
早籼品种湘早籼45号孕穗期淹涝胁迫试验
2016-07-23 18:12宁金花张艳贵解娜
江苏农业科学 2016年5期
宁金花++张艳贵++解娜


摘要:利用湘早籼45号常规水稻品种孕穗期淹涝胁迫数据,分析了不同淹涝深度和时间下水稻形态差异性,并基于不同淹涝深度,建立株高、绿叶数、卷叶长度、高位分蘖、茎长等形态特征值与淹涝持续时间模型。结果表明:相同淹涝深度下,涝渍持续时间与株高变化率、绿叶数变化率、卷叶长度、高位分蘖率、总茎节长度呈二次曲线关系。不同淹涝深度之间淹涝结束后0 d株高变化率差异不大,淹涝结束后5 d差异明显。淹涝结束后0 d,淹涝深度越浅水稻绿叶数变化越明显。1/2淹与2/3淹处理下,淹涝持续时间5 d以内,高位分蘖茎主要发生在倒1茎节;淹涝时间5 d以上,倒1、2茎节均发生高位分蘖的频率较高。随着淹涝深度的增加,倒1、倒3茎节长度呈缩短趋势,倒2茎节呈伸长趋势,总茎节长度呈缩短趋势。
关键词:水稻;淹涝胁迫;株高;绿叶数;卷叶长度;高位分蘖;茎节长度
中图分类号: S511.2+10.3;S422文献标志码: A文章编号:1002-1302(2016)05-0106-06
洪涝是人类面临的最严重的自然灾害之一,在我国南方主要稻区,稻田多分布于丘陵地带的江河谷地、平原湖地,在水稻生产季节常发生洪涝灾害,已成为制约水稻生产的主要生态逆境因子之一[1]。研究水稻在不同淹涝环境下茎、叶形态的变化特点,对于明确不同淹涝环境下水稻形态特征具有深远意义。Middelboe等指出,水淹植物对光的需求与其自身的形态紧密相关,在全淹的环境中,植物为了适应光强减弱,会产生响应,如改变叶片形态[2]。Enríquez等指出,叶片通过改变光学路径来调节植物的吸光效率[3]。Ruiz等指出,植物根据周围的光环境改变叶片形态使其更有利于光合作用的进行[4]。没有节间的水稻幼苗在部分淹水时,主要通过提高顶部2张叶和叶鞘伸长能力完成[5]。部分淹水的深水稻节间伸长,植株长高。在营养生长阶段,淹涝环境对水稻的典型伤害是消耗已积累的干物质,当水稻植株完全淹没,氧、二氧化碳的供应变得有限,水稻植株以无氧呼吸为主,会导致能源急剧消耗[6]。淹涝环境下,由光照减弱造成植株叶片形态的改变包括叶片伸长[7-8]、增宽以及叶面积改变等[9-10]。淹涝胁迫可促进水稻根系木质化、栓质化,减少根中O2径向泄漏,增加根尖O2 浓度[11]。淹涝可引起叶片可溶性糖、淀粉、总糖含量下降[12] 。水稻孕穗期和乳熟期没顶淹水胁迫处理后,孕穗期叶绿素含量明显降低[13-15]。有学者对水稻高位分蘖也进行了相关研究[16]。虽然前人关于淹涝胁迫对水稻影响方面作了很多研究,但都是针对叶片、叶鞘、茎等某一个或几个形态特征因子,或是从生理特性、产量结构进行研究,从绿叶、卷叶、茎、株高等多个形态特征因子,较全面、系统地分析淹涝胁迫对常规稻孕穗期影响研究相对较少。笔者在前人研究的基础上,从茎、叶等主要形态特征着手,探讨不同淹涝胁迫环境下常规稻孕穗期形态变化特点,明确不同淹涝环境下常规稻的适应机制,旨在为长江中下游地区水稻的选种、育种提供依据。
1材料与方法
1.1试验地点
试验地点位于湖南省长沙农业气象试验站(113°05′E,28°12′N,海拔44.9 m),属于亚热带季风性湿润气候,气候温和,降水充沛,雨热同季,四季分明。 每年的4—8 月是该地区的雨季,局地发生淹涝灾害的频率高。供试水稻品种为湘早籼45号 (湘审稻编号:2007002),属常规中熟早籼,在湖南省作双季早稻栽培,全生育期106 d,株高 80~85 cm,叶片厚实挺直,株型松紧适中,茎秆较粗且弹性好,落色好,不落粒。
1.2试验设计
2013年3月29日播种,大田软盘育秧,4月21日移栽于盆中,盆口径30 cm,底径20 cm,高25 cm。移栽时在水泥池中将水、肥、土壤混匀,分别盛入盆中,每盆土层深度约 23 cm。土壤质地为沙壤土,弱酸性,施用水稻专用复合肥,自来水浇灌。每盆移栽秧苗5株,水稻生育期内,根据水稻长势,适时施肥1~2次。
在水稻孕穗期进行不同淹涝深度和淹涝时间胁迫处理,设3个淹涝深度:1/2淹、2/3淹、3/3淹(全淹),设5个淹水时间,分别为2、3、5、7、9 d,每处理设4个重复,共15个处理,即每个淹涝深度处理20盆,每个淹涝时间处理4盆。6月14日(设为淹涝前0 d)开始,将进入孕穗期的水稻植株按照1/2淹、2/3淹、3/3淹方式放入2个淹涝模拟池进行淹水处理,6月16日(2 d)、6月17日(3 d)、6月19日(5 d)、6月21日(7 d)、6月23日(9 d)陆续取出,放入自然条件下至7月26日相继成熟收获,8月15日试验结束。淹涝深度由预先制作的铁床控制,全淹的20盆直接放入模拟池中,1/2淹的20盆放在模拟池内可以移动的铁床上。2/3淹放入另一模拟池。对不同淹涝深度、不同淹水时间的水稻分别挂牌,顺序放入,达到不同的淹涝时间后取出,置于自然条件下,直至成熟、收获。以整个生育期不受淹处理植株为对照,设4个重复,对照植株移栽至成熟,在自然环境下与处理水稻进行平行观测,直至收获、成熟。
模拟池为人工砌的水泥池,全淹水泥池深度1.3 m,长3.6 m,宽约2.7 m;2/3淹水泥池深度0.9 m,长4.5 m,宽2.7 m。人工定做2个不同高度的铁床,用以控制淹涝深度。全淹为没顶淹没,1/2、2/3淹的深度以所有处理的平均株高(叶高)为基准,淹平均株高的1/2、2/3。
1.3观测项目
对供试品种淹水前后株高、绿叶数、卷叶长度、高位分蘖、茎节长度等形态性状进行观测。绿叶数:在淹水前和取出后对每盆中所有茎上的绿叶数进行统计,按照农业气象相关规范,能进行光合作用的叶片均为绿叶,没展开的新叶也一并计入。部分黄化的叶片则按绿色部分占整个叶片的百分比进行计算,为目测估算值,如绿色部分面积占整个叶片面积的30%,其余部分已变黄,则记为0.3张绿叶,结果保留一位小数。对对照盆中的绿叶数进行平行观测、统计。卷叶长度:观测不同淹涝胁迫处理后倒1叶的卷叶长度,四舍五入取整数。绿叶变化率、高位分蘖率计算公式如下:
绿叶变化率=淹涝结束后N天的绿叶数-前一次的绿叶数前一次的绿叶数×100%;(1)
高位分蘖率=淹涝处理后发生高位分蘖的茎数总茎数×100%。(2)
1.4数据分析
采用Excel 2010以及DPS数据处理系统整理分析数据,采用单因素方差分析和Duncans新复极差检验比较不同处理之间的差异,采用相关分析法检验不同淹涝深度、时间与各观测项目间的相关关系。
2淹涝胁迫对水稻形态特征的影响
2.1淹涝胁迫对水稻株高的影响
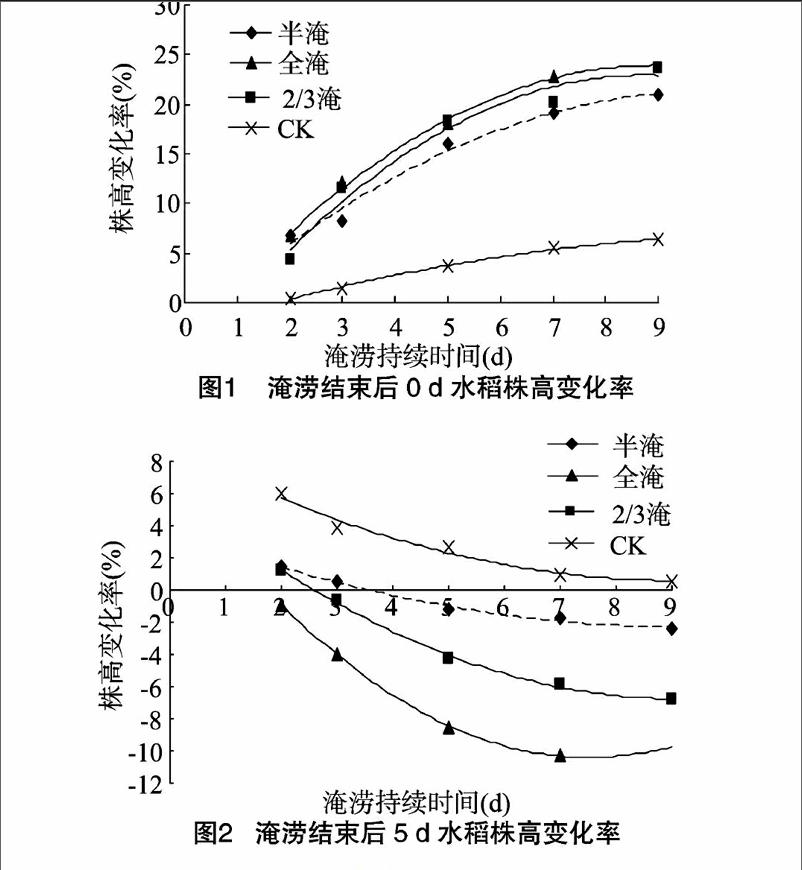
分别观测淹涝前0 d以及淹涝结束后0、5 d的植株株高,并计算株高变化率。由图1、图2可知,对照与不同深度处理的水稻植株淹涝结束后0、5 d的株高变化率变化规律不同,淹涝结束后 0 d 水稻株高变化率随着淹涝持续时间的延长呈增加趋势。淹涝结束后5 d水稻株高随淹涝持续时间的延长呈减少趋势。涝渍持续时间与株高变化率呈二次曲线关系。二次曲线方程及非线性相关系数及曲线方程的极值点见表1。由表1可知,淹涝结束后 0 d 株高变化率存在极大值点,淹涝结束后5 d株高变化率存在极小值点。极大指点反映了植株在淹涝过程中的耐淹能力,由不同淹涝深度曲线的极值点大小可知,全淹处理下植株耐淹能力最差,其次是2/3淹。淹涝结束后5 d的株高变化率反映了水稻遭受淹涝胁迫后的恢复能力,从极值点大小可知,1/2淹、2/3淹与对照差别不大,均是9.0 d左右时最小,全淹是7.5 d,由此可知,全淹涝渍持续时间超过7.5 d,对水稻植株后期植株株高的影响是致命的。淹涝结束后0 d不同深度之间株高变化率差异不大;淹涝结束后5 d,不同深度的淹涝处理之间株高变化率差异明显。相同淹涝持续时间,均是全淹处理株高变化率的绝对值最大。
2.2淹涝胁迫对水稻绿叶数的影响
由图3、图4可知,与对照相比,淹涝结束后0 d水稻绿叶数变化率均与对照变化率规律相反,淹涝结束后5 d与水稻绿叶数变化率均与对照变化率规律基本一致。由表2可知,不同淹涝深度下,水稻绿叶数变化率与淹涝持续时间呈二次曲线关系。从二次曲线方程的极大值点可知,淹涝深度越深,表1淹涝持续时间与水稻株高变化率间的关系淹涝水稻叶片对环境的反应越敏感,绿叶数变化率越大。淹涝结束后0 d水稻绿叶数变化率反映了淹涝过程中水稻植株的耐淹能力,相同淹涝持续时间,淹涝深度越浅,绿叶数变化越明显。淹涝结束后5 d水稻绿叶数变化率反映了水稻植株的自我恢复能力,从曲线的变化趋势可知,5 d后的绿叶数变化率存在极小值点。意味着水稻植株遭受淹涝灾害胁迫后,绿叶数不会一味减少,当指数绿叶数减少到一定程度后,植株的自我恢复能力被迫增强,以维持自身后期的生长发育。
2.3淹涝胁迫对卷叶长度的影响
分别观测淹涝前0 d,淹涝后0、5 d的植株卷叶长度,并计算其变化率,见图5、图6。从图5中可知,和对照相比,淹涝结束后0 d的卷叶长度变化率,三个不同深度处理的水稻植株卷叶长度在淹涝在淹涝过程中有一个由增长至减少的变化过程,即淹涝过程中水稻的卷叶长度存在极大值点,但不同淹涝深度出现极大值点的时间不同。淹涝持续过程中,2/3淹、全淹的卷叶长度最大值出现在淹涝5 d,1/2淹出现在 3 d。淹涝结束后5 d,3个淹涝处理的卷叶长度变化率变化规律与对照一致,卷叶长度变化率均与淹涝持续时间呈开口向上的抛物线关系,均存在一个极小值点,但极值点大小不同。淹涝结束后0 d卷叶长度变化率反映了植株在淹涝过程中水稻叶片的适应机制。由图5可知,淹涝时间在一定范围内,水稻植株通过卷叶来维持生命,超过特定时间,即极值点时间,卷叶长度不再延长,水稻采取其他适应方式来维持生命。淹涝结束后5 d水稻卷叶长度变化率反映了植株叶片淹涝结束后适应机制。淹涝过程中卷叶越明显的水稻植株,淹涝结束后植株展开也越明显(表3)。
2.4淹涝胁迫对水稻茎的影响
2.4.1淹涝胁迫对水稻高位分蘖的影响由表4可知,孕穗期水稻遭受不同深度、不同持续时间的淹涝胁迫影响后,不同茎节发生高位分蘖的情况差异较大。同一淹涝深度,1/2淹与2/3淹淹涝持续时间2~5 d,高位分蘖茎节主要发生在倒1茎节,淹涝持续时间5 d以上,倒1、2茎节均发生高位分蘖的频率较高。全淹处理遭受淹涝胁迫后,倒1、2 茎节均发生高位分蘖的几率较高。同一淹涝深度总高位分蘖率与淹涝持续时间呈二次曲线关系,分别对二次曲线方程求拐点、极值可知,淹涝持续时间为5.6 d时1/2淹的高位分蘖率最高,最高为63.6%;2/3淹为6.5 d时高位分蘖率最大,为78.8%;全淹为4.4 d时最高,为56.5%。3个淹涝深度,淹涝持续时间2 d以上,2/3淹高位分蘖现象最明显,其次是1/2淹(图7)。不同淹涝深度、不同淹涝持续时间的总高位分蘖率均与对照差异极显著。3个不同淹涝深度,淹涝持续时间7 d或以上,倒1、2茎节同时发生高位分蘖的概率高。
2.4.2淹涝胁迫与茎节长度由表5可知,遭受不同深度、不同淹涝时间胁迫,水稻不同茎节长度发生很大变化。随着淹涝深度的增加,倒1、3茎节长度呈缩短趋势,倒2茎节呈伸长趋势,但总的茎节长度亦呈减少趋势。和对照相比,3个淹涝深度处理均表现为倒1、3茎节长度缩短,倒2茎节伸长。
由图8可知,同一淹涝深度,总茎节长度与淹涝持续时间呈二次曲线关系,非线性相关系数分别为:0.9837(1/2淹)、0.992 7(2/3淹)、0.999 9(全淹),非线性相关系数均通过0.01水平显著性检验,由此可知,不同淹涝深度下,水稻总茎节长度与淹涝持续时间呈极显著相关关系。同一淹涝深度,总茎节长度随着淹涝持续时间的延长而增大,对曲线方程求拐点、极值可知,1/2淹淹涝持续时间10.1 d时茎节长度最长,2/3淹为17.7 d时最长,全淹为7.3 d时最长。由此可知,在不同淹涝深度下,水稻茎节均采用伸长生长的“逃逸策略”来适应环境。不同淹涝深度下,随着淹涝持续时间的延长,茎节的伸长生长存在一个极值点。
同一淹涝深度,不同茎节应对淹涝胁迫环境的反应不同,直接表现为倒1、倒2、倒3茎茎节长度不同。同一淹涝深度下,同一茎节长度与淹涝持续时间呈二次曲线关系(曲线方程略)。同一茎节在不同淹涝深度下表现不同。倒1茎节长度最长时,1/2淹淹涝时间持续为6.7 d,2/3淹为7.1 d,全淹为6.0 d。倒2茎节长度最长时1/2淹淹涝持续时间为8.4 d,2/3淹为12.8 d,全淹为6.6 d。倒3茎节长度最小时,1/2淹淹涝持续时间为3.5 d,2/3淹为1.1 d,全淹为2.2 d。
3结论与讨论
3.1株高与淹涝胁迫
植株高度在农业气象观测上是评价作物不同生育期生长状况、品种差异等的重要因子。本试验从株高的变化探讨孕穗期水稻对淹涝环境的敏感性。本研究结果表明,不同淹涝深度下,涝渍持续时间与株高变化率呈二次曲线关系。不同淹涝深度下淹涝0 d株高变化率差异不大,淹涝5 d株高变化率差异明显。
3.2叶片与淹涝胁迫
本试验结果表明,不同淹涝深度下,淹涝结束后0 d的绿叶数变化率的变化规律与对照相反,淹涝结束后5 d与对照基本一致。绿叶数变化率与淹涝持续时间呈二次曲线关系。淹涝结束后0 d,相同淹涝持续时间下,淹涝深度越浅绿叶数变化越明显。淹涝结束后5 d绿叶数变化率存在极小值,绿叶减少到一定程度后,植株的自我恢复能力被迫增强,以维持自身后期的生长发育。由此可见,水稻的耐淹能力与碳水化合物多少及淹涝环境有很大关系。研究指出,植物耐淹能力与淹没前的碳水化合物含量成正相关[17-18]。虽然本试验对不同淹水深度和持续时间下孕穗期水稻绿叶的变化特点进行了探讨,但对该生育期绿叶变化与产量之间的定量关系没有深入探讨,有必要进行更深入的研究。
淹水条件下,植物体生长减慢,叶片数、叶面积减少,同时部分叶片老化、脱落,导致叶片生物量的急剧下降[19]。对于淹涝胁迫环境下叶片的卷叶情况,前人没有多做探讨,但是将叶片卷曲度作为鉴定水稻抗旱能力的指标在育种和栽培上己有应用[20-22]。本试验结果表明,卷叶是不同淹涝环境下孕穗期水稻适应淹涝胁迫环境的一种方式。淹涝环境下,卷叶导致水稻叶面积明显减少,这和前人的研究结果相同。茅弼华等研究指出,淹没6 d,单茎上部3张叶的叶面积为正常稻株的72.2%,淹没10 d的叶面积仅及正常稻株的50.3%[23]。另外,水淹植物对光的需求与其自身的形态紧密相关[24]。在淹涝环境中,为适应光强减弱的环境,植物会产生响应,如根据周围的光环境改变叶片形态,使其更有利于光合作用的进行[25]。由光照减弱造成叶片形态的改变包括叶片的伸长、增宽以及叶面积的改变等[10]。
3.3茎与淹涝胁迫
水稻的孕穗期是水稻生长发育的关键期,该时期遭受淹涝胁迫会致使稻穗不生长或者花粉发育受阻,甚至幼穗颖花与枝梗严重退化,致使植株体内的营养物质发生转移,致使淹涝胁迫后高位分孽大量发生。因高位分蘖的发生,导致分蘖茎与主茎之间、分蘖茎与分蘖茎之间、老茎与新茎之间、分蘖茎与新茎之间的发育不一致,另外遭受淹水处理的水稻植株之间以及处理水稻与对照之间生长发育进程不一致。淹涝胁迫结束后,无法齐穗,直接影响水稻的抽穗及灌浆成熟。对于高位分蘖的原因,有研究人员从气温、光照、水分等方面进行了阐述并提出预防措施。也有人认为,水稻叶腋中有腋芽,条件适宜时能发育为分蘖[26]。还有学者从基因方面对高位分蘖进行了描述,从常规粳稻中筛选到一份自然突变的抽穗期延迟的类树稻突变体lhd3(leafy head 3),该突变体在生长后期,上部节间会继续长出叶片(一般为3张)和高位分蘖,类似于树的侧枝生长,抽穗期延迟,但基部分蘖数不受影响[27]。
虽然高位分蘖使单位面积穗数有所增加,但二次分蘖穗小粒少、生长期延长、产量不高。对长江流域双季稻区早稻而言,由于早稻季节性强,生育时期要求很严,受淹早稻长出的高位芽会明显延长生育期,影响晚稻播种及时移栽。因此,控制受淹早稻的高位芽是早稻耐涝减灾的主要措施之一[28]。茎的变化是水稻生长发育过程中一个非常重要的方面[28-30]。水稻后期株高的变化主要表现为茎的长短,特别是遭受淹涝胁迫的水稻植株。本研究结果表明,随着淹涝深度的增加,倒1、倒3茎节长度呈缩短趋势,倒2茎节呈伸长趋势,但总的茎节长度呈减少趋势。同一淹涝深度,总茎节长度与淹涝持续时间呈二次曲线关系,同一茎节长度与淹涝持续时间呈二次曲线关系。不同茎节应对淹涝胁迫环境的反应不同,直接表现为倒1、倒2、倒3茎茎节长度不同。
参考文献:
[1]李阳生,李绍清. 湖南农业的洪涝灾害问题及对策[J]. 农业现代化研究,1998,19(2):39-42.
[2]Middelboe A L,Markager S. Depth limits and minimum light requirements of freshwater macrophytes[J]. Freshwater Biology,1997,137:553-568.
[3]Enríquez S,Pantoja-Reyes N I. Form-function analysis of the effect of canopy morphology on leaf self-shading in the seagrass Thalassia testudinum[J]. Oecologia,2005,145(2):235-243.
[4]Ruiz J M,Romero J.Effects of in situ experimental shading on the Mediterranean sea-grass Posidonia oceanica[J]. Marine Ecology Progress Series,2001,215:107-120.
[5]Khan A A,Thakur R,Akbar M,et al. Relation of ethylene production to elongation in deep-water rice[J]. Crop Sci,1987,27:1188-1196.
[6]Setter T L,Ellis M,Laureles E V,et al. Physiology and genetics of submergence tolerance in rice[J]. Annals of Botany,1997,79(A):67-77.
[7]Gordon D M,Grey K A,Chase S C,et al. Changes to the structure and productivity of a Posidonia sinuosa meadow during and after imposed shading[J]. Aquatic Botany,1994,47(3/4):265-275.
[8]罗文泊,谢永宏,宋凤斌.洪水条件下湿地植物的生存策略[J]. 生态学杂志,2007,26(9):1478-1485.
[9]Lee K S,Dunton K H. Effect of in situ light reduction on the maintenance,growth and partitioning of carbon resources in Thalassia testudinum Banks ex konig[J]. Journal of Experimental Marine Biology and Ecology,1997,210:53-73.
[10]Campbell S J,Miller C J. Shoot and abundance characteristics of the seagrass Heterozostera tasmanica in Westernport estuary (south-eastern Australia)[J]. Aquatic Botany,2002,73:33-46.
[11]李玉昌,李阳生,李绍清.淹涝胁迫对水稻生长发育危害与耐淹性机理研究的进展[J]. 中国水稻科学,1998,12(增刊1):70-76.
[12]李乐农,麻浩,李继军,等. 淹水处理对不同水稻品种碳氮代谢的影响[J]. 湖南农学院学报,1995,21(3):235-238.
[13]李阳生,李绍清.淹涝胁迫对水稻生育后期的生理特性和产量性状的影响[J]. 武汉植物学研究,2000,18(2):117-122.
[14]Middelboe A L,Markager S. Depth limits and minimum light requirements of freshwater macrophytes[J]. Freshwater Biology,1997,37(3):553-568.
[15]李绍清,李阳生,李达模.乳熟期淹水对两系杂交水稻源库特性的影响[J]. 杂交水稻,2000,15(2):38-40,44.
[16]莫永.水稻高位分蘖发生的原因与预防控制[J]. 种子科技,2010,28(12):33-34.
[17]Singh H P,Singh B B,Ram P C. Submergence tolerance of rainfed lowland rice:search for physiological marker traits[J]. Journal of Plant Physiology,2001,158(7):883-889.
[18]Ram P C,Singh B B,Singh A K,et al. Physiological basis of submergence tolerance in rain-fed lowland rice:prospects for germplasm improvement through marker aided breeding[J]. Field Crop Research,2002,76(2/3):131-152.
[19]卓仁英,陈益泰.木本植物抗涝性研究进展[J]. 林业科学研究,2001,14(2):214-222.
[20]Kawano N I O. Physiological responses of rice seedlings to complete submergence(flash flooding)[J]. Annals of Botany,2009,103(2):161-169.
[21]OToole J C,Cruz R T. Response of leaf water potential,stomatal resistance,and leaf rolling to water stress[J]. Plant Physiology,1980,65(3):428-432.
[22]Turner N C,OToole J C,Cruz R T,et al. Responses of seven diverse rice cultivars to water deficits Ⅰ. Stress development,canopy temperature,leaf rolling and growth[J]. Field Crops Research,1986,13:257-271.
[23]茅弼华,王荫传,陆仁峥. 淹涝水稻生长特点与栽培技术[J]. 江苏农业科学,1992(3):1-5.
[24]Middelboe A L,Markager S.Depth limits and minimum light requirements of freshwater macrophytes[J]. Freshwater Biology,1997,137:553-568.
[25]Ruiz J M,Romero J. Effects of in situ experimental shading on the Mediterranean seagrass Posidonia oceanica[J]. Marine Ecology Progress Series,2001,215:107-120.
[26]Nagasawa N,Miyoshi M,Sano Y,et al. DL regulates both leaf and pistil development in rice[J]. Rice Genetics Newsletter,1996,13:102-105.
[27]吴立文,姜华,胡江,等. 一个水稻类树稻突变体(leafy head 3)基因的精细定位[J]. 中国水稻科学,2009,23(6):583-588.
[28]吕艳东,郭晓红,李红字,等. 肥水耦合对寒地水稻产量的影响[J]. 江苏农业科学,2015,43(11):93-97.
[29]李世峰,刘蓉蓉,吴九林.膜下滴灌水稻不同穗肥运筹处理对水稻产量的影响[J]. 江苏农业科学,2015,43(11):97-99.
[30]马孟莉,孟衡玲,雷恩,等. 铬(Ⅲ)对不同水稻品种种子萌发及幼苗生长的影响[J]. 江苏农业科学,2015,43(7):74-76.李少朋,毕银丽 ,彭星. 接种丛枝菌根真菌对矿井水回灌玉米生长的影响[J]. 江苏农业科学,2016,44(5):112-115.
猜你喜欢
今日农业(2021年21期)2021-11-26
军事文摘(2021年22期)2021-11-26
今日农业(2021年14期)2021-10-14
农村百事通(2019年17期)2019-10-08
现代农业科技(2017年1期)2017-03-06
