
镜湖角突臂尾轮虫种群增长参数的克隆多样性
2016-08-06 02:55曹宏源席贻龙
安徽师范大学学报(自然科学版) 2016年4期
关键词:克隆
彭 彬, 曹宏源, 潘 玲, 席贻龙
(安徽师范大学 生命科学学院,安徽 芜湖 241000)
镜湖角突臂尾轮虫种群增长参数的克隆多样性
彭彬,曹宏源,潘玲,席贻龙
(安徽师范大学 生命科学学院,安徽 芜湖241000)
摘要:以采自芜湖市镜湖水体中的11个角突臂尾轮虫(Brachionus angularis)克隆为对象,在(25±1)℃,0.5×106、1.0×106和2.0×106个细胞/mL的斜生栅藻(Scenedemus obliquus)密度下,应用群体累积培养法研究了角突臂尾轮虫种群增长参数的克隆多样性.结果表明,轮虫的种群增长率、最大种群密度、种群中的平均混交率和平均受精率与食物密度之间的关系具有较高的克隆多样性,食物密度、克隆以及两者间的交互作用对轮虫种群增长率、最大种群密度、平均混交率和平均受精率均有极显著性影响(P<0.01).研究角突臂尾轮虫种群增长参数的克隆在所有的食物密度和克隆组合中,克隆2在2.0×106个细胞/mL食物密度下的种群增长率和最大种群密度均最高,克隆3在1.0×106个细胞/mL食物密度下平均混交率最高,克隆3和9在2.0×106个细胞/mL食物密度下的平均受精率最高.因此,在开展角突臂尾轮虫的规模化培养时,拟首选克隆2和2.0×106个细胞/mL的斜生栅藻密度;而欲开展角突臂尾轮虫休眠卵的批量生产,则拟选择克隆3和1.0×106个细胞/mL的斜生栅藻密度.
关键词:角突臂尾轮虫;克隆;食物密度;种群增长参数
轮虫是鱼类等水产经济动物幼苗的理想开口饵料,其在水产养殖上的应用始于20世纪60年代末.在使用轮虫作为开口饵料时,人们必须关注轮虫的大小、轮虫在水体中的分布和密度、轮虫的保障程度、轮虫是否易于被消化和吸收以及轮虫的营养质量等问题,其中,如何获得大量的轮虫以持续满足水产经济动物苗种生产的需要始终是水产养殖研究人员和从业者关注的问题[1].此问题的解决涉及轮虫种群增长速率的提高和种群密度高峰期的维持等方面.
影响特定种类的轮虫种群增长速率的因素主要有食物种类和密度、温度、水体pH值和轮虫的品系或克隆等[2-6].在常规条件下开展轮虫的规模化培养时,筛选具有较高种群增长速率的轮虫品系或克隆便成为研究工作的重点[2].角突臂尾轮虫是淡水中常见的轮虫种类之一,在水产养殖和环境监测中具有重要的应用价值.作者已开展的试验表明,与投喂奶粉相比,投喂该种轮虫可显著提高泥鳅(Misgurnus anguillicaudatus)的幼苗成活率.有关角突臂尾轮虫的种群增长率等研究已有一些报道,胡好远等[3,4]曾就食物密度对角突臂尾轮虫种群增长的影响等进行专门的研究,但有关其不同克隆间种群增长速率等的差异性研究尚未见报道.本文以采自芜湖市镜湖中的11个角突臂尾轮虫克隆为对象,在三种斜生栅藻密度下,应用群体累积培养法研究了角突臂尾轮虫种群增长参数的克隆多样性,旨在揭示各克隆角突臂尾轮虫的种群增长速率等之间的差异及其与食物密度之间的关系,同时为该种轮虫的规模化培养筛选适宜的克隆.
1材料与方法
1.1轮虫的来源和培养
实验所用角突臂尾轮虫于2014年4月采自芜湖市镜湖.样品采集后,显微镜下随机挑取带卵的非混交雌体于自然光照、(25±1)℃条件下进行克隆培养,培养液为新鲜配制的轮虫培养液[7],食物为HB-4培养基[8]培养的、处于指数增长期的斜生栅藻.
1.2种群增长实验
实验前,将每个轮虫克隆分别置于自然光照、(25±1)℃以及0.5×106、1.0×106和2.0×106个细胞/mL等3个斜生栅藻密度下进行为期一周的预培养.预培养时,以直径为1cm、长度为12cm 的玻璃试管为容器,培养体积为5mL.预培养过程中,每天更换轮虫培养液并投喂相应密度的藻类食物一次,同时通过去除一部分个体使得轮虫种群始终处于指数增长期.
实验开始时,从每个克隆中随机挑取 200 个以上携卵的非混交雌体置于试管中继续培养.4 h后,从各试管中挑取龄长在4h以内的幼体于玻璃杯中继续培养,待其产出第一枚非混交卵时接种至玻璃试管内开始实验,接种密度为4个/mL,培养液体积为5mL.培养条件与预培养时相同.实验过程中,每12h轻轻吹浮沉积于试管底部的藻类食物,每24h对培养液中轮虫各类型雌体计数1次,并投喂饵料.待种群密度较大时,采用抽样计数,用玻璃微吸管将培养液轻轻搅匀后,从试管的3个不同部位于水体中部吸取(0.5-2)mL的培养物进行计数.雌体类型的划分和鉴定与席贻龙和黄祥飞相同[9].
1.3相关参数的定义与计算方法
轮虫种群增长率的计算公式为r=(lnNt-lnN0)/t,式中:N0为起始轮虫密度,即4个/mL;Nt为试验进行到t天时的轮虫种群密度;t为试验天数.参照Dumont等[10]的方法,从轮虫种群指数增长期选取4-6个数据计算种群增长率并最终获得每个重复的平均值.
最大种群密度:实验过程中轮虫种群密度的最大值.
混交雌体百分率:种群中的混交雌体数占总雌体数的百分比,本文使用的是试验期间每日混交雌体百分率的平均值.
混交雌体受精率:种群中产休眠卵的混交雌体数占总混交雌体数的百分比,本文使用的是试验期间每日混交雌体受精率的平均值.
1.4数据的统计分析
采用SPSS 16.0统计分析软件对上述数据进行统计分析.对所得的各种群参数进行正态分布检验后,对符合正态分布的数据进行单因素和双因素方差分析以揭示克隆和食物密度对轮虫各种群参数的影响,应用SNK检验法对同一克隆下各食物密度间、同一食物密度下各克隆间有统计学意义(P<0.05)的轮虫各种群参数的均值作多重比较.
2结果与分析
2.1种群增长率
3个食物密度下,11个角突臂尾轮虫克隆的种群数量动态如图1所示.根据图1所示的种群数量动态计算所得的种群增长率和最大种群密度列于表1和3.
0.5×106、1.0×106和2.0×106个细胞/mL食物密度下,克隆对轮虫种群增长率都具有显著性影响(P<0.05).0.5×106和1.0×106个细胞/mL食物密度下,克隆1和6的种群增长率均趋于最低,克隆2的种群增长率都趋于最大;2.0×106个细胞/mL食物密度下,克隆6的种群增长率最低,但克隆1、2和11的种群增长率最高(表1).
除克隆3、4、9和10的种群增长率均不受食物密度的显著性影响外(P>0.05),克隆1、2、5、6、7、8和11的种群增长率均受食物密度的极显著性影响(P<0.05).其中,克隆1、2和5的种群增长率均在0.5×106和1.0×106个细胞/mL食物密度下无显著性差异,但两者均显著低于2.0×106个细胞/mL食物密度下;克隆7和11的种群增长率均在0.5×106个细胞/mL食物密度下最低,在2.0×106个细胞/mL下最高;克隆6和8的种群增长率对升高的食物密度所作出的反应与其它克隆均不相同(表1).

表1 不同食物密度下11个角突臂尾轮虫克隆的种群增长率(平均数±标准误)
* 同行相同大写字母表示食物密度间无显著性差异,同列相同小写字母表示克隆间无显著性差异.下同.

时间Time(d)图1 不同食物密度下11个角突臂尾轮虫克隆的种群数量动态Fig.1 Population dynamics of eleven Brachionus angularis clones at different food levels
双因素方差分析结果显示,食物密度、克隆以及两者间的交互作用对角突臂尾轮虫种群增长率均有极显著影响(P<0.01)(表2).在所有的食物密度和克隆组合中,克隆2在2.0×106个细胞/mL食物密度下的种群增长率最高.

表2 食物密度和克隆对角突臂尾轮虫种群增长率、最大种群密度、平均混交率和平均受精率的影响(F值)
**P<0.01
2.2最大种群密度
0.5×106、1.0×106和2.0×106个细胞/mL食物密度下,克隆对轮虫最大种群密度都具有极显著性影响(P<0.01).0.5×106和2.0×106个细胞/mL食物密度下,克隆2的最大种群密度均趋于最高;1.0×106个细胞/mL食物密度下,克隆5、10和11的最大种群密度最高(表3).
所有克隆的最大种群密度均受食物密度的显著性影响(P<0.05).克隆1、2、4、7、8和9的最大种群密度均在0.5×106和1.0×106个细胞/mL食物密度下无显著性差异,但两者均显著低于2.0×106个细胞/mL食物密度下;克隆3、5和11的最大种群密度均在0.5×106个细胞/mL食物密度下最低,在2.0×106个细胞/mL下最高;克隆6的最大种群密度在1.0×106个细胞/mL食物密度下较高,在0.5×106和2.0×106个细胞/mL食物密度下较低(表3).
方差分析结果显示,食物密度、克隆差异及其两者间的交互作用对角突臂尾轮虫最大种群密度均有极显著影响(表2).在所有的食物密度和克隆组合中,克隆2在2.0×106个细胞/mL食物密度下的最大种群密度最高.
2.3平均混交率
0.5×106、1.0×106和2.0×106个细胞/mL食物密度下,克隆对轮虫种群中的平均混交率都具有极显著性影响(P<0.01).三个食物密度下,轮虫种群中克隆2、3和9的平均混交率最高(表4).

表3 不同食物密度下11个角突臂尾轮虫克隆的最大种群密度
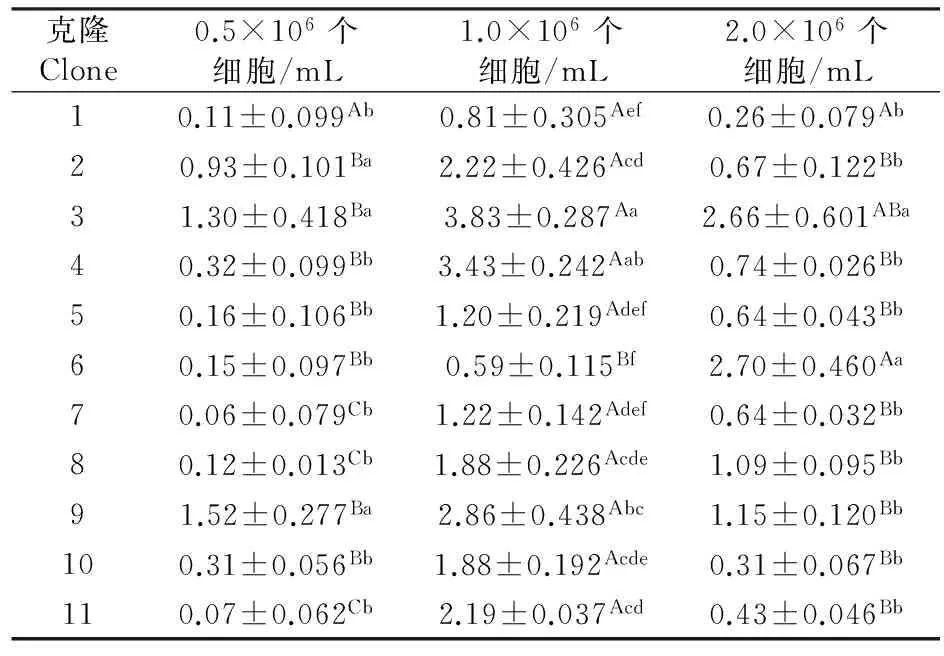
表4 不同食物密度下11个角突臂尾轮虫克隆的平均混交率(%)
除克隆1外,其它所有克隆的平均混交率均受食物密度的显著性影响(P<0.05).克隆2、4、5、9和10的平均混交率均在0.5×106和2.0×106细胞/mL食物密度下无显著性差异,但两者均显著低于1.0×106个细胞/mL食物密度下;克隆3的平均混交率在1.0×106个细胞/mL食物密度下高于0.5×106个细胞/mL食物密度下,但两者均与2.0×106个细胞/mL食物密度下无显著性差异;克隆6的平均混交率在2.0×106个细胞/mL食物密度下较高,在0.5×106和1.0×106个细胞/mL食物密度下较低;克隆7、8和11的平均混交率在0.5×106个细胞/mL食物密度下最低,在1.0×106个细胞/mL下最高(表4).
方差分析结果显示,食物密度,克隆差异及其两者间的交互作用对角突臂尾轮虫平均混交率均有极显著影响(表2).在所有的食物密度和克隆组合中,克隆3在1.0×106个细胞/mL食物密度下平均混交率最高.
2.4平均受精率
0.5×106和2.0×106个细胞/mL食物密度下,克隆对轮虫种群中的平均受精率都具有极显著性影响(P<0.01);但1.0×106个细胞/mL食物密度下,克隆对轮虫种群中的平均受精率无显著性影响(P>0.05).0.5×106个细胞/mL食物密度下,克隆9的平均受精率最高;2.0×106个细胞/mL食物密度下,克隆3的平均受精率最高(表5).
所有克隆的平均受精率均受食物密度的显著性影响(P<0.05).克隆1和10的平均受精率在0.5×106个细胞/mL食物密度下较低,在1.0×106和2.0×106个细胞/mL食物密度下较高;克隆2-8的平均受精率均在0.5×106和1.0×106个细胞/mL食物密度下无显著性差异,但两者均显著低于2.0×106个细胞/mL食物密度下;克隆9的平均受精率在2.0×106个细胞/mL食物密度下高于1.0×106个细胞/mL食物密度下,但两者均与0.5×106个细胞/mL食物密度下无显著性差异;克隆11的平均受精率在2.0×106个细胞/mL食物密度下最高,在0.5×106个细胞/mL食物密度下最低(表5).
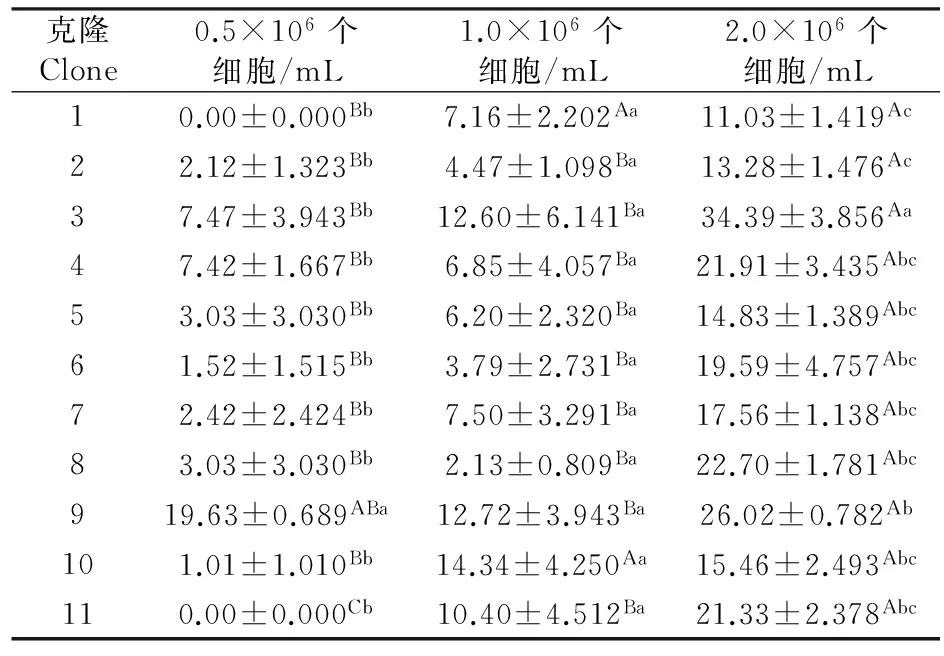
表5 不同食物密度下11个角突臂尾轮虫克隆的平均受精率(%)
方差分析结果显示,食物密度,克隆差异及其两者间的交互作用对角突臂尾轮虫平均受精率均有极显著影响(表2).在所有的食物密度和克隆组合中,克隆3和9在2.0×106个细胞/mL食物密度下的平均受精率最高.
3讨论
3.1食物密度和遗传因素对轮虫孤雌生殖速率的影响
已有研究结果显示,食物密度对轮虫种群增长率和最大种群密度的影响因轮虫种类或品系的不同而异[3-6].25℃下,采自广州的角突臂尾轮虫种群增长率在8.25×106个细胞/mL的蛋白核小球藻(Chlorella pyrenoidosa)密度下最高[3],褶皱臂尾轮虫(B. plicatilis)种群增长率在20.0×106个细胞/mL的酵母密度下最高,而最大种群密度出现在10.0×106个细胞/mL的酵母密度下[5];24℃下,萼花臂尾轮虫种群增长率在1.0×106、2.0×106和4.0×106个细胞/mL的斜生栅藻密度间无显著的差异[6].25℃下,采自广州、芜湖和颖上的角突臂尾轮虫种群增长率分别在10.47×106、7.47×106和7.24×106个细胞/mL的蛋白核小球藻密度下达最大值[4].本研究结果显示,食物密度对角突臂尾轮虫种群增长率和最大种群密度的影响在轮虫克隆间存在着差异,同一食物密度下角突臂尾轮虫种群增长率和最大种群密度在轮虫克隆间也存在着差异,此与Snell & Carrilo[2]、胡好远等[4]和Ma等[11]有关不同地理品系褶皱臂尾轮虫和角突臂尾轮虫、不同地理种群的萼花臂尾轮虫的研究结果均一致.
3.2食物密度和遗传因素对轮虫有性生殖幅度的影响
混交雌体的产生是轮虫有性生殖的开始,受众多的内源性和外源性因素的影响和制约.食物密度对轮虫种群中的平均混交率和平均受精率的影响也因轮虫种类或品系的不同而存在着差异[6,12,13].单个体培养条件下,萼花臂尾轮虫在适宜的蛋白核小球藻密度(1.0-2.0×107个细胞/mL)下后代的混交率最低,高于或低于此密度后代混交率显著提高[13];群体培养状态下,萼花臂尾轮虫种群中的平均混交率在8.0×106个细胞/mL斜生栅藻密度下最低[12].24℃下,萼花臂尾轮虫的平均混交率在1.0×106个细胞/mL食物密度下显著高于2.0×106个细胞/mL食物密度下,而4.0×106个细胞/mL食物密度下的平均混交率与前两者间均无显著的差异[6].本研究结果显示,食物密度对角突臂尾轮虫种群中的平均混交率的影响在轮虫克隆间存在着差异,同一食物密度下角突臂尾轮虫种群中的平均混交率在轮虫克隆间也存在着差异,此与Ma等[11]有关不同地理种群的萼花臂尾轮虫的研究结果一致.
Snell & Carrilo[2]和Ma等[11]认为,不同地理品系的褶皱臂尾轮虫和不同地理种群的萼花臂尾轮虫种群增长率等参数间的差异主要是由各品系轮虫遗传上的差异造成的.本研究中,各克隆角突臂尾轮虫间的遗传差异如何有待进一步研究.
3.3轮虫克隆间生殖差异研究的实践意义
对轮虫不同克隆间种群增长速率和有性生殖发生幅度的比较研究,有助于筛选具有较高种群增长速率和较高有性生殖幅度的轮虫克隆,推动轮虫的规模化培养或其休眠卵的批量生产,促进轮虫在水产养殖业中的应用.就本研究而言,在所有的食物密度和克隆组合中,克隆2在2.0×106个细胞/mL食物密度下的种群增长率和最大种群密度均最高,克隆3在1.0×106cells/ml食物密度下平均混交率最高.因此,在开展角突臂尾轮虫的规模化培养时,拟首选克隆2和2.0×106个细胞/mL的斜生栅藻密度.而欲开展角突臂尾轮虫休眠卵的批量生产,则拟选择克隆3和1.0×106个细胞/mL的斜生栅藻密度.
参考文献:
[1]刘桂云,席贻龙.轮虫与海产鱼类的苗种生产[J].安徽师范大学学报:自然科学版, 2003,26(3):268-271.
[2]SNELL T W, CARRILO K. Body size variation among strains of the rotifers Brachionus plicatilis[J]. Aquaculture, 1984,73:359-367.
[3]胡好远,席贻龙,耿红.食物密度对角突臂尾轮虫种群增长、个体大小、 卵体积的影响[J].应用生态学报,2002a,13(7):875-878.
[4]胡好远,席贻龙,耿红.食物密度对三个品系角突臂尾轮虫种群增长、个体大小、卵体积的影响[J].动物学研究,2002b,23(5):384-388.
[5]董婧,于叶辉, 赵莹莹,等.温度和食物对褶皱臂尾轮虫种群增长及繁殖的影响[J].现代畜牧兽医,2014,4:35-39.
[6]王敏,席贻龙,周彬,等.利用种群增长研究镜湖夏季萼花臂尾轮虫对水温的适应[J].水生生物学报,2014,38(6):1018-1023.
[7]GILBERT J J. Mictic female production in rotifer Brachionus calyciflorus[J]. Journal of Experimental Zoology, 1963,153(2):113-124.
[8]章宗涉,黄祥飞.淡水浮游动物研究方法[M].北京:科学出版社,1991,364-367.
[9]席贻龙,黄祥飞.种群起始密度对萼花臂尾轮虫休眠卵形成的影响[J].应用生态学报,2000,11(3):273-276.
[10]DUMONT H J, SARMA S S S, ALI A J. Laboratory studies on the population dynamics of Anuraeopsis fissa (Rotifera) in relation to food density[J]. Freshwater Biology, 1995,33:39-46.
[11]MA Qin, XI Yi-Long, ZHANG Jin-Yan, et al. Differences in asexual and sexual reproduction among eight geographic populations of Brachionus calyciflorus (Rotifera) from China[J]. Journal of Freshwater Ecology, 2009,24(2):273-278.
[12]陈芳,席贻龙,徐小平.食物浓度对萼花臂尾轮虫混交雌体形成的影响[J].安徽师范大学学报,2005,28(4):452-455.
[13]杨家新,黄祥飞.藻类对萼花臂尾轮虫繁殖的影响[J].湖泊科学,1998,10(1):42-48.
DOI:10.14182/J.cnki.1001-2443.2016.04.012
收稿日期:2015-11-15
基金项目:国家自然科学基金(31170395);国家星火计划重大项目(2013GA710001).
作者简介:彭彬(1990-),男,安徽芜湖人,硕士研究生;通讯作者:席贻龙(1965-),男,安徽肥东人,教授,博士.
中图分类号:Q178.1
文献标志码:A
文章编号:1001-2443(2016)04-0371-06
Clonal Diversity of Population Growth Parameter of Brachionus Angularis from Lake Jinghu
PENG Bin,CAO Hong-yuan,PAN Ling, XI Yi-long
(College of Life Sciences, Anhui Normal University, Wuhu 241000, China)
Abstract:By means of population accumulative culture and with Scenedemus obliquus as the rotifers’ food, the population growth parameters of eleven Brachionus angularis clones collected from Lake Jinghu were investigated at (25±1)℃ and three food levels (0.5×106, 1.0×106 and 2.0×106 cells/mL). The results showed that the relationships between population growth rate, maximal population density, mean mixis rate as well as mean fertilization rate and food level differed with clone. Population growth rate, maximal population density, mean mixis rate and mean fertilization rate were all affected very significantly by food density, clone and their interactions (P<0.01). Among all the combinations of food levels and clones, clone 2 cultured at 2.0×106 cells/mL of S. obliquus had the highest population growth rate and maximal population density, clone 3 cultured at 1.0×106 cells/mL of S. obliquus had the highest mean mixis rate, and clone 3 and 9 cultured at 2.0×106 cells/mL of S. obliquus had the highest mean fertilization rate. Clone 2 of B. angularis was the optimal for mass culture.
Key words:Brachionus angularis; clone; food level; population growth parameter
引用格式:彭彬,曹宏源,潘玲,等.镜湖角突臂尾轮虫种群增长参数的克隆多样性[J].安徽师范大学学报:自然科学版,2016,39(4):371-376.
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
电脑报(2020年28期)2020-07-31
河北果树(2020年2期)2020-05-25
小学科学(学生版)(2019年5期)2019-05-21
中央民族大学学报(自然科学版)(2018年1期)2018-06-27
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
时代英语·高二(2015年2期)2015-05-18
现代检验医学杂志(2015年4期)2015-02-06
