
生殖激素对龟类生殖调控的研究进展
2016-08-11 11:33李晓楠
安徽农业科学 2016年17期
李晓楠,方 庆
(川北医学院,四川南充 637100)
生殖激素对龟类生殖调控的研究进展
李晓楠,方 庆*
(川北医学院,四川南充 637100)
迄今为止,已发现有11种生殖激素参与龟类生殖调控。雌二醇和睾酮在促性腺激素释放激素、卵泡刺激素和黄体生成素的调控下主导调控龟类性腺发育、配子生成、交配、产卵及子代性别决定等重要生殖环节。孕酮、前列腺素、精氨酸催产素、皮质酮、甲状腺素和褪黑激素协同调节龟类生殖活动。在人工繁殖方面,使用GnRH等有助于克服龟类繁殖障碍,提高繁殖效率。对生殖激素对龟类生殖活动的调控作用与机理及其应用的研究进展进行了综述。
生殖激素;龟类;生殖调控
龟类是具有重要科研和经济价值的一个动物类群。长期以来,由于对野生资源的过度开发利用和栖息地的破坏,目前全球约50%的龟类濒临灭绝。人工繁殖是保护濒危龟类和实现其经济价值的重要手段,但是目前仍有许多龟类的人工驯养繁殖没有成功[1]。龟类的生殖活动受生殖激素的调控,在人工饲养条件下部分种类因为环境不适而导致内分泌失调,出现繁殖障碍,而利用外源激素则能有效调控繁殖活动[2-3]。生殖激素对龟类生殖调控的研究长期以来受到动物学家的广泛重视,迄今为止已经在《GeneralandComparativeEndocrinology》、《JournalofEndocrinology》、《Endocrinology》、《BiologyofReproduction》等期刊发表了110多篇有关研究论文,其中我国学者发表有关论文约15篇。笔者对龟类生殖激素的调控作用与机理及其应用情况进行了综述,以期为我国龟类生殖调控研究提供参考。
1 龟类生殖激素的种类、调控作用与机理
生殖激素是指与生殖活动有直接或间接关系的激素。生殖激素根据其来源可分为下丘脑激素、垂体激素、性腺激素等[4];根据首要作用模式,可分为神经激素和性腺激素等[5];根据激素结构,可分为蛋白类激素、甾体类激素等[5]。目前,已知调节龟类生殖活动的生殖激素有11种。促性腺激素释放激素(Gonadotropin-releasinghormone,GnRH)是由下丘脑分泌的一种神经多肽,能调节垂体中促性腺激素的合成与分泌,从而控制性腺激素的合成[6]。促性腺激素包括卵泡刺激素(Follicule-stimulatinghormone,FSH)和黄体生成素(Luteinizinghormone,LH),二者均是腺垂体嗜碱性细胞分泌的蛋白类激素。性腺分泌的激素主要包括雌二醇(Estradiol,E2)、睾酮(Testosterone,T)和孕酮(Progesterone,P),三者均为甾体类激素,分别主要来自于卵巢颗粒细胞、睾丸间质细胞和黄体。此外,皮质酮(Corticosterone,B)[7]、前列腺素(Prostaglandin,PG)[8]、精氨酸催产素(Argininevasotocin,AVT)[9]、甲状腺素(Thyroxine,T4)[10]和褪黑激素(Melatonin,MLT)[11]均参与了龟类的生殖调控过程。龟类生殖周期主要包括性腺的发育成熟、配子生成、交配、产卵及孵化5个重要环节。11种生殖激素的来源、化学特性及作用模式见表1。
表1龟类11种生殖激素的来源、化学特性及作用模式
Table1Source,chemicalcharacteristicandfunctiontypeof11reproductivehormonesinturtles

名称Name简称Abbreviation主要来源Mainsources化学特性Chemicalcharacteristics作用模式Actionmode促性腺激素释放激素GonadotropinreleasinghormoneGnRH下丘脑多肽神经卵泡刺激素FolliclestimulatinghormoneFSH垂体糖蛋白性腺黄体生成素LuteinizinghormoneLH垂体糖蛋白性腺雌二醇EstradiolE2卵巢类固醇性腺睾酮TestosteroneT睾丸类固醇性腺孕酮ProgesteroneP黄体类固醇性腺皮质酮CorticosteroneB肾上腺甾体类其他前列腺素ProstaglandinPG广泛分布脂肪酸其他精氨酸催产素ArgininevasotocinAVT垂体多肽神经甲状腺素ThyroxineT4甲状腺蛋白类其他褪黑激素MelatoninMLT松果体胺类神经
1.1性腺及附属性腺的发育和成熟性腺的生长发育直至成熟是龟类一切生殖活动发生的基础。GnRH、FSH、LH、E2和T通过下丘脑-垂体-性腺(Hypothalamic-pituitary-gonad,HPG)轴参与龟类性腺发育调控,其中E2和T起主导作用(图1)。对四眼斑水龟(Sacalia quadriocellata)的研究表明,GnRH可以促进睾丸发育,能使睾丸重量显著增加[12]。FSH和LH同样能够促进龟类睾丸生长及附睾的发育[13]。蛇鳄龟(Chelydra serpentina)中E2和T浓度升高分别能够促使雌性卵巢卵泡的生长和雄性睾丸的成熟[10,14]。此外,E2和T还能够调控附属性腺(输卵管、附睾等)的功能。例如,E2能够协同P调节生殖管道的运动性[15],附睾中高含量的T有助于维持附睾管细胞的形态及功能[16]。
1.2配子生成性腺的生长发育往往伴随着配子的生成,因此E2和T也是龟类卵子和精子生成调控的关键激素,其作用同样由HPG轴调控。龟类中GnRH对配子的生成是必需的,缺少GnRH会导致滞育现象[17]。研究表明,多种龟类中E2浓度升高能够促进卵巢卵泡生长[10],但对精子生成的作用尚不明确,尽管E2受体在雄龟精原细胞和曲细精管中有表达[18]。T调控精子生成与成熟,雄性四眼斑水龟血清中T含量在夏季升高,与精子的发生和成熟同步[19]。在陆龟中T水平在精子生成时期达到峰值,同时研究还表明附睾中高含量的T有助于精子的成熟[16],但T对卵子生成的作用研究尚未见报道。在雌龟排卵过程中,除了E2外,P是能够促进排卵的另一个主要激素。在加拉帕戈斯象龟(Geochelone nigra)和锦龟(Chrysemys picta)的研究中均证实P浓度升高可以促进排卵[14,20]。
FSH能够促进卵子精子生长并刺激排卵[21],而LH仅对精子生成有促进作用[13],这可能是因为性腺激素抑制了LH的调节作用。在龟类中,E2对LH有直接的抑制作用[22],而T则能够通过转化为E2间接发挥抑制作用[23]。
1.3交配行为T对龟类的交配行为有重要的影响。在雄龟中,T是影响雄性生殖行为的主导激素。对海龟的研究表明T峰值出现在交配期[7]。多种陆龟中,雄龟求偶和交配行为随着T浓度的升高而显著增加[20]。在四眼斑水龟中GnRH也被证实能够促进雄性求偶行为[24],据此推测GnRH通过HPG轴上调T水平发挥的作用。T不仅能够增加雄龟的求偶交配行为,还能够引发雌龟的交配行为。在雌性海龟中,交配接受能力与T水平呈正相关,高水平的T促进其接受雄性爬跨行为[25]。研究表明,雌龟能够产生吸引雄性的信息素,T可能促进信息素的释放,从而加强雌龟的交配行为[26]。
1.4产卵龟卵在输卵管中与精子结合受精,再由卵壳腺分泌富含钙质的蛋壳包裹,形成具有白色蛋壳的龟蛋。迄今为止,对这一过程的激素调控机理尚未见报道。雌性绿海龟(Chelonia mydas)体内E2和T含量在产卵期均下降到较低水平[27],据此推测E2和T对龟卵发育及产卵无显著作用。大量研究表明,龟卵从输卵管发育至产卵过程中,P含量显著升高,而且一年中P浓度会随着雌龟多次产卵出现多次峰值[20]。对绿海龟的研究还表明P能够增加龟卵在子宫中的保留时间[27]。这些研究表明P可能促进龟卵的发育,但抑制龟卵的排出。此外,PG能够通过刺激子宫内膜收缩和黄体溶解促进产卵[8],而AVT则通过刺激PG释放来加强其促产卵作用[9]。
1.5子代性别决定温度依赖性别决定模式(Temperature-dependentsexdetermination,TSD)被认为是龟类子代性别决定最普遍、最典型的方式[28]。然而,近些年越来越多研究表明生殖激素能够影响TSD中子代的性别。对红耳滑龟(Trachemys scriptaelegans)在孵化温度敏感期用外源E2处理导致本该是雄性的子代体内形成卵巢,而抑制E2作用后则导致睾丸的形成[29]。类似的研究报道表明高水平的T也会导致雄性后代的产生[30]。研究表明,孵化温度能够影响孵化过程中E2合成关键酶芳香化酶的合成,从而影响子代体内E2的合成以及卵巢或睾丸的生成,进而决定子代的性别[29]。龟类体外试验表明,在温度敏感期用芳香化酶抑制剂处理龟蛋,即使在雌性孵化温度下子代依然是雄性[31]。因此,龟类TSD是通过体内E2和T的相对含量来发挥作用的。
综上所述,在GnRH、FSH和LH的调控下,E2和T主导调控龟类性腺发育、配子生成、雌雄交配及子代性别决定等主要生殖过程,其中P调节排卵过程,PG和AVT促进产卵,如图1所示。虽然龟类激素的调控研究取得了一定的进展,但其主要生殖过程中生殖激素的调控机理尚不确定,例如T4和MLT,特别是交配和产卵过程中的生殖激素调控机理还需要更深入的研究。
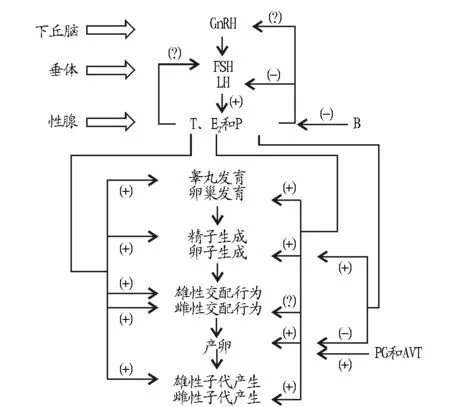
注:GnRH.促性腺激素释放激素;FSH.卵泡刺激素;LH.黄体生成素;E2.雌二醇;T.睾酮;P.孕酮;B.皮质酮,由肾上腺分泌;PG.前列腺素;AVT.精氨酸催产素;(+)表示促进;(-)表示抑制;(?)表示作用不确定;→表示调控; 表示分泌。Note:GnRH.Gonadotropin-releasing hormone;FSH,Follicule-stimulating hormone;LH,Luteinizing hormone;E2,Estradiol;T,Testosterone;P,Progesterone;B,Corticosterone,secreted by adrenal gland;PG,Prostaglandin;AVT,Arginine vasotocin;(+),Activation;(-),Inhibition;(?),Uncertain effect;→,Manipulation, ,Secretion.图1 龟类中生殖激素的生殖调控作用示意Fig.1 Schematic diagram of hormonal manipulation of turtle reproduction
2 生殖激素在龟类人工繁殖中的应用
人工繁殖是保护濒危龟类和实现其经济价值的重要手段。然而,目前在龟类人工繁殖过程中不交配、不产卵等繁殖障碍问题和繁殖效率低问题严重阻碍龟类人工繁殖的进程。龟类人工繁殖的相关研究大多集中在养殖环境的改善,近些年也有少量研究报道利用生殖激素处理来解决龟类人工繁育问题,并取得了一定的进展。
2.1利用生殖激素克服繁殖障碍大多数龟类在人工繁殖过程中都存在繁殖障碍问题,长时期的养殖压力能够激发龟的应激反应,引起B水平升高,从而干扰龟的繁殖活动[7]。生殖激素能够调节生殖生理及内分泌活动,促进龟类的交配繁殖。四眼斑水龟被注射GnRH类似物和人绒毛膜促性腺激素,同时降低B含量后不仅促进交配,同时促进产卵,成功实现了四眼斑水龟的人工繁殖[3]。开曼群岛的龟养殖中心于1977年向来自野外的雌性绿海龟注射人催产素成功使其连续数年产卵[7]。另外有研究表明,用人催产素处理使来自35种属的255只雌龟出现排卵现象[32]。因此,生殖激素能够克服龟类人工繁殖过程中不交配、不产卵等繁殖障碍,促进龟类繁殖。
2.2利用生殖激素提高繁殖效率自然状态下,欧洲陆龟(Testudo graeca terrestris)需要12~14年达到性成熟,但是利用具有抗雌激素作用的三苯氧胺处理后能够缩短雌龟到达性成熟的时间,使其提前进入成熟可育阶段,因此子代数量也有所增加[33]。在多种雌龟[如钻纹龟(Malaclemys terrapin)、卡罗莱纳箱龟(Terrapene carolina)和拟鳄龟]中,卵在受精后能保留4年才排出体外[34],利用生殖激素处理可能促使龟卵提早排出,提高繁殖效率。同时,研究表明不同种属龟类产卵数量存在较大差异,可能是因为体内生殖激素含量不同[35]。因此,利用生殖激素提高龟类产卵数量是人工繁育过程中提高龟类繁殖效率的潜在有效途径。
综上所述,生殖激素处理有助于解决龟类人工繁殖过程中遇到的繁殖障碍和繁殖效率低问题,虽然目前已经取得了一定的进展,但是生殖激素的应用技术尚不成熟,大部分生殖激素的应用未见研究报道,已研究的生殖激素应用技术尚未得到广泛推广,且其效果存在种间差异,易受养殖环境的影响,因此还需要进一步研究。
3 小结与展望
目前已发现有11种激素参与龟类生殖活动的调控。E2和T在GnRH、FSH和LH的调控下主导调控龟类的性腺发育、配子生成、交配、产卵及子代性别决定等重要生殖环节。其他6种激素协同调节龟类生殖活动:P调节排卵,PG和AVT促进产卵,B因参与机体的应激反应而抑制龟类的生殖活动,T4和MLT的作用尚不清楚。在龟类人工繁殖过程中,外源使用GnRH等多种生殖激素有助于克服龟类繁殖障碍,提高其繁殖效率。
尽管生殖激素对龟类生殖活动的调控在理论和实践方面都取得了一定的进展,但仍有诸多问题有待研究和解决。①E2受体在雄龟睾丸中表达,但其作用并不清楚。作为E2合成前体的T对卵子生成是否有作用尚无研究。②LH作用被E2和T抑制,因此LH促进精子生成的具体调控机理仍需要进一步研究。③E2是否调控雌龟交配行为尚需要进一步研究。④大量龟卵滞留未排出是由于卵发育不成熟还是产卵活动被抑制目前还不确定,而P是否能够促进卵在生殖管道中的发育以及P在产卵过程中的作用均亟待进一步研究。⑤主导调节激素E2和T在实践中尚未被研究,且已应用的生殖激素的作用效果仅在部分龟种生殖过程中得到验证,这些激素能否能普遍应用于各种龟类繁殖尚需进一步研究。⑥生殖激素的应用效果受养殖环境的严重制约,如何排除B对龟生殖活动的抑制值得进一步探讨。
针对以上龟类生殖激素调控研究中存在的问题,今后的研究工作可从以下方面开展:①进一步研究生殖激素对配子生成的调控机制,例如E2对精子生成及T对卵子发育的作用、性腺激素对GnRH和FSH的反馈作用等。②进一步探讨生殖激素对交配行为的影响,这将有助于解决龟人工繁殖过程中的不交配问题,例如E2对雌龟交配行为是否有影响、外源T处理对雌雄交配行为是否有促进作用。③研究生殖激素在促进龟产卵方面的应用,例如P对卵发育的作用、外源PG和AVT对产卵的促进作用等。
[1] 史海涛,洪关玲,傅丽容,等.龟类的养殖与保护[J].生物学通报,2009,44(1):18-21.
[2]CURRYLOWAF,TIFTMS,MEYERJL,etal.SeasonalvariationsinplasmavitellogeninandsexsteroidsinmaleandfemaleEasternBoxTurtles,Terrapene carolina carolina[J].Generalandcomparativeendocrinology,2013,180:48-55.
[3]HEB,LIUY,SHIH,etal.Captivebreedingofthefour-eyedturtle(Sacalia quadriocellata)[J].Asianherpetologicalresearch,2010,1(2):111-117.
[4]WUTTKEW,JARRYH,SEIDLOVA-WUTTKED.Definition,classificationandmechanismofactionofendocrinedisruptingchemicals[J].Hormones(Athens),2010,9(1):9-15.
[5]SCHATTENH,CONSTANTINESCUGM.Comparativereproductivebiology[M].Iowa:IowaStateUniversityPress,2007:118-120.
[6]KALRASP.Mandatoryneuropeptide-steroidsignalingforthepreovulatoryluteinizinghormone-releasinghormonedischarge[J].Endocrinereviews,1993,14(5):507-538.
[7]VALENTEAL,VELARDER,PARGAML,etal.Reproductivestatusofcaptiveloggerheadseaturtlesbasedonserumlevelsofgonadalsteroidhormones,corticosteroneandthyroxin[J].Veterinaryjournal,2011,187(2):255-259.
[8]GUILLETTELJJR,CREEA,GROSSTS.Endocrinologyofovipositioninthetuatara(Sphenodon punctatus):I.Plasmasteroidsandprostaglandinsduringnaturalnesting[J].Biologyofreproduction,1990,43(2):285-289.
[9]BUTLERDG,SNITMANFS.RenalresponsestomesotocininWesternpaintedturtlescomparedwithtantidiureticresponsetoargininevasotocin[J].Generalandcomparativeendocrinology,2005,144(2):101-109.
[10]LABRADA-MARTAGNV,MÉNDEZ-RODRGUEZLC,MANGELM,etal.Applyinggeneralizedlinearmodelsasanexplanatorytoolofsexsteroids,thyroidhormonesandtheirrelationshipswithenvironmentalandphysiologicfactorsinimmatureEastPacificgreenseaturtles(Chelonia mydas)[J].ComparativebiochemistryandphysiologypartA:Molecularandintegrativephysiology,2013,166(1):91-100.
[11] 李闯,汪继超,张杰,等.光周期对四眼斑水龟繁殖影响[J].动物学杂志,2013,48(2):193-199.
[12]DIAZPC,KELLERC,ANDREUAC.Clutchfrequency,eggandclutchcharacteristicsandnestingactivityofspur-thighedtortoises,Testudo graeca,insouthwesternSpain[J].CanadianJournalofZoology,1996,74:560-564.
[13]RAYPP,MAITIBR.Gonadotropinsandsexhormonesmodulateinterrenalfunctioninsoft-shelledturtle[J].Foliabiologica(Krakow),2002,50(3/4):115-120.
[14]KAKIZOEY,FUJIWARAM,AKUNEY,etal.Cyclicalchangesofplasmasexsteroidsincaptivebreedingloggerheadturtles(Caretta caretta)[J].Journalofzooandwildlifemedicine,2010,41(4):643-648.
[15]LORANC,NOVILLOA,CALLARDIP.Synergisticroleforpituitarygrowthhormoneintheregulationofhepaticestrogenandprogesteronereceptorsandvitellogenesisinfemalefreshwaterturtles,Chrysemys picta[J].Generalandcomparativeendocrinology,2005,140(1):25-32.
[16]SEREAUM,LAGARDEF,BONNETX,etal.DoestestosteroneinfluenceactivitybudgetinthemaleGreektortoise(Testudo graeca graeca)?[J].Generalandcomparativeendocrinology,2010,167(2):181-189.
[17]TSUIHW.Stimulationofandrogenproductionbythelizardtestis:SiteofactionofovineFSHandLH[J].Generalandcomparativeendocrinology,1976,28(4):386-394.
[18]GISTDH,BRADSHAWS,MORROWCM,etal.Estrogenresponsesysteminthereproductivetractofthemaleturtle:Animmunocytochemicalstudy[J].Generalandcomparativeendocrinology,2007,151(1):27-33.
[19] 傅丽容,贺斌,洪美玲,等.四眼斑水龟血浆生殖激素季节性变化[J].动物学杂志,2008,43(3):56-59.
[20]SCHRAMMBG,CASARESM,LANCEVA.SteroidlevelsandreproductivecycleoftheGalápagostortoise,Geochelone nigra,livingunderseminaturalconditionsonSantaCruzIsland(Galápagos)[J].Generalandcomparativeendocrinology,1999,114(1):108-120.
[21]CHIENJT,SHENST,LINYS,etal.MolecularcloningofthecDNAencodingfollicle-stimulatinghormonebetasubunitoftheChinesesoft-shellturtlePelodiscus sinensis,anditsgeneexpression[J].Generalandcomparativeendocrinology,2005,141(2):190-200.
[22]PAVGIS,LICHTP.Steroidalmodulationofpituitarygonadotropin-releasinghormoneresponsivenessinyoungturtles,Pseudemys scripta[J].Generalandcomparativeendocrinology,1990,78(3):331-343.
[23]TSAIPS,HAYESTB,LICHTP.Roleofaromatizationintestosterone-inducedinhibitionofluteinizinghormonesecretioninfemaleturtles,Trachemys scripta[J].Biologyofreproduction,1994,50(1):144-151.
[24]HEB,LIUY,SHIH,etal.Effectofluteinizinghormone-releasinghormoneanalogueonthesexualbehaviorofsacaliaquadriocellata[J].Asianherpetologicalresearch,2010,1(1):40-43.
[25]SELCERKW,SMITHS,CLEMENSJW,etal.Androgenreceptorintheoviductoftheturtle,Trachemys scripta[J].ComparativebiochemistryandphysiologypartB:Biochemistryandmolecularbiology,2005,141(1):61-70.
[26]MUOZA.Chemo-orientationusingconspecificchemicalcuesinthestripe-neckedterrapin(Mauremys leprosa)[J].Journalofchemicalecology,2004,30(3):519-530.
[27]AL-HABSIAA,ALKINDIAY,MAHMOUDIY,etal.PlasmahormonelevelsinthegreenturtlesChelonia mydasduringpeakperiodofnestingatRasAl-Hadd-Oman[J].Journalofendocrinology,2006,191(1):9-14.
[28] 黄敏毅,张育辉,王宏元.非哺乳类脊椎动物性别决定的研究进展[J].动物学杂志,2004,39(4):104-108.
[29]RAMSEYM,CREWSD.Steroidsignalingandtemperature-dependentsexdetermination-reviewingtheevidenceforearlyactionofestrogenduringovariandeterminationinturtles[J].Seminarsincellanddevelopmentalbiology,2009,20(3):283-292.
[30]BELAIDB,RICHARD-MERCIERN,PIEAUC,etal.SexreversalandaromataseintheEuropeanpondturtle:Treatmentwithletrozoleafterthethermosensitiveperiodforsexdetermination[J].Journalofexperimentalzoology,2001,290(5):490-497.
[31]NAKAMURAM.Themechanismofsexdeterminationinvertebrates-aresexsteroidsthekey-factor?[J].JournalofexperimentalzoologypartA:Ecologicalgeneticsandphysiology,2010,313(7):381-398.
[32]EWERTMA,LEGLERJM.Hormonalinductionofovipositioninturtles[J].Herpetologica,1978,34(3):314-318.
[33]LAPIDR,NIRI,SNAPIRN,etal.Reproductivetraitsinthespur-thighedtortoise(Testudo graeca terrestris):Newtoolsfortheenhancementofreproductivesuccessandsurvivorship[J].Theriogenology,2004,61(6):1147-1162.
[34]MOLLED.Reproductivecyclesandadaptations[M].NewYork:CornellUniversityPress,1979:305-331.
[35]RAFFERTYAR,EVANSRG,SCHEELINGSTF,etal.Limitedoxygenavailabilityinuteromayconstraintheevolutionoflivebirthinreptiles[J].Americannaturalist,2013,181(2):245-253.
ResearchProgressofHormonalManipulationinTurtleReproduction
LIXiao-nan,FANGQing*
(NorthSichuanMedicalCollege,Nanchong,Sichuan637100)
Todate, 11kindsofimportantreproductivehormoneshavebeenfoundinvolvedinthemanipulationofturtlereproduction.Estradiolandtestosteroneplaydominantregulatoryroleintheimportantactivitiesofgonadaldevelopment,gametegeneration,copulation,ovipositionandoffspringsexdeterminationundertheregulationofgonadotropin-releasinghormone,follicule-stimulatinghormoneandluteinizinghormone.Progesterone,prostaglandin,argininevasotocin,corticosterone,thyroxineandmelatoninsynergisticallyregulateturtlereproductiveactivities.Inturtleartificialbreedingresearches,usingGnRHandoxytocincontributestoovercomereproductiveobstaclesandimprovereproductiveefficiency.Theresearchadvancesofhormonalmanipulationinturtlereproductionwerereviewed.
Reproductivehormone;Turtle;Reproductivemanipulation
川北医学院2013年质量项目。
李晓楠(1987- ),女,河南南阳人,助教,硕士,从事动物繁育生理和动物生殖生理学研究。*通讯作者,助理研究员,硕士,从事实验动物管理工作。
2016-05-15
S966.5
A
0517-6611(2016)17-130-04
猜你喜欢
文萃报·周五版(2022年24期)2022-06-21
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
广西林业科学(2016年3期)2016-03-16
中国马铃薯(2015年3期)2015-12-19
食品工业科技(2014年15期)2014-03-11
微创泌尿外科杂志(2014年4期)2014-02-28
