
不同施肥水平对桢楠多胚苗生长及光合生理的影响
2016-08-06 05:37胡红玲胡庭兴陈玉凤廖玉华周光良杨珊珊
西北植物学报 2016年6期
谭 飞,胡红玲,胡庭兴,陈玉凤,廖玉华,周光良,罗 杰,杨珊珊
(四川农业大学 林学院,成都 611130)
不同施肥水平对桢楠多胚苗生长及光合生理的影响
谭飞,胡红玲*,胡庭兴,陈玉凤,廖玉华,周光良,罗杰,杨珊珊
(四川农业大学 林学院,成都 611130)
摘要:采用营养袋培育的方法,以苗龄 45 d(根茎初步木质化)的桢楠幼苗为试验材料,选取单胚苗(D)、双胚大苗 (SD)和双胚小苗 (SX) 3种类型桢楠幼苗栽植于袋中,根据年施复合肥量标准设置对照(CK,每袋0 g)、中(M,每袋0.2 g)和高(H,每袋0.3 g)3个施肥水平,于7月初开始施肥,每20 d施1次共分4次(每次25%)施入,分别于8月、10月中旬测定幼苗生长指标和光合生理指标,探讨施肥对桢楠多胚苗生长和光合生理的影响。结果显示:(1)中量施肥(M)对3类幼苗的苗高和地径生长量有明显的促进作用,而高量施肥(H)在整个试验过程中对幼苗生长的影响表现为先促进后抑制的趋势。(2)中量施肥(M)可以促进3类幼苗叶片的叶绿素合成,延长叶片功能周期,增大净光合速率和气孔导度,进而提高幼苗在强光照、高CO2环境下的光合作用能力。(3)中量施肥可以使幼苗的光补偿点(LCP)、CO2补偿点(CCP)下降,光饱和点(LSP)、CO2饱和点(CSP)上升,增强幼苗适应环境的能力,积累更多的有机物;高量施肥对幼苗的光合指标也表现为先促进后抑制的作用。研究表明,适量(每袋0.2 g)施肥能促进各类桢楠幼苗光合色素合成、光合作用效率以及对环境的适应能力,从而促进幼苗的生长;适量施肥也使双胚大苗较快缩小与单胚苗生长指标间的差距,同时也对双胚小苗生长表现出一定的促进作用,这对于促进珍贵树种桢楠的种质资源保护、提高苗木出圃率和降低育苗成本有重要意义。
关键词:桢楠;多胚苗;施肥;生长;光合生理
桢楠(Phoebezhennan)为樟科(Lauraceae)楠属(Phoebe)常绿乔木,为国家二级保护渐危种,是组成常绿阔叶林的主要树种。桢楠树干通直,树姿优美,既是上等的用材树种,又是极好的绿化树种[1-2]。由于历代过度砍伐利用,致使这一珍贵的树种资源近于枯竭。而现存林分多系人工栽培的半自然林和风景保护林,在庙宇、公园、庭院等处尚有少量的大树,但大多过熟,且病虫危害较严重,相继衰亡。因其生长缓慢和种苗资源缺乏,严重制约了楠木人工林的发展[3-4]。作者所在课题组在前期桢楠育苗过程中发现其种子具有多胚现象,且一些母树种子多胚率高达40%,因此拟通过对桢楠多胚苗生长动态和光合特性的研究,为桢楠苗木的培育及后续人工林的发展提供理论依据与技术参考。
目前,在被子植物中已有36个科415个种中发现了无融合生殖类型[5-6],其中至少有17个科47个属86个种的植物中有多胚现象存在,最早在1719年由Leeuwenhok在柑橘属(Citrus)[ 7-8]植物中发现,之后在水稻(Oryzasativa)上也发现了双胚苗含有低频率的无融合生殖方式[9-10],随后在其它作物如玉米(Zeamays)[11]、苎麻(Boehmerianivea)[12]韭菜(Alluimtuberosum)[13]、咖啡(Coffealiberica)[14]、芒果(Mangiferaindica)[15]等作物上也有过双胚苗的报道,2000年前后国内外曾掀起多胚苗现象研究的热潮[16-17]。
施肥是促进植物生长、提高植物产量的一种高效、低成本的耕作方式。在林业生产中,适量施肥可大幅度提高林木的生长量。目前,对于楠属植物的研究已有相关报道,主要以研究其生长规律、群落分布、资源保护及园林绿化利用为主,对容器育苗、繁殖技术和经营措施的研究则少有报道。其中,张群英探讨了密度和施肥对楠木播种苗生长的影响[18];董立军等[19]研究了施肥对闽楠[Phoebebournei(Hemsl.) Yang]等3种樟科植物容器苗生长的影响;贺维等[20]详细研究了施肥对桢楠幼苗光合生理、生长及土壤肥力的影响。虽然前人的研究已经涉及到了施肥对楠属植物生长的影响,但关于施肥对楠属植物多胚苗生长的影响未见报道。因此,本研究拟通过测定不同胚数桢楠幼苗形态以及光合生理参数对施肥的响应特征,初步探讨施肥对桢楠多胚苗生长的影响,为促进桢楠育苗成效,提高优良种质资源的利用率和推动其人工林资源的发展提供理论依据和技术参考。
1材料和方法
1.1试验地概况
试验地位于四川省雅安市四川农业大学科研园区(102°59′57″E,29°58′21″N)大棚内,海拔580 m。该地区属亚热带湿润气候,阴雨较多,光照不足,年均气温16.2 ℃,≥10 ℃年积温5 231 ℃,7月平均最高气温29.9 ℃,1月平均最低气温3.7 ℃,年均日照时数1 039.6 h,无霜期298 d,年均降水量1 774.3 mm,空气平均湿度79%。
1.2试验材料
试验采用统一规格的聚乙烯塑料营养袋(口径8 cm、高12 cm),土壤选择当地常见的农耕土,其主要理化性质为:质地中壤,有机质23.75 g·kg-1,pH 6.13,全氮1.54 g·kg-1,铵态氮1.75 mg·kg-1,全磷1.34 g·kg-1,全钾27.12 g·kg-1。使用前去除原有植物残体和少量石砾,并每袋加入复合肥0.05 g作基肥(每袋土壤重量500 g左右);2014年5月20日采用四川农业大学科研园区培育的桢楠幼苗90株(苗龄 45 d),及时移栽到营养袋(每袋1株)并适量浇水。栽植后进行常规水肥管理,用Hh1土壤水分速测仪(ML2x,GBR)监测并控制土壤含水量,保持袋内土壤湿度在18%左右(即本试验用土田间持水量的70% ~ 80% )。
1.3试验设计
根据苗圃楠木种子的出苗情况(主要为单胚和双胚苗,极少数为3胚及以上),本试验选用3类幼苗,分别是单胚苗(D)、双胚大苗(SD)和双胚小苗(SX)。选用复合肥(29% N、16% N、10% N)进行施肥,每类幼苗分别设置不施肥(CK)、中量(M)和高量(H)3个施肥水平,每个水平重复10次,总计90袋,并将每袋进行编号,分别观测记录幼苗的生长情况。
根据相关文献[21-22],林木1年生幼苗全年施氮量一般为45~90 kg·hm-2(按纯氮计算),本试验采用的营养袋面积约为0.005 m2,换算为每袋的年施氮量为0.045 g,根据复合肥的含氮量计算出年施肥量为0.2 g,作为基本量(记作中量M),并另设1.5倍基本量(0.3 g/袋,记作高量H),对照(CK)不施肥。将总施肥量分为4份,于7月初开始施肥,每隔20 d施 1次,共4次,施肥时直接将复合肥撒于土壤表层,再通过喷水溶解渗入土壤。在8月中旬(半量施肥处理后)和10月中旬(全量施肥处理后)测定各处理的幼苗株高及地径,同时用Li-6400便携式光合仪(Li-Cor Inc.,USA)测定各项光合指标,之后立即采叶片进行光合色素含量测定。
1.4测定指标及方法
1.4.1形态指标用直尺测量株高(0.01 cm),电子游标卡尺测量地径(0.01 mm),地径从两个相互垂直的方向测定,取其平均值作为实际地径。
1.4.2光合色素含量采用分光光度法[23]测定光合色素含量。剪取各植株成熟完好的叶片洗净剪成小碎片称量0.5 g加提取液(80%丙酮与无水乙醇1∶1混合)10 mL。取1 mL稀释至10 mL后分别在470、646和 663 nm处测定其吸光度A470、A646和A663。每处理重复测定3次。各色素含量的计算公式如下:
叶绿素a浓度Chl a=12.21A663-2.81A646
叶绿素b浓度Chl b=20.13A646-5.03A663
类胡萝卜素浓度Car=(1 000A470-3.27Chl a-104Chl b)/229
叶绿体色素的含量/(mg·g-1)=色素的浓度×提取液体积×稀释倍数/样品鲜重
1.4.3光合生理特征参数采用Li-6400光合仪,设定光照强度为1 200 μmol·m-2·s-1,CO2浓度为400 μmol·mol-1,温度为25 ℃,相对湿度为45%~65%,分别在8月中旬和10月中旬于各处理桢楠幼苗自顶端向下选取第一轮成熟功能叶片,测定其净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),每个处理测定5片叶子,每片叶子记录10个数据。
1.4.4光合-光响应曲线和光合-CO2响应曲线选取相同部位的成熟叶片,采用Li-6400光合仪测定各处理植株叶片光响应曲线。光合有效辐射(PAR)梯度从低到高设定为0、25、50、75、100、200、400、600、800、1 000、1 200、1 600、2 000 μmol·m-2·s-1,测定时叶室温度设为30 ℃,利用Li-6400-01液化CO2(400 μmol·mol-1)稳定CO2浓度;测定各处理植株叶片光合CO2响应曲线,温度设为30 ℃,光照强度设为1 200 μmol·m-2·s-1,CO2浓度梯度设定为0、20、50、80、100、120、160、180、200、400、800、1 000、1 200 μmol·mol-1。
1.5数据处理
采用直角双曲线模型进行光合-光响应和光合-CO2响应曲线拟合[24],并分析计算相应的特征参数。采用SPSS 20.0统计软件对试验数据进行单因素方差分析(One-way ANOVA),并用最小显著差数法(LSD法)进行多重比较,Sigmaplot 12.5进行图表制作。
2结果与分析
2.1施肥对桢楠多胚幼苗生长指标的影响
表1显示,在施肥处理前(2014年6月1日前),桢楠幼苗生长指标(地径和株高)在单胚苗(D)、双胚大苗(SD)和 双胚小苗(SX)3类幼苗间的差异均显著(P<0.05),而在同类幼苗各施肥处理间均没有显著差异(P>0.05)。施肥后,地径和株高的生长变化对施肥的响应基本一致。在施肥前期(7~8月),D和 SD两类幼苗各施肥处理间地径生长量无显著差异,而SX各施肥处理地径均显著低于D和SD相同施肥处理的地径,此期幼苗的株高生长量也有类似的表现;施肥后期(9~10月)各处理株高、地径生长量和总生长量显示,D和SD两类幼苗中量施肥水平(DM、SDM)的地径和株高生长量均显著高于其他施肥处理,表现出明显的促进作用,中量施肥对SX也有促进作用,但差异不显著;而高量施肥(H)对3类幼苗生长均呈现出明显的抑制作用。以上结果说明在各类桢楠幼苗植入营养袋初期少量的养分供应便可促进其生长,而过量的施肥反而抑制其生长。
2.2施肥对桢楠多胚苗叶片光合色素含量的影响
由表2可见,在8月份,桢楠单胚苗(D)、双胚大苗(SD)和 双胚小苗(SX)叶片叶绿素(Chl)总量、类胡罗卜素(Car)含量均随着施肥水平的增加呈先增后减的趋势,且3类幼苗都以中量施肥(M)促进效果最好,其叶绿素(Chl)总量与CK差异显著(P<0.05)。通过比较3类幼苗之间CK发现,SD和D类幼苗的叶绿素(Chl)总量差异不大,而SX幼苗的叶绿素含量要显著低于前两者。10月份的测定结果表明,施肥后期各类桢楠幼苗的光合色素含量变化趋势和上述生长指标变化基本保持一致,中量施肥(M)处理对各类幼苗光合色素含量有一定的促进作用,但与不施肥对照(CK)差异不显著,而高量施肥(H)处理却对各类幼苗光合色素含量表现出显著(P<0.05)的抑制作用。可见,桢楠双胚小苗的光合色素含量明显低于单胚苗和双胚大苗,苗期适量施肥可一定程度上增加各类幼苗的光合色素含量,但施肥过多也会抑制光合色素的合成。
2.3施肥对桢楠多胚苗的净光合速率及气体交换参数的影响
由图1可知,在不施肥条件下(CK),桢楠单胚苗(D)、双胚大苗(SD)和 双胚小苗(SX)3类幼苗的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)在两次测定(8月和10月)中总体上表现为SD≈D >SX,且10月份SX的这些指标与SD和D的相应指标间的差异缩小。在第2次施肥后(8月份),D、SD和SX 3类幼苗净光合速率及气体交换参数表现为随施肥量的增加而呈现先上升后下降的趋势,中量施肥(M)各类幼苗的Pn、Gs、Ci始终处于最高状态,并且与CK的相应值均表现出显著差异(P<0.05),而高量施肥(H)对这些气体交换参数的也有一定的促进作用,但不如M处理明显。 在第4次施肥后(10月份),D、SD和SX 3类幼苗的气体参数随着施肥水平的增加同样呈现先上升后下降的趋势;但与CK相比,M处理对各类幼苗的气体交换参数仍表现为不同程度的促进作用,而H处理对3类幼苗则都表现为显著的抑制作用(P<0.05)。以上结果说明苗期适量施肥可增加各类幼苗的光合参数值,过量施肥反而有一定的抑制作用。
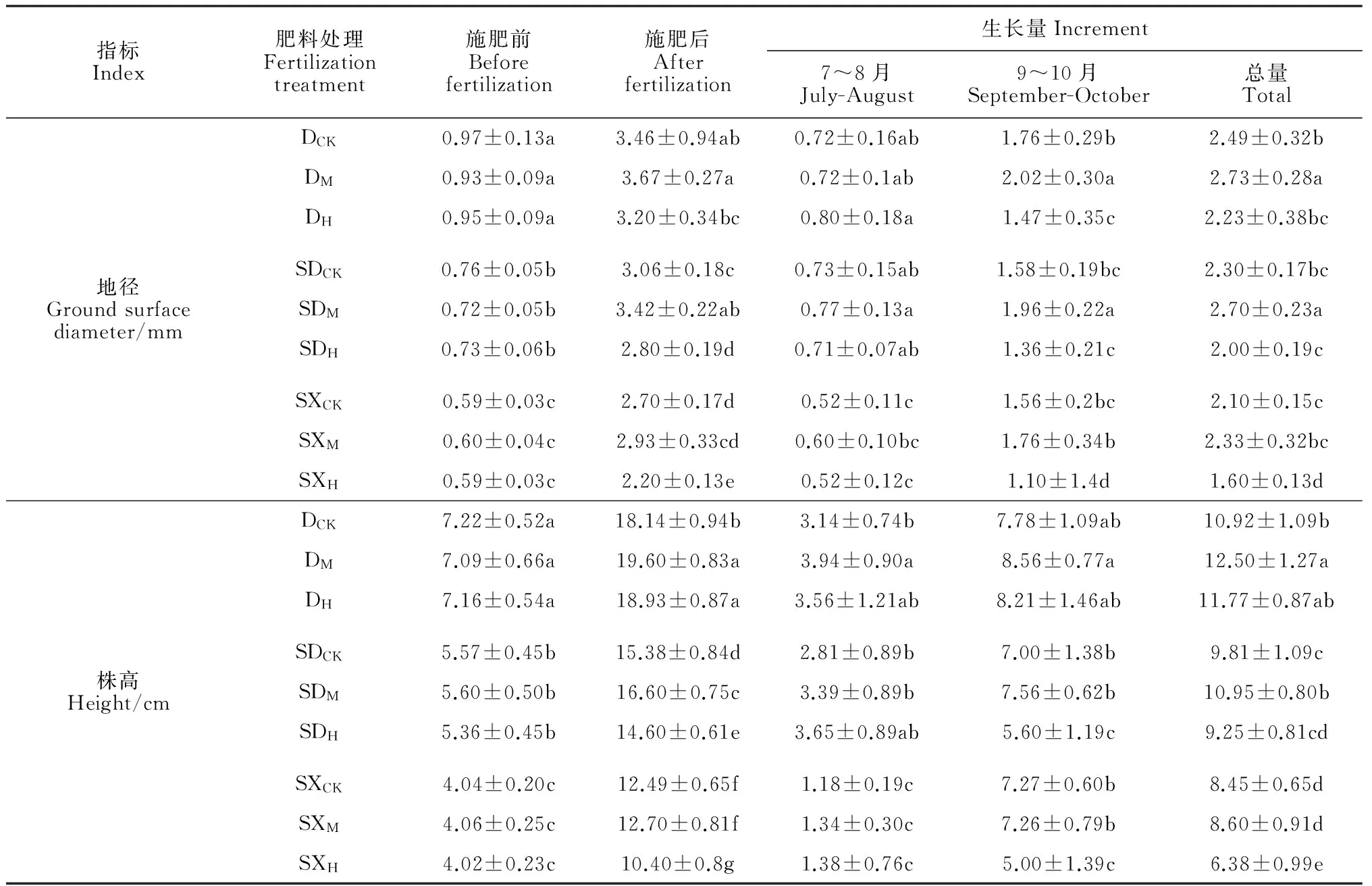
表1 不同施肥量处理桢楠多胚苗地径和株高在(6~ 10月)5个月内的变化
注: D、SD、 SX分别表示单胚苗、双胚大苗、双胚小苗;CK、M、H分别表示不施肥(对照)、中量施肥和高量施肥;同列不同小写字母表示差异达到0.05显著水平;下同
Note: D,SD and SX denote single embryo seedlings, bigger one of twin embryo seedlings and smaller one of twin embryo seedlings respectively; CK, M and H denote without, moderate and high amount of fertilizer applied;Different letters in the columns indicate significant differences at 0.05 level among the treatments, respectively; The same as below

表2 不同施肥量处理桢楠多胚苗光合色素含量的变化
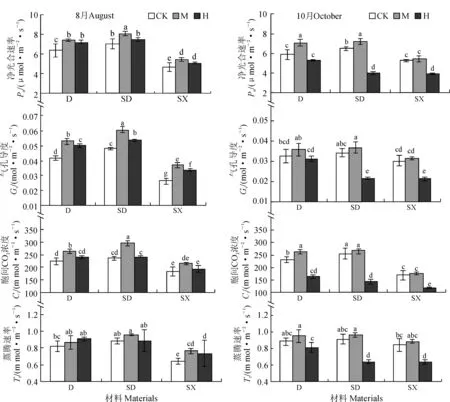
不同小写字母表示不同处理间差异达0.05显著水平;D、SD、SX分别代表了单胚苗、双胚大苗、双胚小苗;CK、M、H代表不同的施肥水平。下同图1 不同施肥量处理的桢楠多胚苗叶片净光合速率及气体交换参数Different normal letters over the bars indicate significant differences among the treatments at 0.05 level,respectively; The same as belowFig. 1 The net photosynthetic rate and gas exchange parameters of poly embryo seedlings of P. zhennan S. Lee under different levels of fertilizer application
2.4施肥对桢楠多胚苗叶片光响应曲线、CO2响应曲线及其特征参数的影响
2.4.1光响应曲线及其特征参数3类幼苗各施肥处理的光响应曲线变化趋势如图2所示。施肥45 d后(8月份),3类幼苗D、SD、SX的净光合速率(Pn)当光合有效辐射(PAR)在0~600 μmol·m-2
·s-1区间内时均随PAR的增加而迅速增加,当PAR高于600 μmol·m-2·s-1时,各处理的Pn值增幅减小,最终趋于稳定,3类幼苗Pn总体呈现出SD≈D>SX的趋势;但随着施肥量和处理时间的不同,在同一光照强度下各施肥处理间Pn的值有较大差异,基本表现为 M>H>CK,说明初期的施肥对桢楠各类幼苗生长均有较大的促进作用。同时,由表3可知,随着施肥水平的增加, 3类桢楠幼苗的最大净光合速率(Pn max)、光饱和点(LSP)与光响应曲线的变化趋势一致(M>H>CK),而光补偿点(LCP)则呈现相反的趋势(M 在10月份,当PAR接近500 μmol·m-2·s-1时,3类幼苗各施肥处理的Pn值已经趋于稳定,其光响应曲线的变化趋势以及Pn max、LSP都呈现出M>CK>H的现象,且3类幼苗施肥处理间的差异比8月份更大;说明此时中量施肥对桢楠各类幼苗的光合作用仍有一定的促进效果,而高量施肥处理则由前期的促进转变为显著的抑制作用。 2.4.2CO2响应曲线及特征参数由图3可见,在8月和10月份3类桢楠幼苗各施肥处理的Pn-CO2响应曲线同样符合植物光合CO2响应的变化规律,即在低CO2浓度范围内(0~800 μmol·mol-1),随着CO2浓度的增强,各处理的幼苗叶片Pn迅速上升,最后趋于稳定。在8月份2次施肥处理后,3类幼苗中的中量施肥(M)和高量施肥(H)处理的Pn-CO2响应曲线始终高于不施肥对照(CK),并呈现出M>H>CK的趋势,但相同CO2处理浓度下各施肥处理对应的Pn大多差异不显著。在10月份经4次施肥处理后,3类幼苗中的中量施肥(M)处理的Pn-CO2响应曲线始终高于高量施肥(H)和对照(CK),而高量施肥处理则处于最低状态,且相同CO2处理浓度下各施肥处理间差异显著。可见,3类桢楠幼苗之间,在相同CO2处理浓度下, SX对应的Pn要远小于SD、D,且差异显著。 图2 不同施肥量处理桢楠多胚苗的光响应曲线Fig. 2 The light response curve of poly embryo seedlings of P. zhennan S. Lee under different levels of fertilizer application 图3 不同施肥量处理桢楠多胚苗的CO2响应曲线Fig. 3 The CO2 response curve of poly embryo seedlings of P. zhennan S. Lee under different levels of fertilizer application. 同时,表3表明,8月份3类幼苗不同施肥水平处理的最大净光合速率(Pn max)、CO2饱和点(CSP)与Pn-CO2响应曲线之间存在相同的变化趋势,即M>H>CK;而其CO2补偿点(CCP)则呈现为CK>H>M的关系。在10月份,3类幼苗不同施肥处理间的Pn max、CSP表现为H 3讨论 大量研究表明,植物的多胚现象广泛存在于自然界中,目前对楠属多胚现象的研究已有报道,如黄宇研究发现闽楠种子在不同贮藏时间和不同培养条件下发芽率和多胚率都有所不同[25];余道平研究发现,桢楠多胚率达12.4%,其中双胚苗为9%,其余为3胚苗和4胚苗[26]。由于植物种子自身含有的营养成分有限,在育苗时会造成单胚苗和多胚苗在个体上有较大的差异,这对其后期的生长会产生很大的影响[27-28]。在本研究中,施肥初期双胚大苗(SD)和双胚小苗(SX)都与单胚苗(D)的生长指标存在一定的差距,而施肥后明显缩小了SD与D生长指标的差距,但SX却与D相差甚远。通过施肥来促进双胚苗的生长,提高育苗效率,有利于保存该珍贵树种更多的种质资源,繁育更多的园林绿化苗木并获得更多的经济效益。 前人在对各类树种幼树的施肥研究中发现,在一定的施肥量范围内,苗木的生物量随着养分的增加而增加,但超过最大吸收量反而会抑制幼苗的生长。如当年施氮量处于中氮水平下(0.6 g,桶口面积为0.07 m2),桢楠的生长最优,生长指标和光合参数都与对照有明显的差异[20];适量的氮、磷营养能够明显促进湿地松幼苗的生长、提高净光合速率,而氮磷营养过量时则会抑制幼苗的生长,降低叶片的净光合速率[29]。所以适量的养分供给才能使苗木的质量与产量达到最大化。本实验中,桢楠多胚苗的生长和光合生理指标的变化都与上述相关研究的结果一致。 表3 不同施肥处理桢楠多胚苗光响应和CO2响应特征参数 3.1施肥与桢楠多胚幼苗光合生理参数的变化 叶绿素含量的多少能一定程度上反映植物光合作用的强弱,叶绿素主要由含N化合物组成,所以施肥与它存在密切的关系。本试验发现,初期施肥能够增加桢楠幼苗叶片的叶绿素含量,提高净光合速率,从而增加光合产物的积累,同时D、SD和SX 3类幼苗在中量施肥(M)都表现出明显的生长优势,然而多次施肥后发现高量施肥(H)会减少叶绿素的含量,降低光合速率,从而抑制幼苗的生长,但长势稍好的DH受到的抑制不明显,长势较弱的SDH、SXH则受到了严重的抑制,这与幼苗对氮的吸收、转化和利用有直接关系。合理施肥可以延缓叶片衰老,延长生理周期,积累更多的有机物[30],本研究也发现施肥使得桢楠幼苗的类胡萝卜素有所提高,而后期高量施肥则会抑制幼苗类胡罗卜素的合成,促使植物叶片加速衰老。 其次,本研究中还发现,施肥初期3类幼苗各施肥处理的中量施肥(M)和高量施肥(H)都对净光合气体交换参数有显著促进作用,这与钱燕萍和袁颖红得到的结论相同[31-32];而在施肥后期中量施肥(M)的促进作用有所减弱,高量施肥则对净光合速率和气体交换参数表现出一定的抑制作用,这与王力朋[33]的研究结果一致。此外,从桢楠幼苗的光响应曲线和CO2响应曲线来看,施肥初期3类幼苗的最大净光合速率(Pn max)随着施肥水平的增加而增加,同时施肥又使幼苗的光补偿点(LCP)、CO2补偿点(CCP)都有所下降,使幼苗能够更好地在弱光和低浓度的CO2环境条件下生长,而光饱和点(LSP)和CO2饱和点(CSP)都有所上升,表明施肥可以提高桢楠幼苗在强光照、高CO2环境下的光合作用能力,促进幼苗积累更多的有机物质,这与杨自立的研究结果相似[34];而在施肥结束后高量施肥对3类幼苗的2种响应特征参数均有明显的抑制作用。 另外,在各类桢楠幼苗间,SDM的叶绿素、类胡萝卜素、光合气体参数以及响应曲线的最大净光合速率与DCK的相应指标相比虽然差异性仍然显著,但其差值均有所减小,表明SDM与DCK生理指标的差距不断缩小,且SXM与DCK的差距在2次的测定中也相继缩小,表明适量施肥对SX的生长也有一定促进作用。 3.2施肥与桢楠多胚幼苗生长参数的变化 在本试验施肥初期,不同施肥水平对3类幼苗生长均有一定促进作用,但差异不是特别显著,可能是因为选用的农田土较肥沃,以及添加基肥过多造成各处理之间差异不是很大。 而到试验后期,中量施肥对幼苗生长表现出明显的促进作用,而高量施肥对D、SD和SX生长的抑制作用依次增强,表明生长越弱的植株在土壤养分过多的情况下受到的抑制作用更强。所以,在中量施肥(M)条件下,D与SD桢楠幼苗能够快速生长,并且SD能够缩小与D的生长指标之间的差距,而SX对施肥的响应不明显。但是,也有相关研究表明不同梯度复合肥的施肥量对幼苗生长的影响不显著;并且长期施用化学肥料容易导致土壤板结、酸化,残留养分容易以地表径流、侵蚀和淋溶等方式损失,并造成环境污染[35-36],因此选择高效、低成本、无公害的有机肥对桢楠多胚幼苗进行施肥可能获得更好的效果,这将有待于开展进一步的研究。 综上所述,在本试验中,复合肥中量施用能有效促进3类桢楠幼苗光合色素合成、增强光合作用效率以及环境的适应能力,从而对桢楠多幼苗的生长有显著的促进作用,而高量施用对3类幼苗生长则有一定的抑制作用;中量施肥水平则能较快缩小双胚大苗(SD)与单胚苗(D)生长指标间的差距,对双胚小苗(SX)生长也有一定的促进作用,总体上增加了合格苗数量,提高苗木出圃率,减少育苗成本。 参考文献: [1]李冬林, 金雅琴, 向其柏. 我国楠木属植物资源的地理分布、研究现状和开发利用前景[J]. 福建林业科技, 2004, 31(1): 5-9. LI D L, JIN Y Q, XIANG Q B. The geographical distribution, research status and development alutilization prospect ofPhoebeNeesplant resource of our country[J].JournalofFujianForestryScienceandTechnology, 2004, 31(1): 5-9. [2]向其柏,季春峰. 浙江楠后选模式标本的重新指定[J]. 南京林业大学学报(自然科学版),2013,4:163-164. XIANG Q B, JI C F. On the redesignation of lectotype of Phoebe chekiangensis[J].JournalofNanjingForestryUniversityNaturalScienceEdition, 2013, 4: 163-164. [3]龙汉利,张炜,宋鹏等.四川桢楠生长初步分析[J].四川林业科技,2011,32(4):89-91. LONG H L, ZHANG W, SONG P,etal. An analysis of the growth ofPhoebezhennanin Sichuan[J].JournalofSichuanForestryScienceandTechnology, 2011, 32(4): 89-91. [4]张炜,何兴炳,唐森强,等.四川桢楠生长特性与分布[J].林业科技开发,2012,26(5):38-41. ZHANG W, HE X B, TANG S Q,etal. Study on the growth characteristics and distribution ofPhoebezhennanin Sichuan[J].ChinaForestryScienceandTechnology, 2012, 26(5): 38-41. [5]蔡雪, 母锡金, 朱至清, 等. 几种具无融合生殖特性的植物多胚和多现苗象的观察[J]. 植物学报, 1997, 39(7): 590-595. CAI X, MU X J, ZHU Z Q ,etal.Polyembryony and multiple seedlings in the apomictic[J].PlantScienceJournal, 1997, 39(7): 590-595. [6]母锡金, 蔡雪, 孙德兰, 等. 被子植物的无融合生殖和它的应用前景[J]. 作物学报, 2001, 27(5): 590-599. MU X J, CAI X, SUN D L ,etal.Apomixis and Its Application prospect[J].ActaAgronomicaSinica, 2001, 27(5): 590-599. [7]张斯淇, 徐强, 邓秀新. 无融合生殖与柑橘多胚现象的研究进展[J]. 植物科学学报, 2014, 1: 88-96. ZHANG S Q, XU Q, DENG X X. Advances in apomixis and polyembryony research inCitrusplants[J].PlantSclenceJournal, 2014, 01: 88-96. [8]洪棋武, 李喜庆, 江东. 单胚清见橘橙×椪柑产生高比例多胚种子及多胚有性的分子检测[J]. 中国农业科学, 2003, 07: 829-833. HONG Q W , LI X D, JIANG D. High ratio of polyembryonic seeds in monoembryony Kiyomi× ponkan and molecular evidences against asexual origins for polyembryonies[J].ScientiaAgriculturaSincia, 2003, 7: 829-833. [9]邓鸿德. 多胚水稻胚位与苗位的观察研究[J]. 西北植物学报, 1992, 12(1): 1-8. DENG H D. Observation and analysis on the location of embryo and seedling polyembryonic rice[J].ActaBotanicaBoreali-OccidentaliaSinica, 1992, 12(1): 1-8. [10]黎垣庆, 袁隆平. 水稻双胚苗遗传学研究[J]. 作物学报, 1990, 16(2): 176-182. LI Y Q,YUAN L P.Genetic studies on rice double embryo seedlings[J].ActaAgronomicaSinica, 1990, 16(2): 176-182. [11]李春秋, 祁永红, 王巍. 玉米无融合生殖的诱发及其在玉米育种上的应用[J]. 农业与技术, 2005, 25(1): 69-74. LI C Q, QI Y H, WANG W. Effects of fertilizers on growth of Italian ryegrass seedling[J].Agriculture&Technology, 2005, 25(1): 69-74. [12]温岚, 喻春明, 王延周, 等. 多胚苎麻的诱导及多胚性遗传分析[J]. 中国麻业科学, 2011, 3: 136-141. WEN L,YU CH M,WANG Y Z,etal. Study of polyembryonic ramie strain induction and polyembryony heredity analysis[J].PlantfiberSciencesinChina, 2011, 3: 136-141. [13]冯辉, 翟玉莹. 韭菜多胚苗及其无融合生殖关系的研究[J]. 园艺学报, 2007, 34(1): 225-226. FENG H, ZHAI Y Y. Polyembryonic seedlings and the relationship between apomixia and the occurrences of polyembryonic seedlings in Chinese Chive[J].ActaHorticulturaeSinica2007, 34(1): 225-226. [14]莫饶,郑成木,朱稳,等. 咖啡多胚现象与多胚苗形态发育的研究[J]. 植物学通报, 2004, 2: 189-194. MO R, ZHENG C M,ZHU W,etal. The polyembryony and morphogenesis of multiseedlings in coffee[J].ChineseBulletinofBotany, 2004, 2: 189-194. [15]莫饶, 罗远华, 周世民. 几个品芒果种的胚性及多胚苗遗传分析[J]. 热带亚热带植物学报, 2005, 6: 475-479. MO R, LUO Y H, ZHOU S M. Polyembryony in mango (Mangifera indica L.) and genetic analysis[J].JournalofTropicalandSubtropicalBotany, 2005, 6: 475-479. [16]FILONOVA L H, VON ARNOLD S, DANIEL G,etal. Programmed cell death eliminates all but one embryo in a polyembryonic plant seed[J].CellDeathandDifferentiation, 2002, 9(10): 1 057-1 062. [17]GUPTA P, SHIVANNA K R, MOHAN RAM H Y. Apomixis and polyembryony in the guggul plant,Commiphorawightii[J].AnnalsofBotany, 1996, 78(1): 67-72. [18]张群英. 密度和施肥对楠木播种苗生长的影响[J]. 福建林业科技, 2011, 38(2): 81-83. ZHANG Q Y. Effects of density and fertilization onPhoebebourneiseedling growth[J].JournalofFujianForestryScienceandTechnology, 2011, 38(2): 81-83. [19]董立军, 朱晓婷, 林夏珍等. 施肥对三种樟科植物容器苗生长的影响[J]. 北方园艺, 2011, (13): 73-77. DONG L J, ZHU X T, LIN X Z,etal. Effects of fertilization on the growth of three Lauraceae seedlings in containers[J].NorthernHorticulture, 2011, (13): 73-77. [20]贺维, 胡庭兴, 王锐,等. 施肥对桢楠幼苗光合生理及生长特性的影响[J]. 西北植物学报, 2014, 34(6): 1 187-1 197. HEI W,HU T X,WANG R,etal.Effect of fertilization on photosynthetic physiology and growth characteristics ofPhoebezhennanseedlings[J].ActaBotanicaBoreali-occidentaliaSinica,2014, 34(6): 1 187-1 197. [21]沈国舫. 森林培育学[M]. 北京: 中国林业出版社, 2001. [22]陈宜木. 山茶属3种食用油植物苗期施肥效应研究[D]. 福州: 福建农林大学, 2012. [23]熊庆娥. 植物生理学实验教程[M]. 成都: 四川科技出版社, 2003. [24]叶子飘. 光合作用对光合CO2响应模型的研究进展[J]. 植物生态学报, 2010, 34(6): 727-740. YE Z P. A review on modeling of responses of photosynthesis to light and CO2[J].ChineseJournalofPlantEcology, 2010, 34(6): 727-740. [25]黄宇. 闽楠种子萌发与多胚苗研究[J]. 福建林业科技, 2014, 03: 27-31. HUANG Y.Study on the Seed Germination and Multi-seedlings in Phoebe bournei(Hemsl.) Yang[J].JournalofFujianForestryScienceandTechnology, 2014, 3: 27-31. [26]余道平, 彭启新, 胡庭兴,等. 桢楠多胚现象与多胚苗形态发育的研究[J]. 四川林业科技, 2015, 1: 35-37. YU D P,PENG Q X, HU T X,etal.Research on polyembryony ofPhoebezhennanand Morphogenesis of its polyembryonic seedlingsJournalofSichuan[J].ForestrySciencesandTechnology, 2015, 1: 35-37. [27]周劲松, 罗绍春, 汤泳萍,等. 芦笋种子发芽率、发芽势测定及多胚现象观察[J]. 江西农业学报, 2014, 26(4): 30-32. ZHOU J S,LUO S C,TANG Y P,etal.Studies on germination rate and germination potential of seeds ofAsparagusofficinalisand occurrence of polyembryonic seedlings[J].ActaAgriculturalJiangxi, 2014, 26(4): 30-32. [28]黄雪梅, 杨彩群, 陈之红,等. 黄花风铃木种子活力快速丧失及多胚苗现象[J]. 广东农业科学,2012, 39(12): 50-52. HUANG X M, YANG C Q, CHEN Z H,etal. The rapid loss ofTabebuiachrysanthaseed vigor and occurrence of polyembryonic seedlings[J].GuangdongAgriculturalSciences, 2012, 39(12): 50-52. [29]肖兴翠. 不同营林措施对湿地松林分生长及养分循环的影响研究[D]. 长沙:中南林业科技大学, 2013. [30]勾玲, 闫洁, 韩春丽,等. 氮肥对新疆棉花产量形成期叶片光合特性的调节效应[J]. 植物营养与肥料学报, 2004, 10(5): 488-493 GOU L, YAN J, HAN C L,etal. Effects of nitrogen rates on photosynthetic characteristics and yield of high-yielding cotton in Xinjiang[J].PlantNutritionandFertilizingScience, 2004, 10(5): 488-493. [31]钱燕萍, 祝遵凌. 配方施肥对欧洲鹅耳枥幼苗光合生理的影响[J]. 东北林业大学学报, 2015, 11:32-36. QIAN Y P, ZHU Z L. Effects of formulated fertilization on photosynthetic characteristics ofCarpinusbetulusseedlings[J].JournalofNortheastForestryUniversity, 2015, 11:32-36. [32]袁颖红,樊后保,等. 长期施肥对水稻光合特性及水分利用效率的影响[J]. 生态学杂志, 2009, 11: 2 239-2 244. YUAN Y H,FAN H B,etal. Effects of long term fertilization on rice photosynthetic traits and water use efficiency.[J].ChineseJournalofEcology,2009, 11: 2 239-2 244. [33]王力朋, 晏紫伊, 李吉跃,等. 氮素指数施肥对3个楸树无性系光合特性的影响[J]. 林业科学研究, 2013, 1: 46-51. WANG L P, YAN Z Y, LI J Y,etal. Effects of Nitrogen Exponential Fertilization on Photosynthetic Characteristics of Three Catalpa bungei Clones[J].ForestryResearch, 2013, 1: 46-51. [34]杨自立,马履一,贾忠奎,等. 不同供氮水平对栓皮栎播种苗光响应曲线的影响[J]. 北京林业大学学报,2011,5:56-60. YANG Z L, MA L Y, JIA Z K,etal. Effect of different nitrogen supply on light-response curves ofQuercusvariabilisseedlings[J].JournalofBeijingForestryUniversity, 2011, 5: 56-60. [35]李絮花. 施肥制度与土壤可持续利用[D]. 北京:中国农业科学院, 2005. [36]魏欣. 中国农业面源污染管控研究[D]. 陕西杨陵:西北农林科技大学, 2014. (编辑:裴阿卫) 文章编号:1000-4025(2016)06-1172-10 doi:10.7606/j.issn.1000-4025.2016.06.1172 收稿日期:2016-01-18;修改稿收到日期:2016-05-26 基金项目:国家“十二五”科技支撑计划(2011BAC09B05);四川省“十二五”作物育种攻关项目(2011NZ0098-10) 作者简介:谭 飞(1991-),男,在读硕士研究生,主要从事林木栽培、培育方向的研究。E-mail;635556526qq.com *通信作者:胡红玲,副教授,硕士生导师,主要从事森林培育学相关研究。 E-mail:15039081@qq.com 中图分类号:Q945.79 文献标志码:A Growth and Photosynthesis ofPhoebezhennanPolyembryonic Seedlings under Different Fertilizer Levels TAN Fei, HU Hongling*, HU Tingxing, CHEN Yufeng, LIAO Yuhua,ZHOU Guangliang, LUO Jie,YANG Shanshan (College of Forestry, Sichuan Agricultural University, Chengdu 611130, China) Abstract:The nutrition bag cultivation was used to investigate the effect of fertilization on growth and photosynthetic physiology of machilus(Phoebe zhennan)polyembryonic seedlings. In the study, three types of seedlings i.e. single embryo seedlings(D), bigger one of twin embryo seedlings (SD), and smaller one of twin embryo seedlings (SX) were selected and planted as experimental material. The age of seedlings for planting was 45 d and the rhizome was preliminary lignification. The experiment was set as three levels of fertilizer application, i.e. control (CK, 0 g per bag), medium level (M, 0.2 g per bag) and high level (H, 0.3 g per bag). Compound fertilizer was applied and the amount of fertilizer was divided into 4 parts and began to apply in early July and ended in early September with the interval of 20 d. The determination of seedling growth and physiological indexes of photosynthesis were conducted in mid of August and mid of October. The results showed that: (1) The medium level(M) of fertilization significantly promoted the height and ground diameter growth of three types seedlings, while high level(H) of fertilization showed the effect of promoting first and then inhibition, and indicated the excessive fertilization. (2) The medium level(M) of fertilization significantly promoted the synthesis of chlorophyll of three types of seedlings, prolonged the functional period of leaves, increased net photosynthetic rate and stomatal conductance, and furthermore improve the ability of photosynthesis of the seedlings under high light intensity and the environment of high CO2 concentration. (3) The fertilization can decrease the light compensation point (LCP) and CO2 compensation point (CCP) of seedlings, and increase the light saturation point (LSP) and CO2 saturation point (CSP), promoting seedlings to adapt to the environment and accumulate more organic matter. Whereas the high amount of fertilization on the photosynthetic indicators of seedlings also showed the role of promotion first and then inhibition. The results showed that synthesis of photosynthetic pigment in all machilus seedlings and photosynthetic efficiency as well as environmental adaptability are enhanced under moderate fertilizer application, which helped to promote the growth of seedlings; the medium level (M) of fertilizer application can rapidly reduce the gap between the growth indicators of bigger one of twin embryo seedlings and single embryo seedlings . and also plays a promoting role on the growth of smaller one of twin embryo seedlings. The results indicated the important significance for the conservation of machilus germplasm resources, and for the promotion of qualified rate of seedlings and reduction of the cost of raising seedling. Key words:machilus(Phoebe zhennan); polyembryonic seedling; fertillization; growth; photosynthetic physiology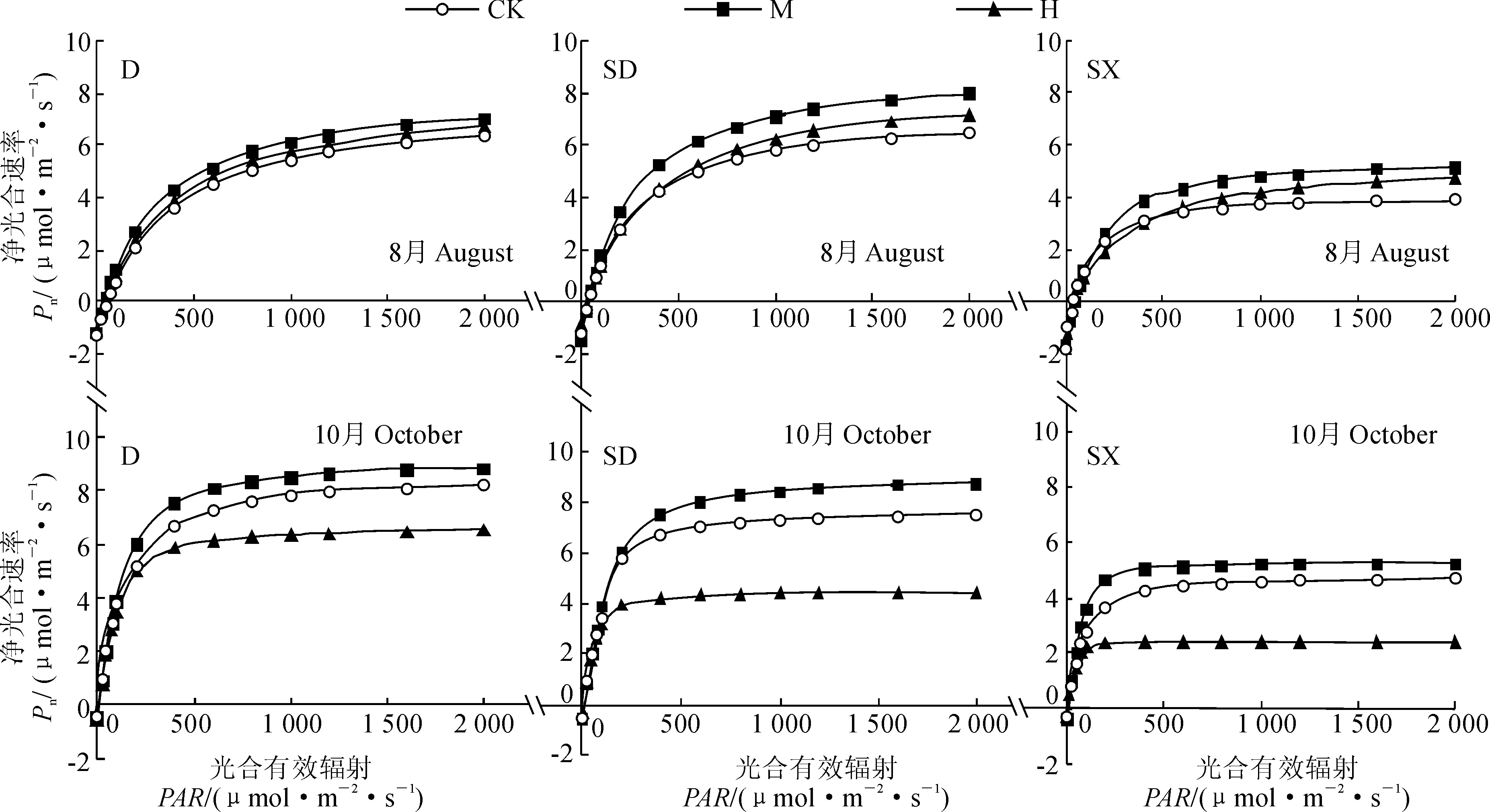

 猜你喜欢
小读者(2021年2期)2021-03-29疯狂英语·新悦读(2019年11期)2019-12-18华人时刊(2019年13期)2019-11-17NBA特刊(2018年21期)2018-11-24文苑(2018年22期)2018-11-19安徽农学通报(2016年24期)2017-01-12吉林农业(2016年12期)2017-01-06安徽农学通报(2016年21期)2016-12-22现代农业科技(2016年20期)2016-12-20红领巾·萌芽(2016年1期)2016-09-10
猜你喜欢
小读者(2021年2期)2021-03-29疯狂英语·新悦读(2019年11期)2019-12-18华人时刊(2019年13期)2019-11-17NBA特刊(2018年21期)2018-11-24文苑(2018年22期)2018-11-19安徽农学通报(2016年24期)2017-01-12吉林农业(2016年12期)2017-01-06安徽农学通报(2016年21期)2016-12-22现代农业科技(2016年20期)2016-12-20红领巾·萌芽(2016年1期)2016-09-10
