
金叶银杏半同胞子代无性系的叶色和色素含量变化及呈色机制分析
2016-08-12 01:13郁万文祝遵凌曹福亮周春丽
植物资源与环境学报 2016年1期
郁万文, 祝遵凌, 曹福亮, 周春丽
(1. 南京林业大学南方现代林业协同创新中心, 江苏 南京 210037; 2. 江苏苏林园林工程有限公司, 江苏 宿迁 223600)
金叶银杏半同胞子代无性系的叶色和色素含量变化及呈色机制分析
郁万文1,2, 祝遵凌1, 曹福亮1, 周春丽1
(1. 南京林业大学南方现代林业协同创新中心, 江苏 南京 210037; 2. 江苏苏林园林工程有限公司, 江苏 宿迁 223600)
对4月份至11月份金叶银杏‘万年金’(Ginkgobiloba‘Wannianjin’)32个半同胞子代无性系与亲本的叶色差异进行比较;比较了不同色系叶片的色素含量和比值及叶色参数(L*、a*和b*)的变化,分析了叶色参数与叶片色素含量的相关性;并观察了不同色系的叶绿体超微结构。结果表明:32个半同胞子代无性系可被分为金黄、浅黄、草绿和蓝绿4个色系。随时间推移,草绿和蓝绿色系叶片的总叶绿素(Chl)、叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)含量均呈“双峰型”变化趋势,Car/Chl和Car/Chla比值的变幅均较小;而金黄和浅黄色系叶片的上述色素含量呈“升高—降低—升高”变化趋势,Car/Chl和Car/Chla比值总体呈“迅速下降—相对稳定—缓慢升高”的变化趋势。各色系叶片的上述色素含量在夏季均不同程度下降,Car/Chlb比值变化差异较大,且金黄和浅黄色系的各色素含量均低于草绿和蓝绿色系。随时间推移,金黄和浅黄色系叶片的L*、a*和b*值以及草绿和蓝绿色系叶片的L*和b*值均先降低后升高,后2个色系的a*值则先升高后下降;并且,前2个色系的L*和b*值总体上显著高于后2个色系,而a*值则总体上低于后2个色系。金黄色系的Chla和Chl含量与L*和a*值显著负相关,而其Car/Chl和Car/Chla比值则与L*、a*和b*值显著或极显著正相关;浅黄色系的Chlb含量与a*值显著负相关,其Car/Chla比值与L*和b*值以及Car/Chlb比值与a*值均显著正相关;草绿色系的Chla含量与L*值显著负相关,其Car/Chla比值与L*和b*值以及Car/Chlb比值与a*值均显著正相关;这3个色系叶片的其余指标间以及蓝绿色系叶片的各指标间均无显著相关性。观察结果显示:金黄和浅黄色系的叶绿体基粒片层发育不健全,基粒片层可见但排列较疏松,且无明显垛叠,分布范围小而稀疏;蓝绿和草绿色系叶绿体的基粒类囊体垛叠层数均较多,基粒片层发达且排列紧致、整齐,分布范围大而稠密。综合分析结果表明:‘万年金’4个色系半同胞子代无性系叶片的呈色差异和叶色变化由多种因素控制,其中,Car/Chl和Car/Chla比值高且叶绿体基粒片层发育不健全是叶片呈黄色的主要原因。
金叶银杏; 叶色; 色素含量; 叶色参数; 超微结构; 呈色机制
传统的广普性绿化植物色彩单一、绿化形式单调,而彩叶植物多数具有独特的叶色,宜作为园林绿化植物的首选,在园林配置中的应用也越来越广。目前,人们已经成功选育出金叶国槐(Sophorajaponicaf.flavi-rameus)、金叶榆(Ulmuspumila‘Jinye’)、金叶水杉(Metasequoiaglyptostroboides‘Gold Rush’)、金叶黄栌(Cotinuscoggygria‘Golden Spirit’)、黄金榕(Ficusmicrocarpa‘Golden Leaves’)、金叶刺槐(Robinapseudoacaa‘Frilis’)、皇冠栾(Koelreuteriabipinnata‘Huangguan’)等多种金叶植物,并有针对性地开展了一系列繁育研究。然而,对彩叶植物生态生理学特性,特别是叶色形成机制方面的研究尚不多见,且对彩叶植物叶色动态变化及呈色机制的研究也无统一结论。何亦昆等[1]认为,彩叶植物的叶片呈色与叶肉细胞所含色素的种类、相对含量及其在叶片中的分布和组织定位有关,尤其是叶肉细胞中的色素种类及其相对比例的变化和时空组合最终决定了叶片呈色[2-3]。Tsuzuki等[4]认为,苗期显现的叶色变异稳定、便于观察且易于检测,是呈色机制研究的有效载体。
金叶银杏‘万年金’(‘Wannianjin’)为银杏(GinkgobilobaLinn.)的1个芽变品种,由南京林业大学选育,其叶色在春季即为金黄色,具有很高的观赏价值。随着叶龄的增大,‘万年金’的叶色由金黄色逐渐转为黄绿色,并在盛夏出现“返绿”现象,观赏价值随之降低。观察结果显示,由‘万年金’种子繁育出的实生苗出现明显的叶色分化,有黄色和绿色之分,且不同单株间叶色深浅不一。目前,关于金叶银杏的研究主要集中在组织培养[5]和扦插育苗[6]等方面,笔者在前期对‘万年金’的叶色发育、光合特性、选育和区试等也进行了探索性研究[7-8],然而关于‘万年金’叶片的呈色机制尚不明确。
为了弄清金叶银杏‘万年金’半同胞子代无性系的叶色分化情况,明确不同色系的叶色发育规律和呈色机制,作者对‘万年金’4个色系半同胞子代无性系叶片的色素含量和叶色参数(包括L*、a*和b*)的变化及叶绿体的超微结构进行了比较,以期为金叶植物的呈色机制研究和良种选育提供指导和借鉴。
1 材料和方法
1.1材料
于2006年9月下旬采集‘万年金’的成熟种实,经常规调制后将得到的种子置于4 ℃冰箱中保存、备用;翌年春天进行催芽和播种,共获得32株实生苗;于2010年3月下旬对所有实生苗进行剪枝嫁接,形成32个半同胞子代无性系,依次编号1至32,其中,26号无性系为双胚实生苗,一个主干上的叶片呈浅黄色,记作26H,另一个主干上的叶片呈草绿色,记作26L;另采集亲本的普通枝条和金叶芽变枝条进行嫁接,形成2个亲本无性系,分别记作CK1和CK2。将各无性系按照编号顺序种植于南京林业大学银杏园内,株距0.5 m、行距1.0 m,采取常规日常管理方式培育1 a。
南京林业大学银杏园地处亚热带和暖温带过渡地区,属亚热带季风气候区,年均温15.4 ℃,最高月均温28.1 ℃,最低月均温-1.5 ℃,年降水量1 106 mm,年均空气相对湿度76%,土壤的最大冻结深度为0.09 m,无霜期232 d。土壤为黄棕壤,0~30 cm土层为沙壤土,pH 7.36,土壤中有机质、全氮、全磷和全钾的含量分别为26.85、1.17、0.49和4.64 g·kg-1,碱解氮、速效磷和速效钾的含量分别为46.27、9.78和95.2 mg·kg-1。
1.2方法
1.2.1叶色参数的测定参照丁廷发等[9]的方法测定叶色参数,具体操作略有改动。分别于2011年4月20日、 5月20日、 6月20日、 7月25日、 9月 5日、9月20日、10月20日和11月20日的10:00左右随意采集各无性系植株顶芽下第4至第6位成熟、完整的功能叶1枚,平放在桌面上,将英国皇家比色卡置于叶旁,使用 SONY DSC-HX1数码相机记录叶片的实际视觉场景,叶片与相机镜头的间距为60 cm;以叶柄为轴将叶片分为左右两部分,用Photoshop CS 8.01软件将各部分中部的叶色转化成Lab模式(其中,L*表示黑白亮度,值阈为0~100,数值越小颜色越暗;a*表示红绿之间的变化,值阈为-120~120,正值偏红,负值偏绿;b*表示黄蓝之间的变化,数值越大颜色越黄,数值越小颜色越蓝),每个无性系各检测4株,视为4个重复。描绘32个半同胞子代无性系在观赏期和全年的叶色差异彩块图,并采用SPSS 22.0统计分析软件进行不同色系叶色参数与色素含量间的相关性分析。
1.2.2叶片色素含量的测定参照熊庆娥[10]的方法测定叶片色素含量,略有改动。在每个无性系供拍照的叶片中随机选择1枚叶片,按照不同色系叠加,剪成长3 mm、宽1 mm的碎片,混合后称取0.05 g,加入10 mL丙酮-乙醇(等体积比)混合液,避光并封口浸提24 h;取上清液,使用岛津UV-1800紫外可见分光光度计(日本岛津公司)测定光密度值,并计算叶片的叶绿素a、叶绿素b、总叶绿素及类胡萝卜素含量,每个指标重复测定4次。
1.2.3叶绿体超微结构的观察参照Ruppel等[11]的方法制作超微切片,略有改动。于叶片成熟初期(即5月20日),根据各色系的L*、a*和b*的均值,选取L*、a*和b*值均接近均值的19号(金黄色系)、20号(蓝绿色系)、22号(草绿色系)和31号(浅黄色系)子代无性系以及普通亲本无性系(CK1)和金叶芽变亲本无性系(CK2),取正常生长植株顶芽下的第5枚叶片,切成宽0.1 cm长条,用戊二醛和锇酸固定,并经体积分数70%、80%、90%、100%乙醇系列脱水,用包埋剂渗透后用Epon812环氧树脂包埋,切成超薄切片;先后用醋酸双氧铀和柠檬酸铅进行双重染色,自然干燥后用HITACHI 7650透射电镜(日本日立高新技术公司)观察并拍照。每个无性系选择3个玻片进行观察,选择最佳照片进行分析。
1.3数据处理和统计分析
将最佳观赏期32个子代无性系的比色结果直接导入Photoshop CS 8.01软件,统计各子代无性系叶色参数L*、a*和b*,并输出RGB最佳颜色模式制作成反映真实叶色的色块;采用K-means方法对各子代无性系的叶色参数L*、a*和b*进行聚类。采用EXCEL 2013和SPSS 22.0软件对相关数据进行差异显著性分析和相关性分析。
2 结果和分析
2.1观赏期半同胞子代无性系叶色的比较分析
金叶银杏‘万年金’32个半同胞子代无性系及2个亲本无性系叶色的比较结果见图1。结果显示:金叶银杏‘万年金’32个半同胞子代无性系叶片呈现深浅不一的黄色或绿色,说明其子代实生苗的叶色出现明显的遗传分化。
K-means聚类结果显示,金叶银杏‘万年金’32个半同胞子代无性系被划分为金黄、浅黄、草绿和蓝绿4个色系,与比色卡的比色结果相符。其中,金黄色系对应的色卡区域为Yellow-green group N144A和151A,包括16、19、21和24号半同胞子代无性系,具有较高的观赏价值;浅黄色系对应的色卡区域为Yellow-green group N144A、144C和145A,包括1、3、4、5、6、7、8、9、10、11、17、18、25、26H、31和32号半同胞子代无性系,具有一定的观赏性;草绿色系对应的色卡区域为Green group 137C~138B,包括2、15、22、23、26L、27、28、29和30号半同胞子代无性系;蓝绿色系对应的色卡区域为Green group 137C~138A,包括12、13、14和20号半同胞子代无性系。
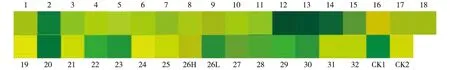
1-32: 半同胞子代无性系,其中,26H和26L为双胚实生苗的2个主干 Half-sib progeny clones, in which, 26H and 26L are two trunks of double embryo seedling; CK1: 普通亲本无性系 Common parent clone; CK2: 金叶芽变亲本无性系 Parent clone of gold leaf bud mutation.
图1金叶银杏‘万年金’32个半同胞子代无性系及其亲本无性系叶色的比较(以4月20日的比较结果为例)
Fig. 1Comparison on leaf color of 32 half-sib progeny clones of ‘Wannianjin’ of golden-leaf ginkgo (GinkgobilobaLinn.) and
their parent clones (taking comparison result in April 20 as an example)
2.2不同色系半同胞子代无性系叶片色素含量的比较分析
2.2.1色素含量的比较金叶银杏‘万年金’不同色系半同胞子代无性系叶片的色素含量见表1。结果表明:在4月20日至11月20日期间,草绿和蓝绿色系无性系的总叶绿素(Chl)、叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)含量均随时间推移呈现“双峰型”变化趋势,且二者的Chl、Chla、Chlb和Car含量基本无显著差异(P>0.05),并在4月20日至6月20日期间显著高于金黄和浅黄色系无性系(P<0.05);而金黄和浅黄色系无性系的Chl、Chla、Chlb和Car含量则随时间推移呈现“升高—降低—升高”的变化趋势,并且这2个色系叶片的Chl、Chla、Chlb和Car含量均较低且无差异显著。
总体而言,草绿和蓝绿色系无性系叶片的色素含量峰值多出现在5月20日,而金黄和浅黄色系无性系的峰值则出现在7月25日;夏季(7月25日至9月5日)各色系叶片的色素含量均不同程度下降;进入秋季后,各色系叶片的色素含量又不同程度升高;11月份时,草绿和蓝绿色系叶片的色素含量均有所下降,而金黄和浅黄色则略有升高。
2.2.2类胡萝卜素与叶绿素含量比值的比较金叶银杏‘万年金’不同色系半同胞子代无性系叶片中类胡萝卜素(Car)与叶绿素a(Chla)、叶绿素b(Chlb)和总叶绿素(Chl)含量的比值见表2。结果表明:在4月20日至11月20日期间,金黄和浅黄色系无性系叶片的Car/Chl和Car/Chla比值随时间推移的变化趋势较为一致,总体表现为在生长前期(6月20日前)迅速降低、生长中期(6月20日至9月5日)相对平衡、生长后期(9月5日后)缓慢升高的趋势;草绿和蓝绿色系无性系叶片的Car/Chl和Car/Chla比值的变化趋势也较为一致,均表现为小幅波动。然而,各色系叶片的Car/Chlb比值变化趋势有较大差异,金黄色系叶片的Car/Chlb比值表现为在生长前期迅速降低并于6月20日降至最低值后逐渐升高,而蓝绿色系的变化趋势大体上与金黄色系相反;浅黄色系叶片的Car/Chlb比值表现为在生长前期迅速降低、生长中期维持在相对平衡的状态、生长后期升高并维持在相对平稳的水平,而草绿色系的变化趋势与浅黄色系恰好相反。
表1金叶银杏‘万年金’不同色系半同胞子代无性系叶片中色素含量的变化1)
Table 1Change in pigment contents in leaf of different color series of half-sib progeny clones of ‘Wannianjin’ of golden-leaf ginkgo (GinkgobilobaLinn.)1)

色系Colorseries不同日期(MM-DD)叶片的叶绿素a含量/mg·g-1 Chlorophyllacontentinleafatdifferentdates(MM-DD)04-2005-2006-2007-2509-0509-2010-2011-20金黄色Goldenyellow0.241±0.068b0.651±0.109b0.869±0.050b0.969±0.086a0.798±0.110bc0.579±0.047a0.672±0.045b0.762±0.056a浅黄色Lightyellow0.303±0.088b0.656±0.095b0.846±0.050b0.946±0.237a0.473±0.046c0.564±0.109a0.782±0.043ab0.867±0.106a草绿色Grassgreen1.240±0.125a1.489±0.063a1.614±0.150a1.314±0.054a1.207±0.115a0.977±0.189a1.355±0.276a0.804±0.343a蓝绿色Blue-green1.308±0.227a1.762±0.046a1.670±0.217a1.237±0.163a0.911±0.017ab0.945±0.143a1.231±0.124ab0.983±0.389a

色系Colorseries不同日期(MM-DD)叶片的叶绿素b含量/mg·g-1 Chlorophyllbcontentinleafatdifferentdates(MM-DD)04-2005-2006-2007-2509-0509-2010-2011-20金黄色Goldenyellow0.074±0.016b0.170±0.035c0.240±0.001b0.238±0.035a0.167±0.022b0.123±0.006a0.112±0.020b0.152±0.009a浅黄色Lightyellow0.119±0.033b0.252±0.012bc0.276±0.004b0.269±0.045a0.179±0.025b0.129±0.018a0.165±0.019b0.203±0.024a草绿色Grassgreen0.446±0.034a0.514±0.086ab0.342±0.017a0.309±0.024a0.261±0.003a0.256±0.063a0.382±0.048a0.285±0.114a蓝绿色Blue-green0.425±0.081a0.587±0.119a0.350±0.011a0.329±0.017a0.296±0.025a0.246±0.094a0.434±0.057a0.321±0.133a
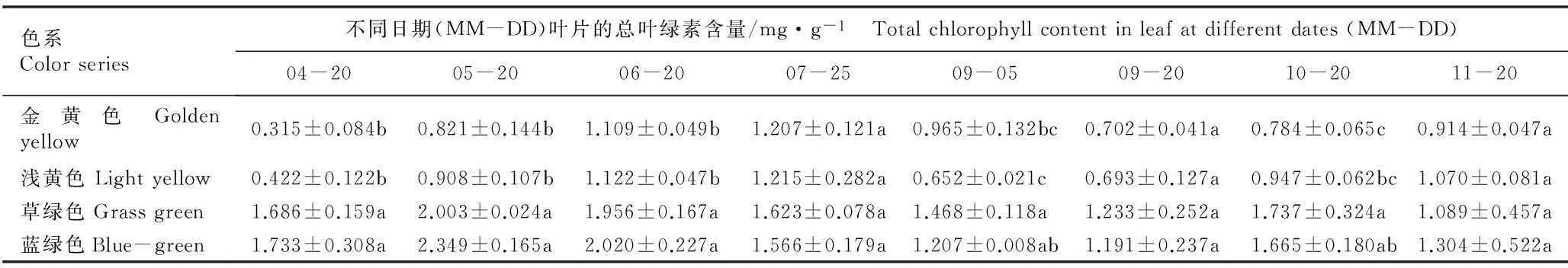
色系Colorseries不同日期(MM-DD)叶片的总叶绿素含量/mg·g-1 Totalchlorophyllcontentinleafatdifferentdates(MM-DD)04-2005-2006-2007-2509-0509-2010-2011-20金黄色Goldenyellow0.315±0.084b0.821±0.144b1.109±0.049b1.207±0.121a0.965±0.132bc0.702±0.041a0.784±0.065c0.914±0.047a浅黄色Lightyellow0.422±0.122b0.908±0.107b1.122±0.047b1.215±0.282a0.652±0.021c0.693±0.127a0.947±0.062bc1.070±0.081a草绿色Grassgreen1.686±0.159a2.003±0.024a1.956±0.167a1.623±0.078a1.468±0.118a1.233±0.252a1.737±0.324a1.089±0.457a蓝绿色Blue-green1.733±0.308a2.349±0.165a2.020±0.227a1.566±0.179a1.207±0.008ab1.191±0.237a1.665±0.180ab1.304±0.522a
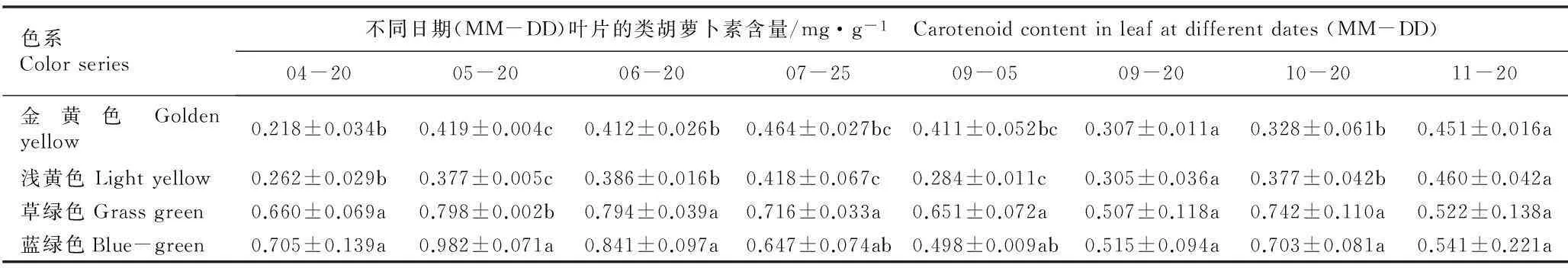
色系Colorseries不同日期(MM-DD)叶片的类胡萝卜素含量/mg·g-1 Carotenoidcontentinleafatdifferentdates(MM-DD)04-2005-2006-2007-2509-0509-2010-2011-20金黄色Goldenyellow0.218±0.034b0.419±0.004c0.412±0.026b0.464±0.027bc0.411±0.052bc0.307±0.011a0.328±0.061b0.451±0.016a浅黄色Lightyellow0.262±0.029b0.377±0.005c0.386±0.016b0.418±0.067c0.284±0.011c0.305±0.036a0.377±0.042b0.460±0.042a草绿色Grassgreen0.660±0.069a0.798±0.002b0.794±0.039a0.716±0.033a0.651±0.072a0.507±0.118a0.742±0.110a0.522±0.138a蓝绿色Blue-green0.705±0.139a0.982±0.071a0.841±0.097a0.647±0.074ab0.498±0.009ab0.515±0.094a0.703±0.081a0.541±0.221a
1)同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
表2金叶银杏‘万年金’不同色系半同胞子代无性系叶片中类胡萝卜素与叶绿素含量比值的变化1)
Table 2Change in ratios of carotenoid content to chloraphyll content in leaf of different color series of half-sib progeny clones of ‘Wannianjin’of golden-leaf ginkgo (GinkgobilobaLinn.)1)

色系Colorseries不同日期(MM-DD)叶片的Car/Chla比值 Car/Chlaratioinleafatdifferentdates(MM-DD)04-2005-2006-2007-2509-0509-2010-2011-20金黄色Goldenyellow0.905±0.130a0.645±0.085a0.474±0.002a0.479±0.018a0.515±0.007b0.530±0.021a0.488±0.052a0.591±0.027a浅黄色Lightyellow0.864±0.145a0.574±0.059a0.457±0.007a0.442±0.079a0.600±0.030a0.540±0.029a0.482±0.025a0.531±0.022a草绿色Grassgreen0.533±0.002a0.536±0.022a0.492±0.026a0.545±0.003a0.540±0.010ab0.519±0.014a0.548±0.051a0.650±0.057a蓝绿色Blue-green0.539±0.009a0.557±0.027a0.504±0.010a0.523±0.012a0.547±0.001ab0.545±0.013a0.571±0.010a0.550±0.004a
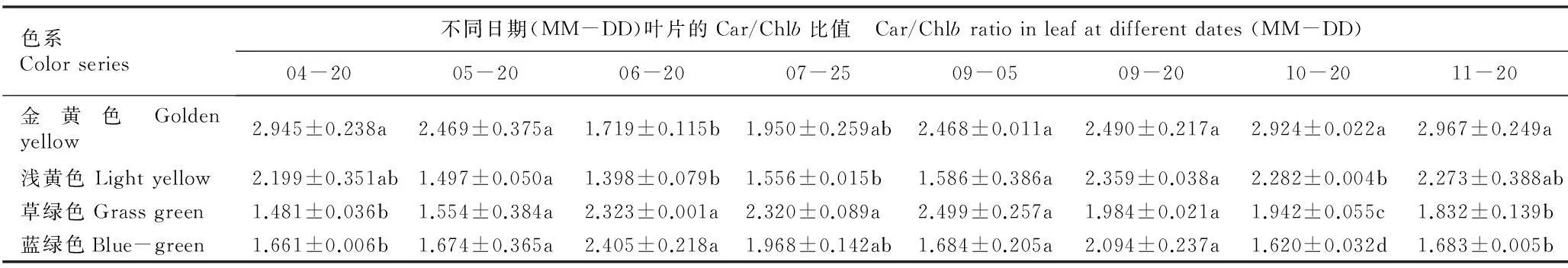
色系Colorseries不同日期(MM-DD)叶片的Car/Chlb比值 Car/Chlbratioinleafatdifferentdates(MM-DD)04-2005-2006-2007-2509-0509-2010-2011-20金黄色Goldenyellow2.945±0.238a2.469±0.375a1.719±0.115b1.950±0.259ab2.468±0.011a2.490±0.217a2.924±0.022a2.967±0.249a浅黄色Lightyellow2.199±0.351ab1.497±0.050a1.398±0.079b1.556±0.015b1.586±0.386a2.359±0.038a2.282±0.004b2.273±0.388ab草绿色Grassgreen1.481±0.036b1.554±0.384a2.323±0.001a2.320±0.089a2.499±0.257a1.984±0.021a1.942±0.055c1.832±0.139b蓝绿色Blue-green1.661±0.006b1.674±0.365a2.405±0.218a1.968±0.142ab1.684±0.205a2.094±0.237a1.620±0.032d1.683±0.005b
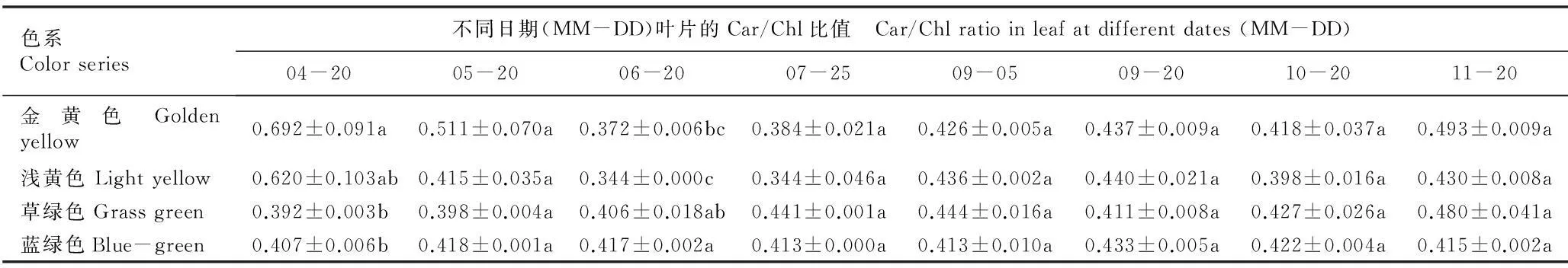
色系Colorseries不同日期(MM-DD)叶片的Car/Chl比值 Car/Chlratioinleafatdifferentdates(MM-DD)04-2005-2006-2007-2509-0509-2010-2011-20金黄色Goldenyellow0.692±0.091a0.511±0.070a0.372±0.006bc0.384±0.021a0.426±0.005a0.437±0.009a0.418±0.037a0.493±0.009a浅黄色Lightyellow0.620±0.103ab0.415±0.035a0.344±0.000c0.344±0.046a0.436±0.002a0.440±0.021a0.398±0.016a0.430±0.008a草绿色Grassgreen0.392±0.003b0.398±0.004a0.406±0.018ab0.441±0.001a0.444±0.016a0.411±0.008a0.427±0.026a0.480±0.041a蓝绿色Blue-green0.407±0.006b0.418±0.001a0.417±0.002a0.413±0.000a0.413±0.010a0.433±0.005a0.422±0.004a0.415±0.002a
1)Chla: 叶绿素a含量 Chlorophyllacontent; Chlb: 叶绿素b含量 Chlorophyllbcontent; Chl: 总叶绿素含量 Total chlorophyll content; Car: 类胡萝卜素含量 Carotenoid content. 同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2.3不同色系半同胞子代无性系叶色及其参数的比较分析
在4月20日至11月20日期间金叶银杏‘万年金’不同色系半同胞子代无性系的叶色变化见图2;各色系的叶色参数变化见表3。
由图2可见:4个色系的叶色均为生长初期偏黄、生长中期渐绿、生长后期渐黄,并且,同一时期金黄和浅黄色系的叶色较草绿和蓝绿色系偏黄。
由表3可见:4个色系叶色参数L*、a*和b*值的变化规律相似。总体上看,金黄和浅黄色系的L*、a*和b*值以及草绿和蓝绿色系的L*和b*值均呈现先降低后升高的变化趋势,而草绿和蓝绿色系的a*值则表现为先升高后下降的变化趋势。金黄和浅黄色系的L*和b*值总体上显著高于草绿和蓝绿色系(P<0.05),并且前二者的L*和b*值在9月5日前后降至最低,而后二者的L*和b*值则在7月25日降至最低;生长中期(6月20日至9月20日),金黄和浅黄色系的a*值基本上显著低于草绿和蓝绿色系,并且前二者的a*值在7月25日前后降至最低,而后二者的a*值则在7月25日前后升至最高。

图2 金叶银杏‘万年金’不同色系半同胞子代无性系叶色的变化
表3金叶银杏‘万年金’不同色系半同胞子代无性系叶色参数(L*、a*和b*)的变化1)
Table 3Change in leaf color parameters (L*, a* and b*) of different color series of half-sib progeny clones of ‘Wannianjin’ of golden-leaf ginkgo (GinkgobilobaLinn.)1)

色系Colorseries不同日期(MM-DD)叶片的L*值 L*valueofleafatdifferentdates(MM-DD)04-2005-2006-20 07-2509-0509-2010-2011-20金黄色Goldenyellow66.86±2.95a60.97±3.14a54.70±1.45a 51.80±1.39a49.02±1.41a51.85±2.08a56.00±0.65a57.29±1.84a浅黄色Lightyellow61.40±1.12a59.17±2.20a54.78±1.21a52.36±1.56a49.23±0.23a49.68±1.89a53.45±1.05a55.55±1.81a草绿色Grassgreen41.56±2.88b35.81±0.75b34.31±0.36b33.58±1.14b35.87±0.78c37.43±0.82b39.07±4.45b47.97±6.08a蓝绿色Blue-green43.18±2.91b37.35±0.43b36.49±0.16b36.16±1.46b39.07±0.01b39.05±0.70b40.44±4.59b49.63±6.14a

色系Colorseries不同日期(MM-DD)叶片的a*值 a*valueofleafatdifferentdates(MM-DD)04-2005-20 06-20 07-2509-05 09-20 10-20 11-20 金黄色Goldenyellow-8.63±1.77a-12.16±3.74a -19.64±0.61c -20.86±0.77b-19.32±0.60b -18.11±1.97b -14.18±0.00a -14.18±1.01a 浅黄色Lightyellow-15.88±1.72b-19.31±0.89a-21.09±0.03c-21.04±0.66b-19.72±0.36b-19.01±1.67b-15.68±0.53a-16.74±1.29a草绿色Grassgreen-16.37±1.69b-12.99±0.73a-11.54±0.13a-11.28±0.55a-12.38±0.63a-11.12±1.08a-13.27±1.16a-15.60±1.30a蓝绿色Blue-green-17.30±1.49b-14.31±0.21a-13.90±0.52b-12.87±0.20a-13.27±0.39a-14.05±0.54ab-15.12±0.28a-15.67±0.68a

色系Colorseries不同日期(MM-DD)叶片的b*值 b*valueofleafatdifferentdates(MM-DD)04-2005-2006-20 07-2509-0509-2010-2011-20金黄色Goldenyellow54.29±0.30a53.69±7.85a38.00±1.29a 35.42±2.14a31.15±0.68a32.50±3.03a38.56±0.30a39.16±7.27a浅黄色Lightyellow53.35±4.03a45.30±2.28a 40.74±2.34a36.06±2.26a31.55±0.44a32.42±2.29a37.00±0.97a38.95±3.17a草绿色Grassgreen23.96±4.05b15.86±1.14b 13.58±0.19b13.95±0.45b14.85±0.82c16.50±0.63b17.75±6.11b29.96±7.05a蓝绿色Blue-green27.30±4.16b18.99±0.63b 17.72±0.88b15.96±0.89b17.75±0.78b19.30±1.14b21.58±4.75b31.08±6.05a
1)同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2.4不同色系半同胞子代无性系叶色参数与叶片色素含量的相关性分析
金叶银杏‘万年金’不同色系半同胞子代无性系叶色参数与叶片色素含量的相关性分析结果见表4。结果表明:各色系的叶色参数与叶片色素含量均呈现一定相关性,其中,金黄和浅黄色系叶片的Car/Chl、Car/Chla和Car/Chlb比值与叶色参数L*、a*和b*值均呈正相关。金黄色系叶片的Chla和Chl含量与L*和a*值呈显著负相关,而其Car/Chl和Car/Chla比值则与L*、a*和b*值呈显著或极显著正相关,其余指标间均无显著相关性。浅黄色系叶片的Chlb含量与a*值呈显著负相关,其Car/Chla比值与L*和b*值呈显著正相关,Car/Chlb比值与a*值呈显著正相关,其余指标间均无显著相关性,Car/Chlb比值与L*和b*值呈中度的负相关关系。草绿色系叶片的Chla含量与L*值呈显著负相关,Car/Chla比值与L*和b*值以及Car/Chlb比值与a*值呈显著正相关,其余指标间均无显著相关性。蓝绿色系叶片的各指标间均无显著相关性,但其Car/Chlb比值与L*和b*值呈中度负相关。
表4金叶银杏‘万年金’不同色系半同胞子代无性系叶片色素含量和叶色参数的相关系数1)
Table 4Correlation coefficient between leaf pigment contents and leaf color parameters of different color series of half-sib progeny clones of ‘Wannianjin’ of golden-leaf ginkgo (GinkgobilobaLinn.)1)

叶色参数Leafcolorparameter金黄色系无性系各指标间的相关系数 CorrelationcoefficientamongdifferentindexesofgoldenyellowcolorseriesclonesChlaChlbChlCarCar/ChlCar/ChlaCar/ChlbL*-0.74#-0.43-0.72#-0.52 0.87##0.85##0.34a*-0.81#-0.65-0.80#-0.580.79#0.84#0.60b*-0.60-0.22-0.59-0.370.78#0.76#0.12

叶色参数Leafcolorparameter浅黄色系无性系各指标间的相关系数 CorrelationcoefficientamongdifferentindexesoflightyellowcolorseriesclonesChlaChlbChlCarCar/ChlCar/ChlaCar/ChlbL*-0.32-0.12-0.180.020.530.75#0.22a*-0.39-0.70#-0.44-0.140.640.530.82#b*-0.45-0.18-0.31-0.160.620.84#0.18

叶色参数Leafcolorparameter草绿色系无性系各指标间的相关系数 CorrelationcoefficientamongdifferentindexesofgrassgreencolorseriesclonesChlaChlbChlCarCar/ChlCar/ChlaCar/ChlbL*-0.72#-0.06-0.60-0.610.470.82#-0.51 a*0.35-0.37-0.58-0.610.29-0.570.69#b*-0.690.02-0.55-0.570.420.81#-0.56
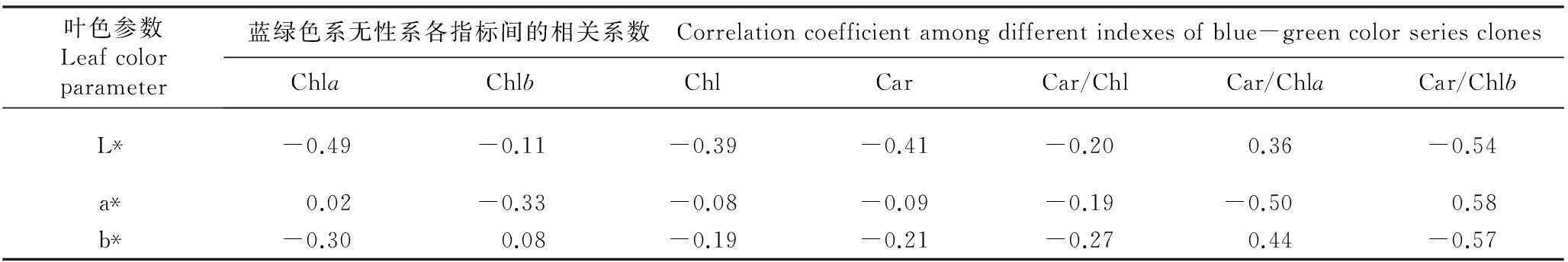
叶色参数Leafcolorparameter蓝绿色系无性系各指标间的相关系数 Correlationcoefficientamongdifferentindexesofblue-greencolorseriesclonesChlaChlbChlCarCar/ChlCar/ChlaCar/ChlbL*-0.49-0.11-0.39-0.41-0.20 0.36 -0.54 a*0.02-0.33-0.08-0.09-0.19-0.500.58b*-0.300.08-0.19-0.21-0.270.44-0.57
1)Chla: 叶绿素a含量 Chlorophyllacontent; Chlb: 叶绿素b含量 Chlorophyllbcontent; Chl: 总叶绿素含量 Total chlorophyll content; Car: 类胡萝卜素含量 Carotenoid content. #:P<0.05; ##:P<0.01.
2.5不同色系半同胞子代无性系叶绿体的超微结构分析
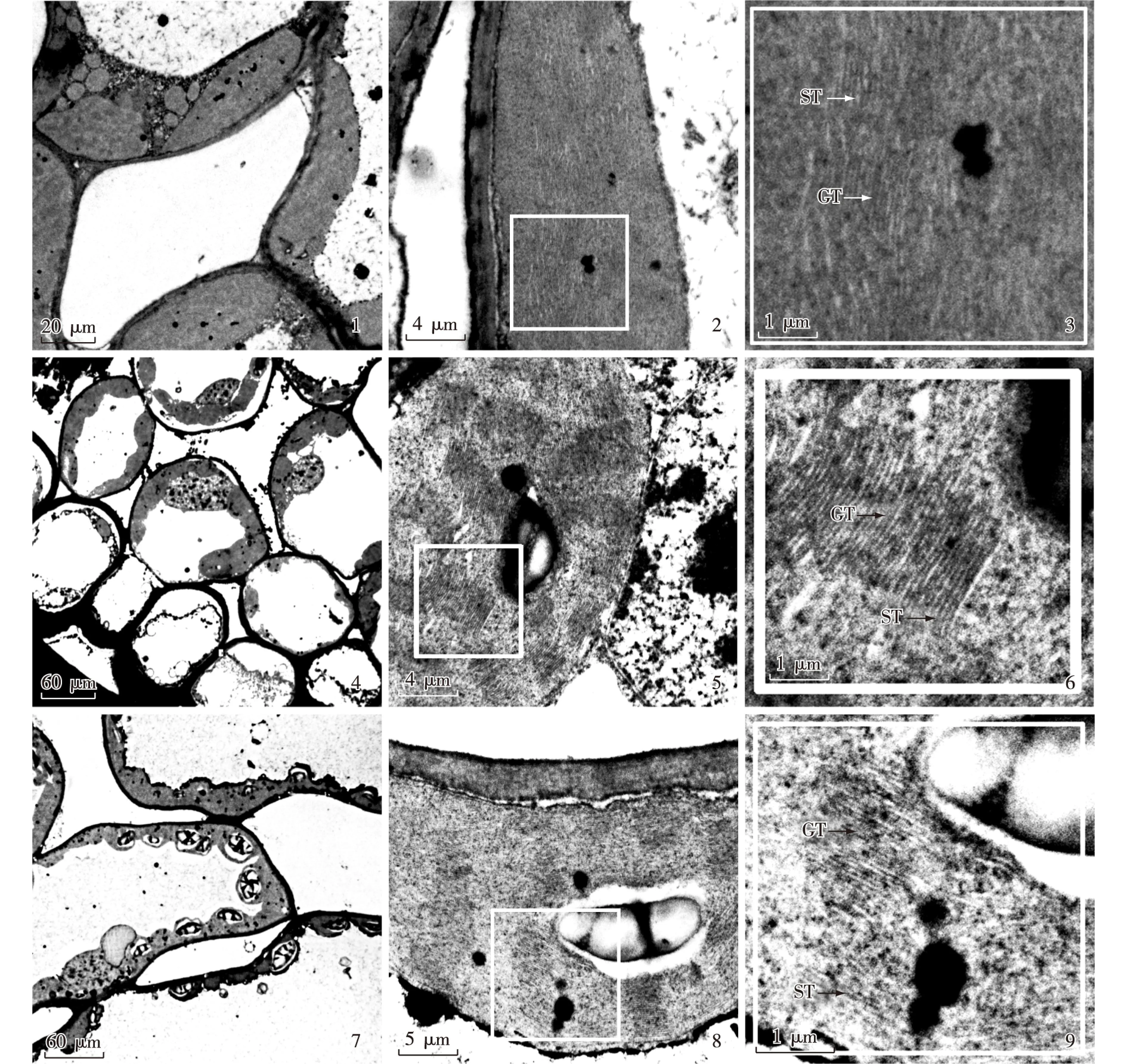
金叶银杏‘万年金’不同色系半同胞子代无性系及普通亲本和金叶芽变亲本无性系叶肉细胞中叶绿体超微结构的观察结果见图版Ⅰ和Ⅱ。
观察结果表明:4个色系半同胞子代无性系及普通亲本和金叶芽变亲本无性系叶绿体的形态及分布基本一致,大多呈椭球形且均环绕在叶肉细胞的内表面;但不同色系叶肉细胞中叶绿体的超微结构存在一定差异,主要表现在基粒数量以及基粒类囊体和基粒片层的垛叠状况等方面。普通亲本无性系 (图版Ⅰ-2,3)以及蓝绿色系 (图版Ⅰ-5,6)和草绿色系(图版Ⅰ-8,9)半同胞子代无性系的叶绿体超微结构类似,基粒类囊体的垛叠层数均较多,并且类囊体发达,排列紧致、整齐,在叶肉细胞中的分布范围大而稠密,基粒之间由基质片层相互交联成网;区别在于蓝绿色系半同胞子代无性系叶片的基粒数量和基粒片层垛叠层数高于普通亲本无性系和草绿色系的半同胞子代无性系。金叶芽变亲本无性系(图版Ⅱ-2,3)以及金黄色系(图版Ⅱ-5,6)和浅黄色系(图版Ⅱ-8,9)半同胞子代无性系叶肉细胞中的叶绿体超微结构类似,基粒片层发育不全,基粒片层可见但排列疏松,无明显垛叠,在叶肉细胞中的分布范围小且稀疏,交联成网状结构的现象不明显,基质片层近平行分布于叶绿体基质中;区别在于浅黄色系半同胞子代无性系叶片的基粒片层垛叠不明显,且垛叠层数较少,而金黄色系半同胞子代无性系和金叶芽变亲本无性系的基粒片层几乎无垛叠现象。
3 讨论和结论
研究结果表明:金叶植物的色素种类及其相对含量的时空变化及叶绿体结构的发育程度均对叶色有影响[12-14];色素是金叶植物体内的主要呈色物质,且受到叶绿体结构发育程度和外部环境因子的共同调控作用[12,15]。通常认为,金叶植物叶片中的Car/Chl比值高于普通叶片[14,16]。在金叶银杏‘万年金’半同胞子代无性系中,金黄和浅黄色系无性系叶片的叶绿素和类胡萝卜素含量基本上均低于草绿和蓝绿色系,且在黄色呈现期前二者叶片的Car/Chl和Car/Chla比值均高于草绿和蓝绿色系,据此推测Car/Chl和Car/Chla比值偏高可能是‘万年金’半同胞子代无性系中金黄和浅黄色系叶片呈现黄色的主要原因之一。与陈芳等[13]和范燕萍等[17]的相关研究结果一致。
本研究中,金叶银杏‘万年金’草绿和蓝绿色系半同胞子代无性系的各色素含量随时间推移均呈明显的“双峰型”曲线,2个峰值分别出现在5月20日和10月20日,这可能与银杏自身的生物学特性有关。众所周知,银杏属于前期生长型树种,在6月下旬停止高生长,这就要求植株在短暂的生长期尤其是速生期内合成并积累大量的碳水化合物,用于组织和器官的构建,因而形成第1个色素合成高峰。
夏季高温和强光能够显著影响植物叶片中色素的合成和相对含量,从而影响叶色[18-21]。在7月25日至9月5日,‘万年金’4个色系半同胞子代无性系叶片的叶绿素和类胡萝卜素含量均下降,其Car/Chl比值降至约0.4,4个色系的叶色最绿,说明‘万年金’4个色系半同胞子代无性系叶片的呈色与色素含量无密切关系,但与Car/Chl比值密切相关。由于存在暗反应能障,导致高温和强光条件下叶绿素捕捉的光能未相应增加,造成机体将过剩的光能从叶绿体传递给O2并生成活性氧,进而损害叶肉细胞的膜系统;而含有共轭双键的类胡萝卜素分子则能保护膜系统免受过剩光能的这种损害作用[12]。本研究中,4个色系半同胞子代无性系的叶绿素和类胡萝卜素含量虽然均有所下降,但后者的降幅相对较小,推测这可能是银杏对夏季高温、强光等逆境的适应性反应。
相关性分析结果表明:‘万年金’4个色系半同胞子代无性系的叶色由Car/Chl、Car/Chla和Car/Chlb比值共同决定,其中,金黄和浅黄色系叶片的Car/Chl和Car/Chla比值与L*、a*和b*值呈显著或中等程度正相关;草绿和蓝绿色系叶片的Car/Chla比值与a*值呈中等程度负相关,Car/Chlb比值与L*和b*值均呈中等程度负相关,说明较高的Car/Chl和Car/Chla比值能够直接反映叶片的黄色程度。
超微观察结果显示:‘万年金’金黄和浅黄色系半同胞子代无性系及金叶芽变亲本无性系的叶绿体基粒片层发育均不完全,基粒片层的排列较疏松且无明显垛叠,且分布范围较窄,这可能是该时期(5月20日)金黄和浅黄色系叶片呈现黄色的结构基础。多数彩叶树种存在返绿现象,这可能与叶绿体结构逐渐发育完善有关。推测,随着叶片的发育,‘万年金’金黄和浅黄色系半同胞子代无性系叶片的叶绿体逐渐发育完全,叶绿素总量逐渐增加,叶色逐渐转绿,能够捕捉到更多的光能进行光合作用,以满足植株生长的需要。这一现象也得到了“对叶色突变体中已经退化或发育不良的叶绿体结构进行重建可使叶片复绿”的研究结论[22-24]的佐证。据此认为这可能是彩叶树种对季节变化和生长节律的一种本能响应。
综上所述,叶片的色素含量及比值、叶绿体结构发育状况及生长环境的光照和温度变化共同引起金叶银杏‘万年金’4个色系半同胞子代无性系叶片的呈色差异及叶色变化,叶片的Car/Chl和Car/Chla比值高且叶绿体基粒片层发育不健全是叶片呈现黄色的主要原因。由于叶色突变是一个复杂的生理现象,因此,应从金叶银杏的遗传学和细胞结构等方面对其叶片呈色机制进行深入研究,同时开展光照、温度、湿度和土壤等环境因子对叶片呈色影响的系统研究。
[1]何亦昆, 代庆阳, 苏学辉. 雁来红叶色转变与超微结构及色素含量的关系[J]. 四川师范学院学报:自然科学版, 1995, 16(3): 195-198.
[2]姜卫兵, 庄猛, 韩浩章, 等. 彩叶植物呈色机理及光合特性研究进展[J]. 园艺学报, 2005, 32(2): 352-358.
[3]赵昶灵, 郭维明, 陈俊愉. 植物花色形成及其调控机理[J]. 植物学通报, 2005, 22(1): 70-81.
[4]TSUZUKI T,TAKAHASHI K,INOUE S-I, et al. Mg-chelatase H subunit affects ABA signaling in stomatal guard cells,but is not an ABA receptor inArabidopsisthaliana[J]. Journal of Plant Research, 2011, 124(4): 527-538.
[5]孙姣. 黄叶银杏的组织培养技术[D]. 南京: 南京林业大学森林资源与环境学院, 2011.
[6]祝凌云. 黄叶银杏硬枝扦插繁殖技术与生根机理研究[D]. 南京: 南京林业大学森林资源与环境学院, 2012.
[7]祝遵凌, 曹福亮, 孟凡军, 等. 黄叶银杏新品种‘万年金’区域化试验[J]. 中南林业科技大学学报, 2012, 32(12): 125-131.
[8]YU W W, CAO F L. Preliminary study on leaf color development and photosynthetic characteristics of golden-leaf ginkgo[J]. Agricultural Science and Technology, 2011, 12(8): 1166-1169, 1173.
[9]丁廷发, 谢必武, 张凤龙. 重庆市5种彩叶植物色素和色彩变化规律研究[J]. 重庆三峡学院学报, 2006, 22(3): 78-80.
[10]熊庆娥. 植物生理学实验教程[M]. 成都: 四川科学技术出版社, 2003.
[11]RUPPEL N J, LOGSDON C A, WHIPPO C W, et al. A mutation inArabidopsisseedling plastid development affects plastid differentiation in embryo-derived tissues during seedling growth[J]. Plant Physiology, 2011, 155(1): 342-353.
[12]朱晓静, 尚爱芹, 杨敏生, 等. 中华金叶榆子代苗光合特性及叶片呈色机制探讨[J]. 西北植物学报, 2014, 34(5): 950-956.
[13]陈芳, 唐秀光, 董倩, 等. 金叶白蜡色素含量与叶色参数的年变化规律研究[J]. 河北农业大学学报, 2012, 35(5): 20-24.
[14]胡海姿, 张睿, 尚爱芹, 等. 金叶植物色素含量对光强的响应[J]. 园艺学报, 2007, 34(3): 717-722.
[15]张建农, 满艳萍, 燕丽萍. 黄化西瓜叶片叶绿体结构与光合作用特性[J]. 果树学报, 2004, 21(1): 50-53.
[16]黄春辉, 高洁, 张晓慧, 等. 黄肉猕猴桃果实发育期间色素变化及呈色分析[J]. 果树学报, 2014, 31(4): 617-623.
[17]范燕萍, 李慧玲, 李浩健. 几种花叶线艺兰叶片色斑色素组成和叶绿体超微结构研究[J]. 华南农业大学学报, 2006, 27(2): 8-12.
[18]郑静静, 杨丽, 苏小雨, 等. 水杨酸对高温强光下小麦叶绿体蛋白酶Deg5和PSⅡ功能的调节作用[J]. 生态学报, 2014, 34(24): 7350-7355.
[19]计玮玮, 邱翠花, 焦云, 等. 高温强光胁迫对砂梨叶片光合作用、D1蛋白和Deg1蛋白酶的影响[J]. 果树学报, 2012, 29(5): 794-799.
[20]刘超, 袁野, 盖树鹏, 等. 强光高温交叉胁迫对牡丹叶片PSⅡ和PSⅠ之间能量传递的影响[J]. 园艺学报, 2014, 41(2): 311-318.
[21]孙永江, 杜远鹏, 翟衡. 高温胁迫下不同光强对‘赤霞珠’葡萄PSⅡ活性及恢复的影响[J]. 植物生理学报, 2014, 50(8): 1209-1215.
[22]何瑞锋, 丁毅, 余金洪, 等. 水稻温敏叶绿素突变体叶片超微结构的研究[J]. 武汉植物学研究, 2001, 19(1): 1-5, 91.
[23]邵继荣, 王玉忠, 刘永胜, 等. 水稻温敏型突变体叶片间断失绿的超微结构[J]. 植物学报, 1999, 41(1): 20-24.
[24]董遵, 刘敬阳, 马红梅, 等. 甘蓝型油菜黄化(苗) 突变体的叶绿素含量及超微结构[J]. 中国油料作物学报, 2000, 22 (3): 27-29, 34.
(责任编辑: 佟金凤)
图版说明Explanation of Plates
图版Ⅰ1-3. 普通亲本无性系: 1. 叶肉细胞显微结构; 2. 叶绿体超微结构; 3. 叶绿体局部超微结构. 4-6. 蓝绿色系半同胞子代无性系(20号无性系): 4. 叶肉细胞显微结构; 5. 叶绿体超微结构; 6. 叶绿体局部超微结构. 7-9. 草绿色系半同胞子代无性系(22号无性系): 7. 叶肉细胞显微结构; 8. 叶绿体超微结构; 9. 叶绿体局部超微结构.
Plate Ⅰ1-3. Common parent clone: 1. Microscopic structure of mesophyll cell; 2. Ultrastructure of chloroplast; 3. Ultrastructure of part of chloroplast. 4-6. Half-sib progeny cloneofblue-greencolorseries(No. 20 clone):4. Microscopicstructureofmesophyllcell;5. Ultrastructureofchloroplast;6. Ultrastructure of part of chloroplast. 7-9. Half-sib progeny clone of grass green color series (No. 22 clone): 7. Microscopic structure of mesophyll cell; 8. Ultrastructure of chloroplast; 9. Ultrastructure of part of chloroplast.
图版Ⅱ1-3. 金叶芽变亲本无性系: 1. 叶肉细胞显微结构; 2. 叶绿体超微结构; 3. 叶绿体局部超微结构. 4-6. 金黄色系半同胞子代无性系 (19号无性系): 4. 叶肉细胞显微结构; 5. 叶绿体超微结构; 6. 叶绿体局部超微结构. 7-9. 浅黄色系半同胞子代无性系(31号无性系): 7. 叶肉细胞显微结构; 8. 叶绿体超微结构; 9. 叶绿体局部超微结构.
Plate Ⅱ1-3. Parent clone of gold leaf bud mutation: 1. Microscopic structure of mesophyll cell; 2. Ultrastructure of chloroplast; 3. Ultrastructure of part of chloroplast. 4-6. Half-sib progeny clone of golden color series (No. 19 clone): 4. Microscopic structure of mesophyll cell; 5. Ultrastructure of chloroplast; 6. Ultrastructure of part of chloroplast. 7-9. Half-sib progeny clone of light yellow color series (No. 31 clone): 7. Microscopic structure of mesophyll cell; 8. Ultrastructure of chloroplast; 9. Ultrastructure of part of chloroplast.
GT: 基粒片层Grana lamella; ST: 基质片层Stromal lamella.
郁万文, 等: 金叶银杏半同胞子代无性系的叶色和色素含量变化及呈色机制分析
图版Ⅰ
YU Wanwen, et al: Analyses on changes in leaf color and pigment contents and
coloring mechanism of half-sib progeny clones of golden-leaf ginkgo
Plate Ⅰ
See the explanation at the end of the text
郁万文, 等: 金叶银杏半同胞子代无性系的叶色和色素含量变化及呈色机制分析
图版Ⅱ
YU Wanwen, et al: Analyses on changes in leaf color and pigment contents and
coloring mechanism of half-sib progeny clones of golden-leaf ginkgo
Plate Ⅱ

See the explanation at the end of the text
Analyses on changes in leaf color and pigment contents and coloring mechanism of half-sib progeny clones of golden-leaf ginkgo
YU Wanwen1,2, ZHU Zunling1, CAO Fuliang1, ZHOU Chunli1
(1. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China; 2. Jiangsu Sulin Garden Engineering Limited Company, Suqian 223600, China),J.PlantResour. &Environ., 2016, 25(1): 43-53
Differences in leaf color of 32 half-sib progeny clones and parents of ‘Wannianjin’ of golden-leaf ginkgo (GinkgobilobaLinn.) from April to November were compared. Changes in leaf pigment contents and ratios and leaf color parameters of (L*, a* and b*) different color series were compared, correlation between leaf color parameters and leaf pigment contents was analyzed, and ultrastructure of chloroplast of different color series was observed. The results show that 32 half-sib progeny clones are divided into four color series including golden yellow, light yellow, grass green and blue-green. As time goes on, contents of total chlorophyll (Chl), chlorophylla(Chla), chlorophyllb(Chlb) and carotenoid (Car) in leaf of grass green and blue-green color series all appear the trend of “double-peak”, change ranges in ratios of Car/Chl and Car/Chlaboth are smaller; while contents of the above pigments in leaf of golden yellow and light yellow color series appear the trend of “increasing-decreasing-increasing”, and ratios of Car/Chl and Car/Chlagenerally appear the trend of “decreasing rapidly-relatively stable-increasing slowly”. Contents of the above pigments in leaf of different color series during summer all decrease at different degrees, difference in their Car/Chlbratio change is large, and pigment contents of golden yellow and light yellow color series are all lower than those of grass green and blue-green color series. As time goes on, L*, a* and b* values in leaf of golden yellow and light yellow color series and L* and b* values in leaf of grass green and blue-green color series all decrease at first and then increase, while a* value of the latter two color series increases at first and then decreases; and L* and b* values of the first two color series are generally significantly higher than those of the latter two color series, while a* value is generally lower than that of the latter two color series. There are significantly negative correlations of Chlaand Chl contents of golden yellow color series with L* and a* values, while significantly orextremelysignificantlypositivecorrelations of its Car/Chl and Car/Chlaratios with L*, a* and b* values. There is a significantly negative correlation of Chlbcontent of light yellow color series with a* value, and significantly positive correlations of its Car/Chlaratio with L* and b* values and Car/Chlbratio with a* value. There is a significantly negative correlation of Chlacontent of grass green color series with L* value, and significantly positive correlations of its Car/Chlaratio with L* and b* values and Car/Chlbratio with a* value. There is no significant correlation among other indexes in leaf of three color series and among indexes in leaf of blue-green color series. The observation result shows that development of grana lamella in chloroplast of golden yellow and light yellow color series is imperfect, which is visible with a loose arrangement, but without obviously stacking, its distribution is small and sparse. Stacking layers of grana thylakoids in chloroplast of blue-green and grass green color series are more, grana lamella is developed with a compact and regular arrangement, its distribution is large and dense. The comprehensive analysis result indicates that coloring difference and leaf color change of four color series of half-sib progeny clones of ‘Wannianjin’ are controlled by many factors, in which, high ratios of Car/Chl and Car/Chlaand imperfect development of grana lamella in chloroplast are the main reasons for appearing yellow leaf.
golden-leaf ginkgo; leaf color; pigment content; leaf color parameters; ultrastructure; coloring mechanism
10.3969/j.issn.1674-7895.2016.01.06
2015-06-09
江苏省“双创计划”企业创新类基金项目; 江苏省“六大人才高峰”项目(2014-NY-045); 江苏高校优势学科建设工程资助项目(PAPD)
郁万文(1979—),男,山东临沂人,博士,讲师,主要从事经济林培育研究工作。
S684.03; S664.3
A
1674-7895(2016)01-0043-11
猜你喜欢
语文世界(小学版)(2021年5期)2021-06-04
中国林副特产(2020年3期)2020-01-15
小资CHIC!ELEGANCE(2019年40期)2019-12-10
新教育论坛(2019年13期)2019-09-10
短篇小说(原创版)(2018年4期)2018-07-22
短篇小说(2018年4期)2018-05-21
天然产物研究与开发(2018年4期)2018-05-07
意林(绘英语)(2018年1期)2018-04-28
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
