
一氧化氮调控青菜根尖细胞壁过氧化物酶抗镉胁迫的研究
2016-08-10 02:58吕文静杨立飞
江西农业学报 2016年7期
吕文静,陈 健,杨立飞*
(1.南京农业大学 园艺学院,江苏 南京 210095;2.江苏省农业科学院 食品质量安全与检测研究所,江苏 南京 210014)
一氧化氮调控青菜根尖细胞壁过氧化物酶抗镉胁迫的研究
吕文静1,陈 健2,杨立飞1*
(1.南京农业大学 园艺学院,江苏 南京 210095;2.江苏省农业科学院 食品质量安全与检测研究所,江苏 南京 210014)
摘要:研究了镉胁迫下一氧化氮对青菜幼苗根尖伸长及其木质素生物合成的影响。结果显示:镉处理显著抑制根的生长,并呈现浓度和时间效应;镉处理诱导根尖细胞壁过氧化物酶(CPX) 活力的显著上升,并呈现浓度和时间效应;外源一氧化氮供体SNP能够显著缓解镉胁迫对根生长的抑制效应;镉处理显著降低了根尖内源一氧化氮水平,而添加SNP有效提升了镉胁迫下根尖内源一氧化氮水平;添加SNP能够显著缓解镉胁迫诱导的根尖CPX和NADH氧化酶活力的上升以及木质素的合成。这些结果说明一氧化氮可通过抑制青菜幼苗根尖CPX介导的木质素生物合成来抵抗镉胁迫。
关键词:镉胁迫;一氧化氮;细胞壁过氧化物酶;木质素;青菜
重金属镉(Cd)是植物体内的非必需元素,随着农业环境中镉污染日趋加重,过量镉严重影响了农作物的生长和品质[1]。镉胁迫能够通过引起植物体内多层面的生理生化响应,最终抑制植物的生长[2-4]。细胞的分化和伸长是决定植物生长的关键因素之一,而细胞壁作为控制细胞形状的骨架结构,其机械强度则决定细胞的分化和伸长[5]。植物细胞壁骨架主要由纤维素、木质素和多糖组成。Hafrén等[6]发现细胞壁的木质化程度显著影响糖类物质的分布和结构,从而影响细胞的伸长。已有研究显示,重金属(如铜、镉、铝等)胁迫能够诱导植物细胞壁中木质素的快速大量合成[7-9]。在细胞壁中存在大量功能性蛋白,其中的细胞壁过氧化物酶(CPX, cell wall peroxidase)是负责催化木质素合成途径最后一步的关键酶,而此催化步骤中所需的过氧化氢则来自NADH氧化酶(NADH oxidase)[10-11]。重金属胁迫诱导的木质素累积则会伴随着CPX和NADH氧化酶活性的升高[7-9]。虽然关于镉胁迫下植物体内信号转导调控已有大量研究[3],但在镉胁迫下木质素合成途径的信号调控方式尚不明晰。
一氧化氮(Nitric oxide, NO)是调控植物响应环境胁迫的重要气体信号分子[12]。根据其调控的下游途径和作用方式不同,一氧化氮既能保护植物抵御重金属胁迫[13],亦能介导重金属胁迫对植物造成的毒害效应[14-15]。有研究表明,一氧化氮与植物根内木质素的合成密切相关[16-17]。然而在重金属胁迫下,一氧化氮是否并怎样调控木质素合成还有待进一步确认。本文以青菜幼苗为材料,研究了镉胁迫下一氧化氮对根的生长和对木质素合成的调控方式,以期为镉胁迫下植物体内生理代谢的调控网络提供新的理论补充。
1材料与方法
1.1试验材料
供试小白菜品种为南京绿领种业生产的五月慢。内源NO荧光染料DAF-DA和外源NO供体SNP购自海门碧云天生物科技有限公司;生理生化测定相关试剂和试剂盒均购自南京建成生物工程研究所;其余常规化学试剂均购自国药集团。
1.2试验设计
小白菜种子经过1%NaClO消毒10 min后,用蒸馏水冲洗3次,在25 ℃条件下避光催芽12 h,然后转入1/4 Hoagland营养液,在光照培养箱(RDN-300B-3,宁波东南)中培养,培养条件为25 ℃/22 ℃(昼/夜),光照周期为14 h/10 h(光/暗),光照强度为200 μmol/(m2·s)。培养2 d后(根长约为3 cm),挑选健壮、长势均一的幼苗,测定初始根长并将其转移到处理液中。设置营养液中CdCl2最终浓度分别为0、20、40、80、120 μmol/L;在处理24 h后,检测根长和根尖CPX活性。对经40 μmol/L CdCl2处理的幼苗,检测其根长和根尖CPX活性在24 h内的变化趋势。为了探讨外源NO对Cd胁迫下根生长的影响,设置以下处理: CK、Cd(40 μmol/L)、Cd(40 μmol/L)+SNP(0.05 mmol/L)、Cd(40 μmol/L)+SNP(0.1 mmol/L)、Cd(40 μmol/L)+SNP(0.2 mmol/L)、Cd(40 μmol/L)+SNP(0.4 mmol/L)、Cd(40 μmol/L)+SNP(0.8 mmol/L);在处理24 h后测定根长。为了进一步探讨NO信号在本研究中的作用,设置以下处理: CK、Cd (40 μmol/L)、Cd(40 μmol/L)+SNP(0.2 mmol/L)、SNP(0.2 mmol/L);在处理24 h后,测定根长、根尖内相关酶活性、根尖内源NO荧光。对处理后的幼苗取根尖1 cm左右,保存于液氮中,用于生化指标的检测。
1.3测定项目与方法
1.3.1根尖中内源NO组织荧光检测与分析对根内NO采用特异性荧光探针DAF-FMDA进行原位检测[18]。将经不同处理的青菜幼苗根部用蒸馏水冲洗3次,再吸干表面水分,将根部浸入到15 μmol/L DAF-FMDA探针溶液中,在25 ℃、避光条件下装载15 min。探针装载完后用蒸馏水冲洗3次,在荧光显微镜(ECLIPSE, TE2000-S, Nikon)下以490 nm为激发波长、525 nm为发射波长,并结合使用Image-Pro Plus 6.0软件统计根尖中DAF-DA荧光密度,以此来反映NO的相对含量。
1.3.2根尖中CPX活力的测定对根尖细胞壁结合蛋白的提取参照Lee等的方法[19]。切取1 cm根尖(1 g),在10 mL磷酸缓冲液(10 mmol/L, pH 6.4)中冰浴研磨。离心10 min(4 ℃,2000 r/min),弃上清。对沉淀用10 mL上述磷酸缓冲液重新悬浮洗涤,再次离心10 min(4 ℃,2000 r/min)。重复此悬浮、洗涤、离心步骤6次后,对沉淀采用NaCl(1 mol/L)悬浮,置于摇床悬摇2 h (30 ℃,130 r/min)。最后离心20 min(4 ℃, 12000 r/min),取上清立即进行酶活测定。
采用愈创木酚还原法[20]测定CPX的活性。反应体系包括:2.0 mL 100 mmol/L醋酸钠(pH 5.4)、1 mL 0.25%(w/v)愈创木酚、10~20L酶提取液和0.1 mL 0.75%H2O2。用间隔读数法记录420 nm处的吸光度值。消光系数为26.6/(mmol/L·cm)。一个酶活力单位定义为在37 ℃条件下,每毫克组织蛋白每分钟催化底物生成1 μmol产物的酶量。
1.3.3根尖中CPX活性的电泳分析采用1.3.2中的方法获得细胞壁蛋白溶液,进行同工酶电泳分析。采用非变性不连续聚丙烯酰胺凝胶电泳(PAGE),不加十二烷基硫酸钠。浓缩胶浓度为5%,分离胶浓度为10%。电泳在4 ℃下进行,电压在浓缩胶阶段为80 V,在分离胶阶段为120 V。电泳结束后下胶,置于含有0.68 mmol/L联苯胺和5 mmol/L H2O2的0.2 mol/L醋酸缓冲液 (pH 5.4)中染色20 min,直至红棕色条带清晰地出现,用去离子水清洗凝胶,终止反应并拍照[21]。
1.3.4根尖中NADH氧化酶活力的测定采用1.3.2中的方法获得细胞壁蛋白溶液,参照Ishida等[22]的方法测定NADH氧化酶的活力。反应体系中含有50mol/L NADH、30 mmol/L醋酸钠缓冲液(pH 6.5)、5 mmol/L MnCl2、20mol/L香豆酸及10~20L酶液。用间隔读数法记录340 nm处的吸光度值。消光系数为 6.22/(mmol/L·cm)。一个酶活力单位定义为每毫克蛋白每分钟氧化1mol NADH吸光值的变化量。
1.3.5蛋白质含量的测定对酶液中的蛋白质含量测定采用Bradford方法(考马斯亮蓝G250染色法),以牛血清白蛋白作为标准蛋白[23]。
1.3.6根尖细胞壁木质素染色参照Siegel[24]的方法。切取60条根的1 cm根尖部分,在10 mL磷酸缓冲液(10 mmol/L, pH 6.4)中冰浴研磨。离心10 min(4 ℃,2000 r/min),弃上清。对沉淀用10 mL上述磷酸缓冲液重新悬浮洗涤,再次离心10 min(4 ℃,2000 r/min)。重复此悬浮、洗涤、离心步骤3次后,弃上清,在沉淀中加入300 μL 1%(w/v)的间苯三酚[溶于20%(v/v)的HCl]显色并拍照。
1.3.7数据统计分析对每个结果采用3次重复的平均值±标准差表示,首先采用SPSS 2.0对试验数据进行单因素方差分析(ANOVA),然后进行F检验,检测不同处理之间在0.05水平下的差异显著性。采用最小显著差数法(LSD)对不同处理间在0.05水平下的差异显著性进行分析和多重比较。
2结果与分析
2.1镉处理对青菜幼苗根生长的影响
CdCl2(20~120 μmol/L)处理显著抑制青菜幼苗根的生长,并呈现浓度和时间效应。20、40、80、120 μmol/L的CdCl2处理24 h后,根的伸长量分别比对照下降了41.2%、53.1%、62.1%、78.2% (P<0.05) (图1A)。在时间进程试验中,我们每隔3 h监测了40 μmol/L的CdCl2处理后的根伸长量。结果显示:40 μmol/L的CdCl2处理6 h后根的伸长量开始显著低于对照;随着处理时间的延长,镉对根的抑制效果愈加明显(P<0.05) (图1B)。
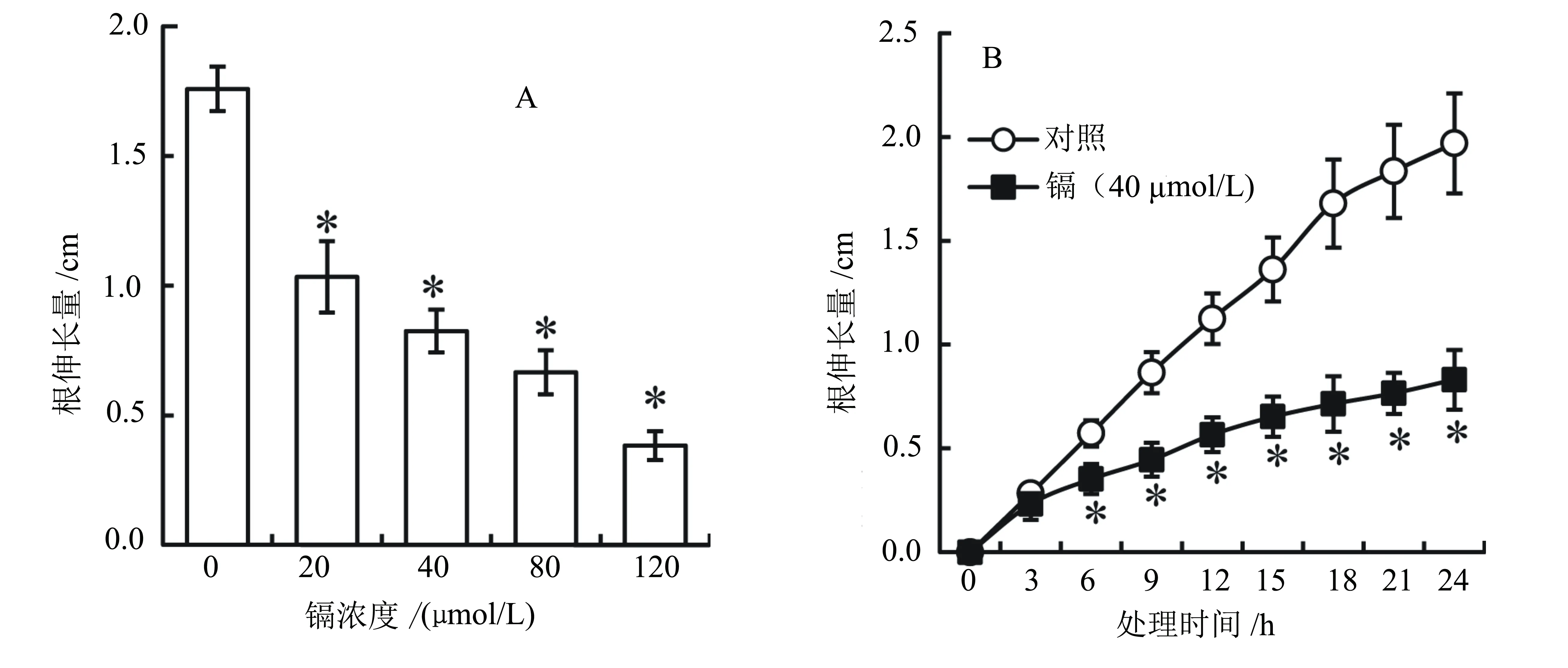
“*”表示在5%水平下与对照差异显著。下同。图1 镉处理对青菜幼苗根伸长量的影响
2.2镉处理对青菜幼苗根尖CPX活力的影响
根尖是植物根系伸长生长的关键部位[25]。酶活测定结果显示:青菜幼苗根尖中CPX活力随着镉处理浓度的升高而逐渐上升,其中20、40、80 μmol/L的CdCl2处理的根尖中CPX活力分别比对照升高了37.5%、69.4%、84.8%(P<0.05)(图2A);120 μmol/L的CdCl2处理的根尖中CPX活力升高趋势减缓,但仍比对照增加了57.4%(P<0.05) (图2A)。酶活时间动态监测结果显示,40 μmol/L的CdCl2处理3 h后根尖CPX活力开始显著高于对照的(P<0.05)(图2B),并随着处理时间的延长而逐渐升高。
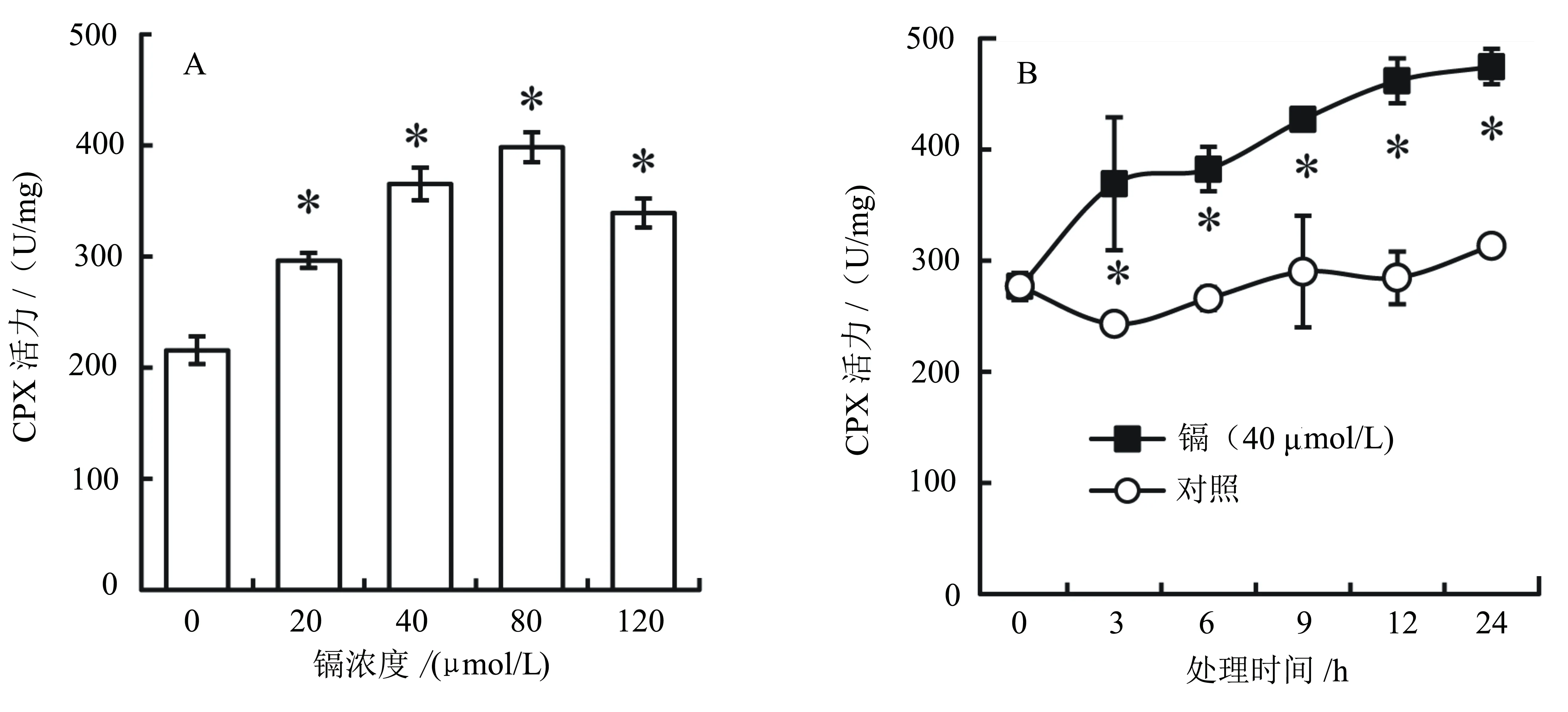
图2 镉处理对青菜幼苗根尖CPX活力的影响
2.3SNP处理对镉胁迫下青菜幼苗根生长的影响
为了研究外源补充一氧化氮对镉胁迫下青菜幼苗根生长的影响,在40 μmol/L的CdCl2处理的同时加入不同浓度的SNP。结果显示:在镉胁迫下,0.1~0.4 mmol/L的SNP能够显著促进根的伸长,其中以0.2 mmol/L SNP最为明显。经SNP(0.2 mmol/L)+Cd(40 μmol/L)组合处理的根伸长量比单独Cd处理增加了30.3%(P<0.05) (图3)。这说明由外源SNP提供的NO能够显著缓解镉胁迫对根的抑制效应。在后续实验中我们采用了0.2 mmol/L这一SNP最佳浓度。
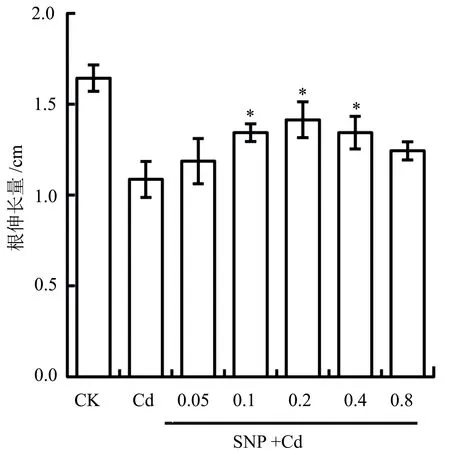
在SNP+Cd组合处理中,SNP的浓度为0.05~0.80 mmol/L,Cd的浓度为40 μmol/L。图3 SNP处理对镉胁迫下青菜幼苗根伸长量的影响
2.4镉处理对青菜幼苗根尖内源一氧化氮含量的影响
DAF-FM DA能够与植物细胞内的一氧化氮反应后发出绿色荧光,可对植物细胞内源一氧化氮进行原位检测。单独SNP处理后的根尖一氧化氮荧光强度较对照显著增强,说明SNP能够作为外源一氧化氮供体提升青菜幼苗根尖内源一氧化氮水平(图4A)。40 μmol/L的CdCl2处理3 h后根尖一氧化氮荧光强度较对照明显下降,而SNP+Cd组合处理较单独Cd处理能够提升根尖一氧化氮水平(图4A)。DAF-FM荧光密度可反映细胞内源一氧化氮的相对含量。统计结果显示:40 μmol/L的CdCl2处理3 h后根尖DAF-FM荧光密度较对照显著下降了44.3% (图4B);而SNP+Cd组合处理后根尖DAF-FM荧光密度则可恢复到对照水平(图4B)。这些结果说明镉胁迫能够抑制根尖内源一氧化氮的产生,而外源添加SNP则可以使镉胁迫下根尖内源一氧化氮恢复到对照水平。
2.5SNP处理对镉胁迫下青菜幼苗根尖CPX活力的影响
酶活测定结果显示,40 μmol/L CdCl2处理的根尖CPX活力较对照显著上升,而加入SNP后CPX活力则下降到对照水平。SNP(0.2 mmol/L)+Cd(40 μmol/L)组合处理的根尖CPX活力较单独Cd处理下降了28.6%(P<0.05)(图5A)。进一步的CPX同工酶活性电泳检测亦显示了相似的结果。本研究发现青菜幼苗根尖CPX有3条棕色活性泳带(Ⅰ、Ⅱ、Ⅲ),即为3个同工酶,这3个同工酶的活性在单独Cd处理后均较对照大幅上升,而加入SNP后则都显著下降(图5B)。单独SNP处理对根尖CPX的总活力和同工酶活性均无显著影响(图5B)。
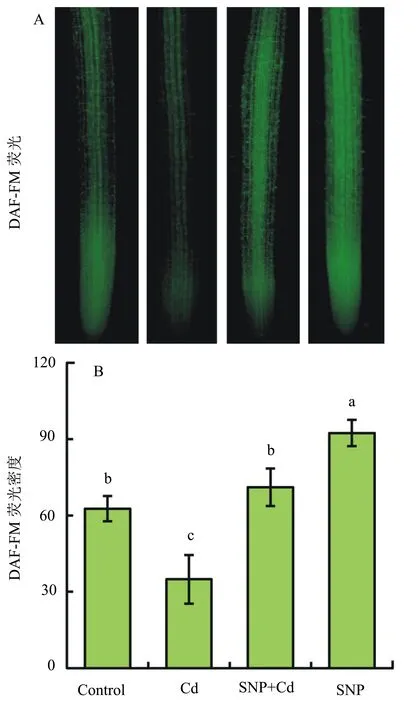
图4 镉处理对青菜幼苗根尖一氧化氮含量的影响
2.6SNP处理对镉胁迫下青菜幼苗根尖NADH氧化酶活力和木质素含量的影响
由于CPX是在NADH氧化酶的辅助下催化木质素的合成的,所以我们亦检测了根尖NADH氧化酶活力的变化。与对照相比较,青菜幼苗根尖NADH氧化酶活力在单独Cd(40 μmol/L)处理后大幅上升,而加入SNP后则又显著下降。单独SNP处理对根尖NADH氧化酶活力无显著影响(图6A)。木质素可与间苯三酚反应呈现棕色,根尖木质素的染色结果显示:与单独Cd处理的深棕色相比较,SNP+Cd共同处理的颜色明显变浅。这说明添加SNP能够显著降低Cd胁迫下根尖木质素的含量(图6B)。
3结论与讨论
本文的试验数据证明:镉胁迫通过刺激青菜幼苗根尖一氧化氮调控的木质素生物合成,增强细胞壁的刚性,导致根的伸长生长受阻。在本研究中,镉胁迫显著抑制根的伸长(图1),此过程伴随着木质素生物合成相关酶(CPX和NADH氧化酶)活性的显著升高(图2、图5、图6A)。Chaoui等[26]的研究发现镉和铜胁迫能够提高豌豆根细胞壁中以离子态结合的过氧化物酶的活性,而非共价态结合的过氧化物酶的活性。本研究从青菜根尖中通过高浓度NaCl分离得到的CPX即为离子态结合的过氧化物酶,说明CPX在响应重金属胁迫方面可以发挥重要作用。这可能与其通过作用于木质素合成调控细胞壁刚性密切相关[11]。另外,在Chaoui等[26]的研究中,20~100 μmol/L的镉处理对豌豆整个根部NADH氧化酶的活性无影响。而本研究发现40 μmol/L的镉处理能够大幅提高青菜幼苗根尖NADH氧化酶的活性(图6A)。这说明位于根尖部位的NADH氧化酶在调控镉胁迫下的细胞壁强度方面可能发挥更为关键的作用,这与根尖细胞的分化和伸长决定根的生长相对应[25]。
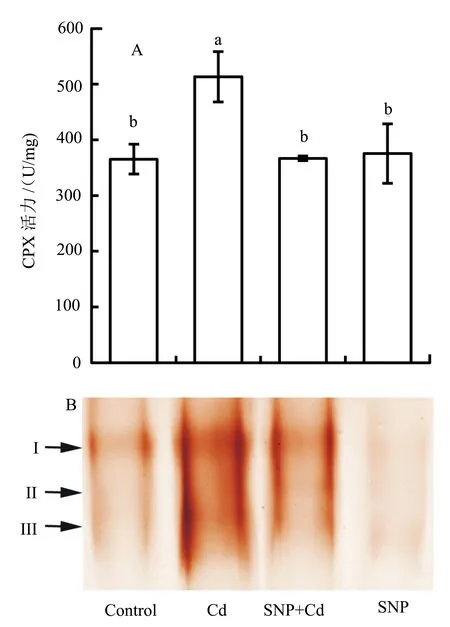
图5 SNP处理对镉胁迫下青菜幼苗根尖CPX活力和同工酶活性的影响
一氧化氮作为多功能信号分子能够全面调控植物对镉胁迫的生理响应[27]。在本研究中,镉胁迫显著降低根尖内源一氧化氮水平,而采用一氧化氮供体SNP处理则能提升内源一氧化氮水平(图4),并显著缓解镉胁迫对根生长的毒害效应(图3)。已有研究表明,降低向日葵根中内源一氧化氮水平后,包括CPX在内的木质素合成途径相关基因的表达量大幅上调[16]。这说明一氧化氮能够负调控木质素的合成。在本研究中,SNP处理能够显著抑制镉胁迫诱导的CPX活性、NADH氧化酶活性、木质素合成(图5、图6)。这说明SNP提供的一氧化氮通过抑制木质素的合成来减弱细胞壁强度,从而缓解了镉胁迫对根的抑制效应。另外有报道显示,铝胁迫下一氧化氮与茉莉酸信号的互作可调控植物根内CPX的活性和木质素的合成[20]。最新的研究显示,提升内源茉莉酸水平能够显著缓解镉胁迫对植物的毒害效应[28]。那么结合本研究结果,在镉胁迫下,一氧化氮与茉莉酸信号能否互作并调控木质素的生物合成?这种可能的调控是否对根的伸长生长有所贡献?通过进一步的研究回答上述问题,将有助于阐明镉胁迫下一氧化氮调控木质素合成的信号网络,从而为镉污染农业环境中作物的安全生产提供理论依据。
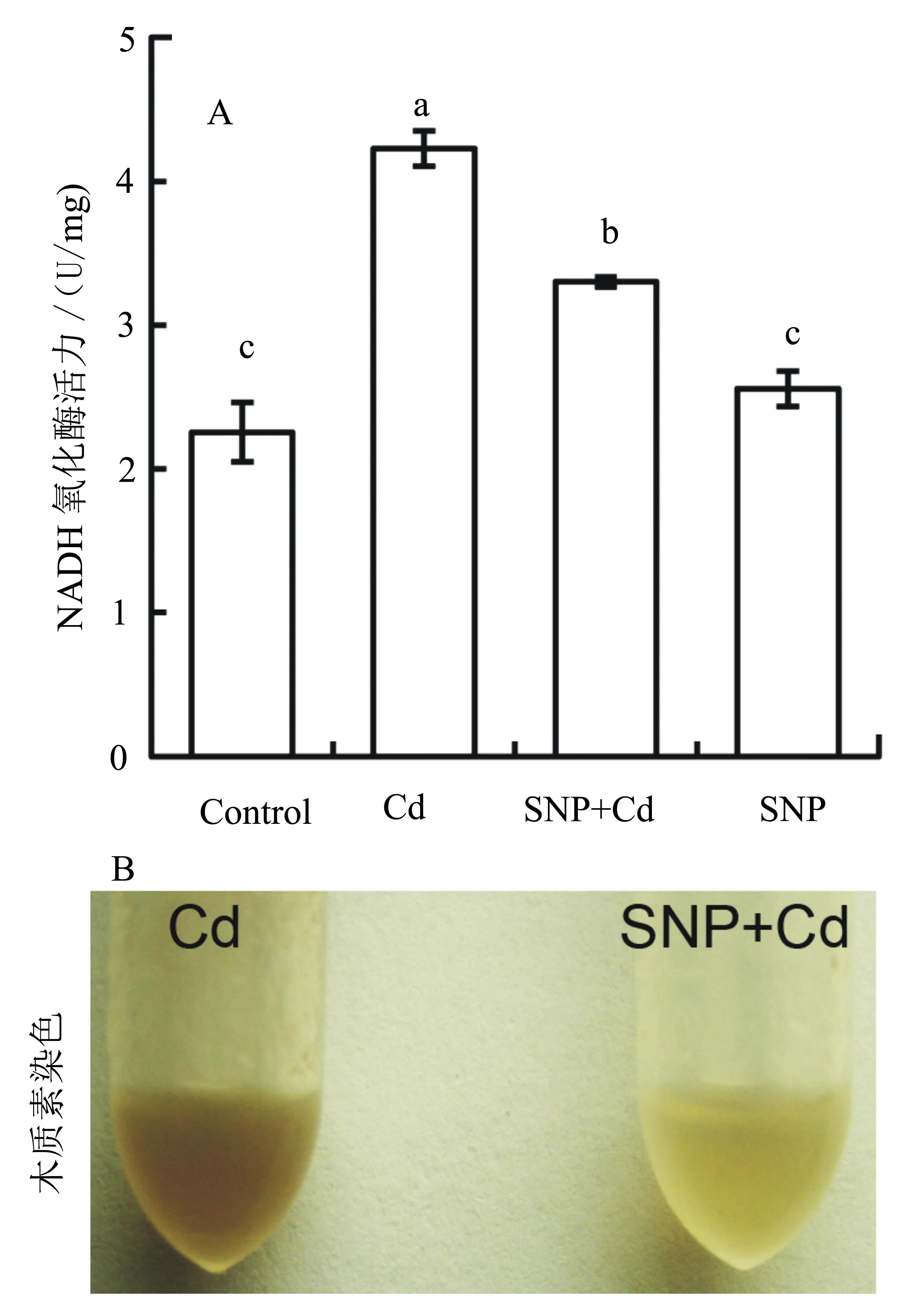
图6 SNP处理对镉胁迫下青菜幼苗根尖NADH氧化酶活力和木质素含量的影响
参考文献:
[1] Satarug S, Garrett S H, Sens M A, et al. Cadmium, environmental exposure, and health outcomes[J]. Ciência & Saúde Coletiva, 2011, 16(5): 2587-2602.
[2] Lin Y F, Aaers M. The molecular mechanism of zinc and cadmium stress response in plants[J]. Cellular and Molecular Life Science, 2012, 69(19): 3187-3206.
[3] DalCorso G, Farinati S, Furin A. Regulatory networks of cadmium stress in plants[J]. Plant Signaling & Behavior, 2010, 5(6): 663-667.
[4] DalCorso G, Farinati S, Maistri S, et al. How plants cope with cadmium: staking all on metabolism and gene expression[J]. Journal of Integrative Plant Biology, 2008, 50(10): 1268-1280.
[5] Cosgrove D J. Growth of the plant cell wall[J]. Nature Reviews Molecular Cell Biology, 2005, 6(11): 850-861.
[6] Hafrén J, Fujino T, Itoh T. Changes in cell wall architecture of differentiating tracheids ofPinusthunbergiiduring lignification[J]. Plant and Cell Physiology, 1999, 40(5): 532-541.
[7] Lin C C, Chen L M, Liu Z H. Rapid effect of copper on lignin biosynthesis in soybean roots[J]. Plant Science, 2005,168(3): 855-861.
[8] Yang Y J, Cheng L M, Liu Z H. Rapid effect of cadmium on lignin biosynthesis in soybean roots[J]. Plant Science, 2007, 172(3): 632-639.
[9] Xue Y J, Tao L, Yang Z M. Aluminum-induced cell wall peroxidase activity and lignin synthesis are differentially regulated by jasmonate and nitric oxide[J]. Journal of Agricultural and Food Chemistry, 2008, 56(20): 9676-9684.
[10] Barceló A R. Hydrogen peroxide production is a general property of the lignifying xylem from vascular plants[J]. Annals of Botany, 1998, 82(1): 97-103.
[11] Grabber, John H. How do lignin composition, structure, and cross-linking affect degradability? A review of cell wall model studies[J]. Crop Science, 2005, 45(3): 820-831.
[12] Besson B A, Pugin A, Wendehenne D. New insights into nitric oxide signaling in plants[J]. Annual Review of Plant Biology, 2008, 59(1): 21-39.
[13] Gallego S M, Pena L B, Barcia R A, et al. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms[J]. Environmental and Experimental Botany, 2012, 83(5): 33-46.
[14] Besson-Bard A, Gravot A, Richaud P, et al. Nitric oxide contributes to cadmium toxicity inArabidopsisby promoting cadmium accumulation in roots and by up-regulating genes related to iron uptake[J]. Plant Physiology, 2009, 149: 1302-1315.
[15] Chen M, Cui W, Zhu K, et al. Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production[J]. Journal of Hazardous Materials, 2014, 267: 40-47.
[16] Corti M G, Pinedo M, Di R J, et al. Nitric oxide is required for determining root architecture and lignin composition in sunflower. Supporting evidence from microarray analyses[J]. Nitric Oxide, 2014, 39: 20-28.
[17] Böhm F, Ferrarese M, Zanardo D L, et al. Nitric oxide affecting root growth, lignification and related enzymes in soybean seedlings[J]. Acta Physiologiae Plantarum, 2010, 32(6): 1039-1046.
[18] Guo K, Xia K. Regulation of tomato lateral root development by carbon monoxide and involvement in auxin and nitric oxide[J]. Journal of Experimental Botany, 2008, 59(12): 3443-3452.
[19] Lee T M, Lin Y H. Changes in soluble and cell wall-bound peroxidase activities with growth in anoxia-treated rice (OryzasativaL.) coleoptiles and roots[J]. Plant Science, 1995, 106(1): 1-7.
[20] Xue Y F, Liu L, Liu Z P, et al. Protective role of Ca against NaCl toxicity inJerusalemartichokeby up-regulation of antioxidant enzymes[J]. Pedosphere, 2008, 18(6): 766-774.
[21] Zhou Z S, Wang S J, Yang Z M. Biological detection and analysis of mercury toxicity to alfalfa (Medicagosativa) plants[J]. Chemosphere, 2008, 70(8): 1500-1509.
[22] Ishida A, Okubo K, Ono K. Formation of hydrogen peroxide by NAD(P)H oxidation with isolated cell wall-associated peroxidase from cultured liverwort cells,MarchantiapolymorphaL.[J]. Plant and Cell Physiology, 1987, 28(4): 723-726.
[23] Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry, 1976, 72: 248-254.
[24] Siegel S M. On the biosynthesis of lignin[J]. Physiologia Plantarum, 2006, 6(1): 134-139.
[25] Dupuy L, Gregory P J, Bengough A G. Root growth models: towards a new generation of continuous approaches[J]. Journal of Experimental Botany, 2010, 61(8): 2131-2143.
[26] Chaoui A, Jarrar B, Ferjani E E. Effects of cadmium and copper on peroxidase, NADH oxidase and IAA oxidase activities in cell wall, soluble and microsomal membrane fractions of pea roots[J]. Journal of Plant Physiology, 2004, 161(11): 1225-1234.
[27] Arasimowicz J M, Floryszak W J, Gwózdz E A. The message of nitric oxide in cadmium challenged plants[J]. Plant Science, 2011, 181(5): 612-620.
[28] Yan Z, Zhang W, Chen J, et al. Methyl jasmonate alleviates cadmium toxicity inSolanumnigrumby regulating metal uptake and antioxidative capacity[J]. Biologia Plantarum, 2015, 59(2): 373-381.
(责任编辑:黄荣华)
收稿日期:2016-01-09
基金项目:江苏省农业科技自主创新资金[CX(14)2096];国家自然科学基金项目(31101537)。
作者简介:吕文静(1988─),女,硕士研究生,主要从事蔬菜栽培生理与生物技术研究。*通讯作者:杨立飞。
中图分类号:S634.3
文献标志码:A
文章编号:1001-8581(2016)07-0011-06
Study on Cadmium-resistance ofBrassicarapathrough Regulating Cell Wall-bound Peroxidase in Its Root Tip by Nitric Oxide
LV Wen-jing1, CHEN Jian2, YANG Li-fei1*
(1. College of Horticulture, Nanjing Agricultural University, Nanjing 210095, China;2. Institute of Food Quality Safety and Testing, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China)
Abstract:This paper studied the influences of nitric oxide (NO) on the root tip elongation and lignin biosynthesis of Brassica rapa seedlings under cadmium (Cd) stress. The results indicated that: Cd treatment induced a remarkable inhibition to the root growth in both dose-dependent and time-dependent manners; Cd treatment resulted in a significant increase in the activity of cell wall-bound peroxidase (CPX) in root tip in both dose-dependent and time-dependent manners; the exogenous NO donor SNP significantly attenuated the Cd-induced inhibition to root elongation; Cd treatment significantly reduced the content of endogenous NO in root tip, while the addition of SNP effectively enhanced the content of endogenous NO in Cd-treated root tip; the addition of SNP significantly decreased the content of lignin and the activities of CPX and NADH oxidase in root tip under Cd stress. These results suggested that NO played an important role in the regulation of Cd resistance by inhibiting CPX-mediated lignin biosynthesis in the root tip of B. rapa seedlings.
Key words:Cadmium stress; Nitric oxide; Cell wall-bound peroxidase; Lignin; Brassica rapa
猜你喜欢
上海农业科技(2022年3期)2022-06-11
意林(儿童绘本)(2019年10期)2019-12-23
上海包装(2019年8期)2019-11-11
小学生优秀作文(低年级)(2018年9期)2018-09-10
小布老虎(2017年1期)2017-07-18
天津造纸(2016年1期)2017-01-15
中华胃食管反流病电子杂志(2016年4期)2016-11-07
中国造纸学报(2015年1期)2015-12-16
中国医疗美容(2015年2期)2015-07-19
中国医学科学院学报(2015年5期)2015-03-01
