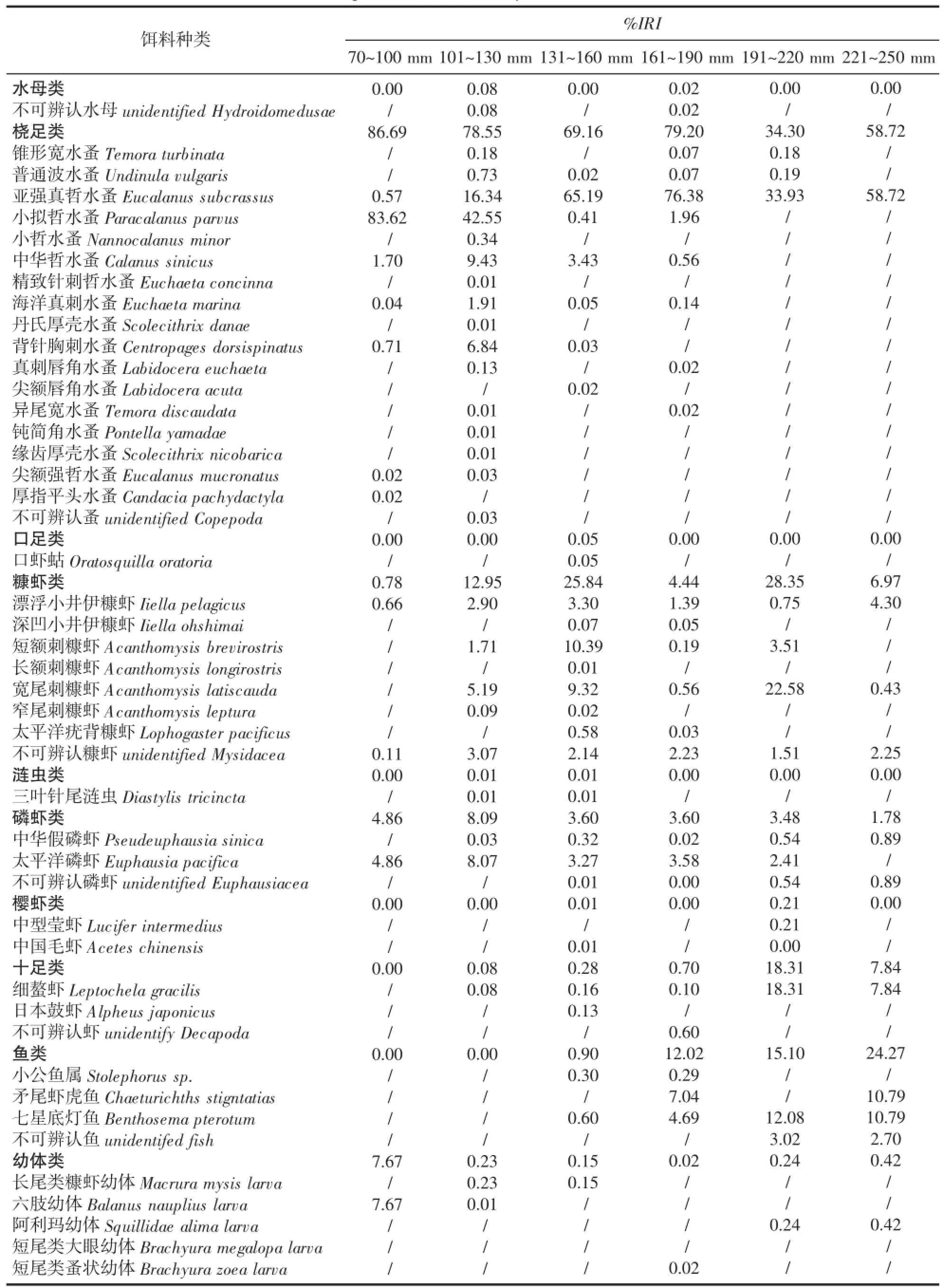
表1 凤鲚不同体长组食物组成Tab.1 Food composition of Coilia mystus in different size class
整个发育过程中,主要饵料的类群同样出现了明显的转换,部分主要类群的饵料种类也出现明显的变化(表1)。桡足类饵料在体长70~190 mm阶段%IRI比重明显高于191~250 mm阶段,70~190 mm阶段%IRI超过69%,191~250 mm阶段%IRI低于60%。糠虾类饵料在体长70~100 mm阶段出现较少,之后明显增多。而十足类饵料则在体长70~190 mm阶段出现较少,之后明显增多。鱼类饵料在体长70~130 mm阶段没有出现,之后随着个体的发育,鱼类在饵料中重要性程度越来越高。幼体类饵料仅在体长70~100 mm阶段占有重要作用,之后幼体类饵料在整个食物中的比例极低。桡足类为整个生命周期中最重要的饵料,但不同的阶段优势种类出现转换,体长70~130 mm阶段小拟哲水蚤占明显优势,体长131~250 mm亚强真哲水蚤占明显优势,体长101~160 mm阶段为两种桡足类饵料优势转换的重要阶段,体长较小阶段小拟哲水蚤为主,体长较大阶段以亚强真哲水蚤为主。其它饵料类群中未出现明显的种类随发育而转换的现象。
3.2东、黄海凤鲚不同发育阶段摄食强度的变化
鱼类的摄食强度随发育变化的规律因鱼而异,MORATO等[19]对黑尾鱼旨Serranus atricauda和HOVDE等[20]对大比目鱼Reinhardtius hippoglossoides的研究发现,其摄食强度随体长的增加而增加;LUKOSCHEK等[21]研究认为,条斑副绯鲤Parupeneus barberinusd的摄食强度随体长的增加而减弱;张波等[22]对带鱼和贺舟挺等[23]对黄鮟鱇Lophius litulon研究表明,其摄食强度随体长的变化不显著。根据胃饱满系数分析(图1),凤鲚不同发育阶段的有两个摄食强度的高峰:70~100 mm,191~250 mm。在发育早期70~100 mm阶段,为了幼体的快速成长,摄食强度相对较大,到了体长191~250 mm的繁殖盛期阶段,为了满足性腺的发育,摄食强度也较大,这和研究发现的蓝圆鲹食性有相似的规律[24]。从不同发育阶段的空胃率情况来分析,空胃率呈现出随体长增加而下降的趋势,说明个体的大小直接影响凤鲚能否捕食到食物的概率,个体较大的凤鲚更容易获得食物,这与凤鲚的摄食习性相关。凤鲚为中小型鱼类,为很多大中型鱼类的主要捕食对象,作为捕食者主要以摄食浮游动物为主,非凶猛的捕食类型,个体大小是决定捕食能力的关键因素之一。
3.3东、黄海凤鲚不同发育阶段饵料的选择
鱼类的口裂和鳃耙间距是饵料大小选择的主要因子[4],口裂大小影响摄食鱼类饵料个体的大小,鳃耙间距大小则影响摄食浮游生物饵料个体的大小,随着鱼类的发育,口裂和鳃耙间距也逐渐变大,可摄食的饵料的大小会随着体长的增加而变大。凤鲚食物团单个饵料的种类随体长增加而增大,符合鱼类发育的摄食规律。
凤鲚体长131~190 mm阶段单个饵料重量变化显著,完成了从体长131~160 mm阶段单个饵料平均重量小于10 mg到体长161~190 mm阶段单个饵料平均重量均大于30 mg的转变,如果仅仅是桡足类种类发生变化,不足以使单个饵料平均重量发生如此巨大的改变,应该是摄食了其他重量更大的饵料,分析该阶段的饵料组成发现此阶段恰好为凤鲚开始摄食小型鱼类的关键阶段,体长131~160 mm阶段鱼类% IRI仅为0.9%,体长161~190 mm阶段鱼类%IRI达到12.02%,表明摄食鱼类是体长131~190 mm阶段单个饵料重量变化显著的重要原因。从鱼类发育生理学来分析,对于一定长度的捕食鱼类来说,其选择的饵料对象的宽度有一个临界的上限和下限,当饵料的宽度超过上限和下限时,摄食的概率会迅速下降。体长131~160 mm阶段是凤鲚口裂发育到可以摄食小型鱼类的阶段,也是凤鲚的口裂发育到开始可以摄食小型鱼类的上限,但是摄食的概率较低,随着发育的推进,凤鲚摄食饵料的宽度范围也随着增大,体长161~190 mm阶段,口裂发育到适合摄食小型鱼类的合适大小,摄食小型鱼类的概率也随之加大,食物中出现鱼类的比例也随之提高,从而出现鱼类在该阶段%IRI迅速上升,进而导致单个饵料的平均重量上升明显。

表2 不同发育阶段凤鲚食物多样性指数的变化Tab.2 Food diversity index of C.mystus in different development stages
凤鲚整个生命周期以桡足类为主要摄食对象,但随着发育的变化桡足类的种类和个体大小发生了变化,体长70~130 mm阶段个体较小的小拟哲水蚤占明显优势,体长131~250 mm个体较大的亚强真哲水蚤占明显优势,这和鳃耙间距大小有密切的关系,体长较大的凤鲚鳃耙间距较大,因此能够摄食个体较大桡足类,且随着个体的发育,凤鲚倾向于摄食个体较大的饵料生物,这也符合“最佳摄食理论”[25]。
参考文献:
[1]周永东,薛利建,徐开达.舟山近海凤鲚的生物学特征研究[J].现代渔业信息,2004,19(8):19-21.
[2]张国祥,倪勇.鲱形目[M]//中国水产科学研究院东海水产研究所,上海市水产研究所.上海鱼类志.上海:上海科技出版社,1990:93-115.
[3]丘书院.鲱形目:鳀科[M]//朱元鼎.福建鱼类志.福州:福建科技出版社,1984:140-158.
[4]殷名称.鱼类生态学[M].北京:中国农业出版社,1995:1-293.
[5]郑重,方金钏.厦门鲚鱼的食料研究——1.六丝鲚(Coilia mystus)的食料分析[J].厦门大学学报:自然科学版,1956(1): 25-44.
[6]刘守海,徐兆礼,田丰歌.长江口及附近水域凤鲚摄食习性的分析[J].上海海洋大学学报,2012,21(4):589-597.
[7]刘守海,徐兆礼.长江口和杭州湾凤鲚胃含物与海洋浮游动物的种类组成比较[J].生态学报,2011,31(8):2 263-2 271.
[8]仲伟,邵鑫斌,胡利华,等.凤鲚瓯江种群的生物学特性[J].温州大学学报:自然科学版,2009,30(4):14-18.
[9]管卫兵,陈辉辉,何文辉.长江口凤鲚生殖群体的动态特征[J].渔业科学进展,2011,32(5):1-9.
[10]国家质量监督检验检疫总局.海洋调查规范第六部分:海洋生物调查[M].北京:海洋出版社,2005.
[11]HYSLOPE J.Stomach contents analysis-a review of methods and their application[J].Journal of Fish Biology,1980,17(4): 411-429.
[12]窦硕增,杨纪明.渤海南部半滑舌鳎的食性及摄食的季节性变化[J].生态学报,1992,12(4):368-376.
[13]KREBS C J.Ecological methodology[M].New York:Harper Collins Publishers,1989:654.
[14]张力.SPSS在生物统计中的应用[M].厦门:厦门大学出版社,2008.
[15]蒋日进,金海卫,周永东,等.东海竹筴鱼的食性[J].应用生态学报,2013,24(7):2 015-2 024.
[16]颜云榕,陈骏岚,侯 刚,等.北部湾带鱼的摄食习性[J].应用生态学报,2010,21(3):749-755.
[17]张波.黄海中部高眼鲽的摄食及随体长的变化[J].应用生态学报,2007,18(8):1 849-1 854.
[18]薛莹,金显仕,张波,等.黄海中部小黄鱼摄食习性的体长变化与昼夜变化[J].中国水产科学,2004,11(5):420-425.
[19]MORATO T,SANTOS R S,ANDRADE J P.Feeding habits,seasonal and ontogenetic diet shift of blacktail comber(Serranus atricauda)(Pisces:Serranidae),from the Azores,north-eastern Atlantic[J].Fisheries Research,2000,49(1):51-59.
[20]HOVDE S C,ALBERT O T,NILSSEN E M.Spatial,seasonal and ontogenetic variation in diet of Northeast Arctic Greenland halibut(Reinhardtius hippoglossoides)[J].ICES Journal of Marine Science,2002,59(2):421-437.
[21]LUKOSCHEK V,MCCORMICK M I.Ontogeny of diet changes in a tropical benthic carnivorous fish,Parupeneus barberinus (Mullidae):relationship between foraging behaviour,habitat use,jaw size,and prey selection[J].Marine Biology,2001,138(6): 1 099-1 113.
[22]张波.东、黄海带鱼的摄食习性及随发育的变化[J].海洋水产研究,2004,25(2):6-12.
[23]贺舟挺,张亚洲,徐开达,等.东、黄海黄鮟鱇的摄食习性及其随发育的变化[J].渔业科学进展,2012,33(2):11-20.
[24]蒋日进,徐汉祥,金海卫,等.东海蓝圆鲹的摄食习性[J].水产学报,2012,36(2):216-227.
[25]GERKING S D.Feeding ecology of fish[M].San Diego:Academic Press,1994.
中图分类号:Q178.53
文献标识码:A
文章编号:1008-830X(2016)01-0008-07
收稿日期:2015-10-14 基金项目:农业部重点实验室[农计发(2015)9号];浙江省科技厅培育创新载体项目(2015F10030);海洋捕捞渔业转型升级项目[浙海渔计(2013)113号]
作者简介:郭爱(1982-),男,湖北黄梅人,博士研究生,研究方向:海洋渔业资源、渔业遥感.E-mail:guoaiai@hotmail.com 通讯作者:周永东,男,安徽安庆人,教授级高工,研究方向:渔业资源.E-mail:zyd511@126.com
The Food Variation in Different Ontogenetic Stages of Coilia mystus in the East China Sea and Southern Yellow Sea
GUO Ai1,2,3,CHEN Feng1,ZHU Wen-bin1,et al
(1.Marine Fisheries Institute of Zhejiang Province,Scientific Observing and Experimental Station of Fishery Resources for Key Fishing Grounds,Ministry of Agriculture,Key Laboratory of Sustainable Utilization of Technology Research for Fishery Resource of Zhejiang Province,Zhoushan316021;2.College of Marine Science and Technology,Shanghai Ocean University,Shanghai201306;3.The Second Institute of Oceanography,SOA,Hangzhou310012,China)
Abstract:Based on samples from the voyages in the East China Sea and southern Yellow Sea(26°00′-35° 00′N and 120°00′-126°30′E),the food composition,feeding intensity,niche breadth of the food,and weight
variation of single species in the food mass of Coilia mystus in different ontogenetic stages were analyzed,and C.mystus in different size classes were also analyzed by clustering.The results showed that the food composition of C.mystus varied in different ontogenetic stages.According to the index of relative importance(IRI)of the food components,copepods were the most important in the food spectrum in the whole ontogeny,and then were mysids and euphausids.The IRI percentage of small fishes in the food composition increased with the increase of the body length.The feeding intensity had two peaks in the juvenile group(body length 70-100 mm)and the adult group(191-250 mm),with mean stomach fullness index higher than 10‰.There was a negative correlation between the percentage of empty stomachs and the body length,i.e.,the percentage of empty stomachs decreased with the increase of the body length.In the food mass,the mean weight of single ingredient increased with the increase of the body length,showing a positive correlation.All the mean weights of single ingredient were lower than 10 mg in the stage of body length 70-160 mm,and higher than 30 mg in the stage of body length 161-250 mm.The results of clustering analysis showed that C.mystus in six size classes could be divided into 3 groups,body length 70-100 mm,101-130 mm,and 131-250 mm,with different food composition.C.mystus mainly fed on copepods and larvae in body length 70-100 mm,copepods,mysids and euphausids in body length 101-130 mm,and copepods,mysids and fishes in body length 131-250 mm.The niche breadth of the food also varied in different ontogenetic stages of C.mystus.The diversity index of the food was the lowest in body length 70-100 mm and the highest in body length 101-130 mm,and didn't vary much in body length of 131-250 mm.
Key words:East China Sea and Yellow Sea;Coilia mystus;development stage;feeding;%IRI
猜你喜欢
作物学报(2022年7期)2022-05-11
当代水产(2021年10期)2022-01-12
华人时刊(2020年21期)2021-01-14
高中生之友(中旬刊)(2019年9期)2019-10-23
中国油脂(2019年4期)2019-04-29
科学与财富(2017年26期)2017-09-23
教育教学论坛(2016年49期)2017-02-27
河北渔业(2014年11期)2014-11-17
江苏农业科学(2014年9期)2014-11-15
果树学报(2013年2期)2013-05-07
