
生物炭对镉胁迫下水稻生长及光合产量的影响
2016-07-23 18:07张学艳曹莹孟军
江苏农业科学 2016年5期
张学艳++曹莹++孟军
摘要:选用辽宁地区水稻主推品种沈农9816,采用盆栽试验,研究了生物炭对镉胁迫下水稻农艺性状以及光合产量的影响。结果表明,水稻农艺性状以及产量指标均呈现Cd1(1 mg/kg)>Cd2(2 mg/kg)>Cd0(0 mg/kg)>Cd4(4 mg/kg)的趋势,添加生物炭后,水稻的生物量、光合参数、产量等指标均呈现C21(4 t/hm2 )>C0(0 t/hm2 )>C210(40 t/hm2 )的趋势。另外,在各镉处理下,添加生物炭C21、C210与C0相比,C21处理各指标增幅均大于对照,且在Cd2浓度下增幅最大。说明生物炭可减轻镉胁迫、促进水稻生长以及提高光合作用,并且最佳用量为C21处理,在Cd2浓度下效果最好。
关键词:生物炭;镉胁迫;光合;产量;水稻;生长
中图分类号: S511.01文献标志码: A文章编号:1002-1302(2016)05-0097-05
随着现代工农业的不断发展,重金属尤其是土壤重金属污染日趋严重,据统计,我国已有超过28万hm2的农田遭受重金属污染[1]。水稻作为全球尤其是亚洲最主要的粮食作物,为65%的中国人口提供主食[2]。镉作为“五毒之首”[3],是稻田中最主要的重金属污染元素[4],它能够通过植物根系吸收,在植物体内产生富集,并通过食物链直接威胁食品生产安全以及人类健康[5-6]。生物炭具有较大的比表面积,较强的强吸附力、抗微生物和化学分解能力,可以在土壤中存留更长时间,为植物提供养分[7-8]。此外,生物炭可以降低重金属的生物有效性,对重金属起到很好的固定效果,减少农作物对重金属的吸收,降低毒害作用[9],成为研究土壤重金属污染改良的热点。近年来,生物炭被认为是解决全球性环境、气候、粮食问题的有效途径,引起了国际国内的普遍重视[10-11]。在我国,以生物炭为核心的秸秆炭化还田不仅可以有效利用生物质资源,增加土壤输入,而且对于保障粮食有效供给尤其具有重要意义[12]。生物炭取之于农,用之于农,在治理重金属污染、改良土壤方面展现出巨大的应用价值。目前,利用生物炭来缓解镉对水稻胁迫影响的研究报道尚不多见,因此,本试验以水稻为研究对象,探讨不同用量生物炭对重金属镉胁迫下水稻生长及光合产量的影响,旨在探明减轻镉胁迫、促进水稻生长以及提高光合作用的最佳生物炭用量。
1材料与方法
1.1试验材料
选用辽宁地区主推水稻品种沈农9816,生物炭由玉米秸秆350 ℃低温厌氧烧制而成,其中镉含量为0.073 mg/kg,pH值为9.23,全氮8.73 mg/kg,全磷8.6 mg/kg,全钾 48.7 mg/kg,硫2.97 mg/kg。土壤采用农田土,棕壤,有机质含量1.18%,碱解氮含量107.33 mg/kg,速效磷含量 34.52 mg/kg,速效钾含量98.41 mg/kg,全镉0.15 mg/kg,有效镉0.09 mg/kg,pH值5.9,阳离子交换量17.21 cmol/kg。
1.2试验设计
采用盆栽试验,在沈阳农业大学北山试验基地进行。试验设有2个处理因素:镉和生物炭。镉试验浓度0、1、2、4 mg/kg,记为Cd0、Cd1、Cd2、Cd4,生物炭添加量:0、4、40 t/hm2,折合每盆炭施用量为0、21、210 g,记为C0、C21、C210。盆栽用土风干过筛,装入聚乙烯桶(高30 cm,直径28 cm),每盆装风干土12.75 kg,于秧苗移栽前1个月将Cd以CdCl2·2.5H2O的形态与土混合,保持盆内水层,使土壤与Cd进行平衡。每盆栽3穴,每穴2苗,各处理12盆,一共144盆,肥料施入量(按土壤质量计算)分别为:尿素0.428 g/kg、CaHPO4·2H2O 0.523 g/kg、K2SO4 0.247 g/kg,于分蘖期每盆追施尿素0.118 g/kg,抽穗期每盆追施尿素0063 g/kg,其余正常管理。取样时期:分蘖期、灌浆期和蜡熟期;成熟期考种测产。在水稻整个生育期,保持水肥条件一致,及时防治病虫草害,保证水稻正常生长发育。
1.3测定项目与方法
1.3.1生物量测定在各取样时期,采用常规方法测量水稻的株高、分蘖数;采用加拿大产WinRHIZO根系分析系统测量总根长、总根体积、总根表面积;取各器官鲜样在105 ℃下杀青30 min,75 ℃烘干至恒质量后用百分之一的电子天平测各部分的干质量。
1.3.2叶绿素取剑叶去掉叶脉,剪碎,称0.04 g,加入10 mL 95%乙醇避光浸泡48 h,测前摇匀,用分光光度计测定,按以下公式计算叶绿素含量:Ca=13.95D665 nm-688D649 nm;Cb=24.96D649 nm-7.32D665 nm;Wa=(Ca·V·k)/m;Wb=(Cb·V·k)/m;Wc=Wa+Wb。式中:Ca、Cb分别为叶绿素a和叶绿素b的浓度;Wa、Wb 、Wc分别为叶绿素a、叶绿素b以及总叶绿素含量;V为提取液体积;k为稀释倍数;m为样品鲜质量。
1.3.3叶面积采用Epson Expression 11000XL叶面积扫描系统测量。
1.3.4光合指标采用CIRAS-3光合仪(美国PP Systems 公司)在晴朗的上午,09:00—11:00,测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等指标。选择功能叶为长势一致的水稻剑叶进行测定,每次测量均为叶片中部(正面)。
1.3.5产量指标成熟期每处理取3盆水稻考种,测定每盆穗数、穗粒数、结实率、千粒质量、每盆产量等指标,采用电子分析天平(感量0.001 g)称质量。
1.3.6数据处理采用Excel 2010、SPSS 17.0对数据进行统计分析。
2结果与分析
2.1生物炭对镉胁迫下水稻生长的影响
由表1可知,生物炭添加下,随镉浓度的增大,水稻株高、分蘖数、地上干质量、地下干质量、总根长、总根体积、总根表面积整体上均呈现先增加后降低的趋势,且均在Cd1浓度达到最大值,呈现Cd1>Cd2>Cd0>Cd4的趋势,与空白组趋势一致。
相同镉浓度条件下,随着生物炭用量的增加,除了总根长、总根体积、总根表面积在分蘖期呈现C210>C21>C0的趋势,其余各农艺性状在整个生育时期均呈现先增加后降低的趋势,C21处理效果最好(表1)。
在不同镉浓度处理下,添加生物炭即C21、C210与C0相比,C21处理各农艺性状均有所增加,C210有所降低,说明C21处理对水稻生长的促进作用更明显。在分蘖期,C21与C210相比,2种生物炭添加量均在Cd4处理增幅最大,且C21各指标增幅更大,C210除在分蘖数、地上干质量比对照有所降低外,其余指标均增加,说明生物炭在水稻生长前期对高浓度镉处理效果好,且C21处理比C210处理的促进作用更明显。添加 C21处理后,株高、地上干质量、地下干质量均在 Cd1浓度达到最大增幅,根系形态指标在Cd2浓度达到最大增幅,分蘖数在Cd4浓度达到最大增幅。蜡熟期,C21处理均比对照有所增加,株高、分蘖数、总根长、总根体积均在Cd1浓度达到最大增幅,地上干质量、总根表面积均在Cd4浓度达到最大增幅,地下干质量在Cd2浓度达到最大增幅,说明在水稻的生长后期,添加生物炭对各农艺性状来看,增幅不如生长前期明显(表1)。2.2生物炭对镉胁迫下水稻叶绿素及光合能力的影响
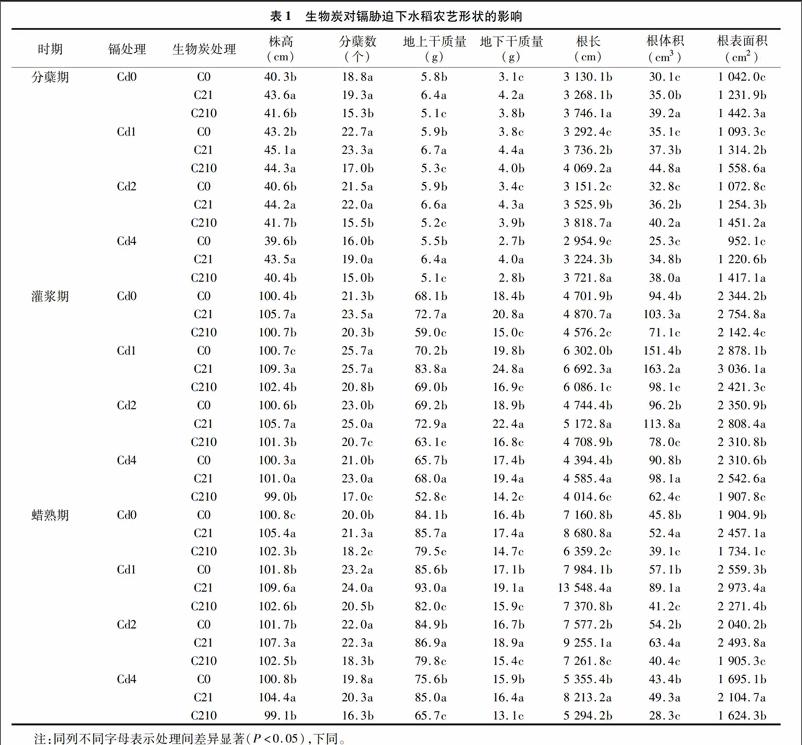
2.2.1对叶绿素含量的影响图1显示,同一炭量添加下,叶绿素总含量随着镉浓度的增加先增加后降低,即Cd1>Cd2>Cd0>Cd4。同一镉浓度添加条件下,叶绿素总含量随着生物炭量的增加呈现C21>C0>C210的趋势,且达到差异显著水平(P<0.05)。在Cd1浓度下,C21在3个生育时期分别比对照高2.17%、1.60%、3.04%;在Cd2浓度下,C21在3个生育时期分别比对照高2.60%、6.89%、4.59%;在Cd4浓度下,C21在3个生育时期分别比对照高2.20%、5.15%、3.41%,且C210均比对照低,说明适量的生物炭(C21处理)对水稻体内叶绿素的形成具有促进作用,而过量的生物炭(C210处理)会产生抑制作用,且C21处理在Cd2浓度下效果最好。在水稻的整个生育时期,叶绿素总含量呈现逐渐降低的趋势,即分蘖期>灌浆期>蜡熟期,说明随着水稻的逐渐衰老,叶片失绿变黄,体内叶绿素总含量也随之降低。
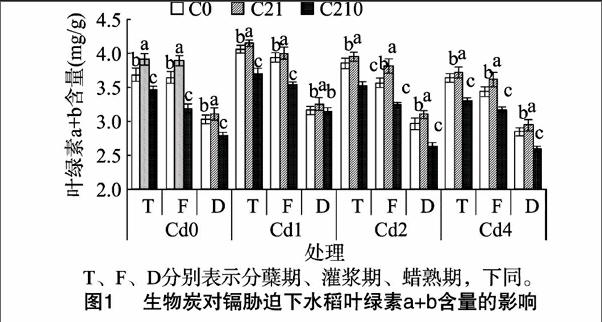
2.2.2对叶面积的影响图2显示,在无生物炭添加条件下,随着镉浓度的增加,水稻分蘖期叶面积变化趋势不显著(P>0.05),相同炭量施加条件下,随着镉浓度的增加,叶面积呈现升高后降低的趋势,即Cd1>Cd2>Cd0>Cd4。在相同镉浓度处理下,叶面积随着生物炭量的增加呈现先增加后降低的趋势,C21处理的叶面积达到最大值,C210处理达到最小值,且达到差异显著水平(P<0.05),这与水稻的农艺性状趋势吻合。在整个生育时期,各处理组均呈现灌浆期>蜡熟期>分蘖期的趋势,这与生长前期叶片数量少且小、生长后期叶片衰老死亡有关。
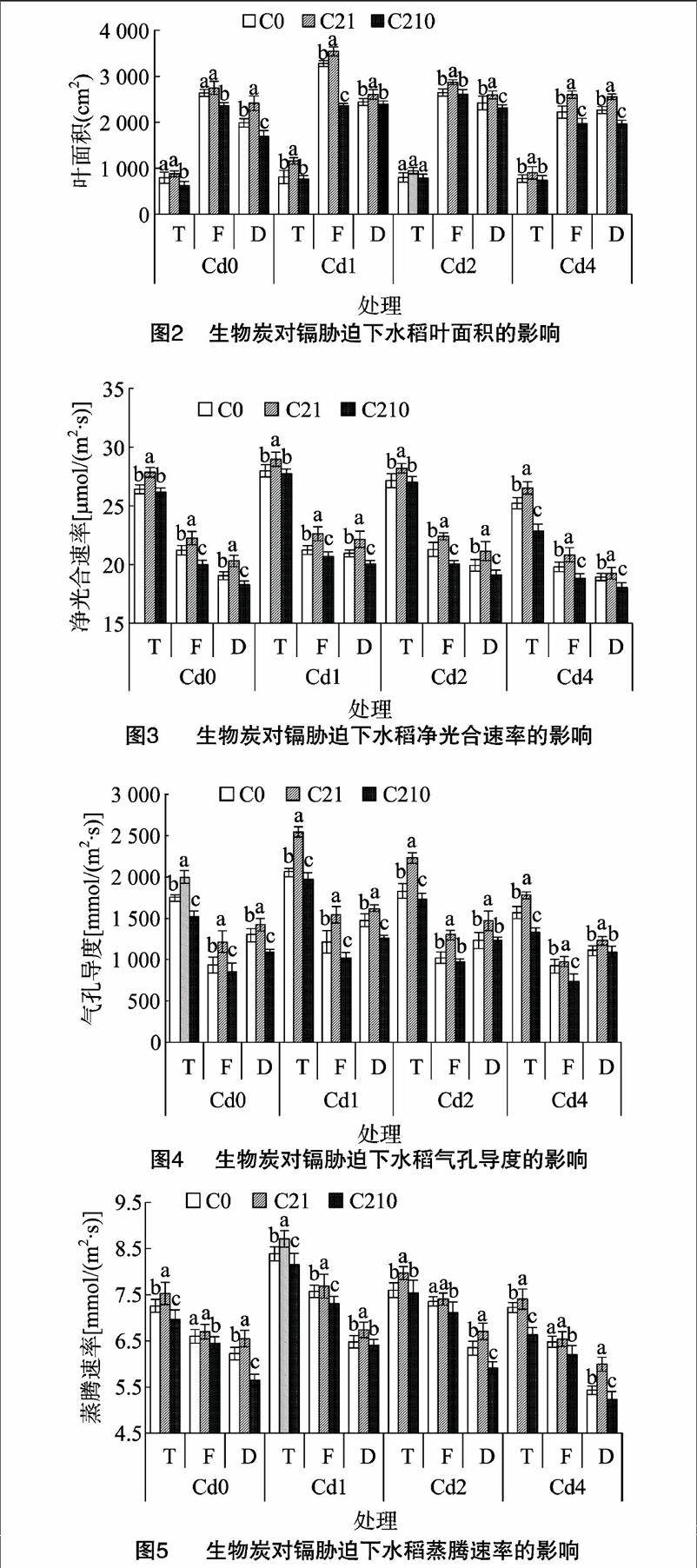
2.2.3对光合能力的影响图3至图6显示,同一生物炭量添加下,水稻净光合速率、气孔导度、蒸腾速率、胞间CO2浓度随着镉浓度的增加呈现先升高后降低的趋势。同一镉浓度条件下,水稻净光合速率、气孔导度、蒸腾速率、胞间CO2浓度随着生物炭量的增加呈现先增加后降低的趋势,说明不管是否有镉胁迫作用,添加生物炭都可以促进水稻净光合速率、气孔导度、蒸腾速率、胞间CO2浓度的增加,其中C21处理的效果最好,C210处理光合能力最弱,各处理达到差异显著水平(P<0.05)。在水稻的整个生长期,水稻的净光合速率、蒸腾速率均在分蘖期达到最大值,其次为灌浆期,最后为蜡熟期,这是因为随着水稻生长期的不断推移,叶片开始衰老,导致光合能力也随之减弱;气孔导度在分蘖期达到最大值,灌浆期达到最小值,蜡熟期又有所回升;胞间CO2浓度在蜡熟期达到最大值。
2.3生物炭对镉胁迫下水稻产量的影响
降低,Cd1浓度达到最大值,Cd4浓度达到最小值。在相同镉浓度处理下,随着炭量的增加,每盆穗数、穗粒数、结实率、每盆产量均随呈现先增加后降低的趋势,其中C21处理表现最好,其次是C0处理,即C21>C0>C210,且各处理达到差异显著水平(P<0.05)。添加生物炭C21、C210与C0对比发现,在各产量指标中,C21处理的增幅均大于C210,在C21处理下,每盆穗数、结实率、产量等指标均在Cd2浓度下增幅最
大,穗粒数、千粒质量在Cd4浓度下增幅最大。在各处理中,C21-Cd1处理每盆产量达到最大值,C210-Cd4处理达到最小值。在各镉处理组,C21处理产量分别比对照高7.21%、806%、7.48%,C210处理产量分别比对照低10.42%、928%、10.41%,由此说明,添加适量的生物炭可以促进水稻产量的增加,过量的生物炭反而会导致水稻减产,并且在镉胁迫下,C21增产效果在Cd2浓度最好,这与水稻农艺性状在生育后期Cd2浓度增幅效果最好相吻合。
2.4水稻叶绿素、叶面积、光合特性与产量之间的相关性
通过相关性分析(表3),叶绿素总含量与叶面积、蒸腾速率呈显著正相关(P<0.05),与净光合速率、气孔导度、胞间CO2浓度、产量呈极显著正相关(P<0.01)。叶面积与净光合速率、气孔导度、蒸腾速率呈显著正相关(P<0.05),与胞间CO2浓度在分蘖期、灌浆期呈显著正相关(P<0.05),与产量呈极显著正相关(P<0.01)。净光合速率与气孔导度、胞间CO2浓度、产量呈极显著正相关(P<0.01),与蒸腾速率呈显著正相关(P<0.05)。气孔导度与蒸腾速率、胞间CO2浓度、产量呈极显著正相关(P<0.01)。蒸腾速率与胞间CO2浓度在分蘖期、蜡熟期呈显著正相关(P<0.05),与产量呈极显著正相关(P<0.01)。胞间CO2浓度与产量呈极显著正相关(P<0.01)。
3讨论与结论
株高、根系形态以及生物量等性状指标常被用作评价作物重金属耐性的重要指标[13]。本研究表明,同一炭量添加下,随着镉浓度的增加,水稻的株高、分蘖数、生物量以及总根长、总根体积、总根表面积会随之先升高后降低,这与“少促多抑”的研究结果相同[14]。同一镉浓度条件下,在水稻的整个生育时期,适量的生物炭(C21处理)可以促进水稻的生长发育,过量的生物炭(C210处理)在水稻生长后期有抑制作用,张晗芝等对玉米苗期的研究也表明,过量的生物炭(48 t/hm2) 对玉米的株高、生物量有显著抑制作用[15]。根系作为植物主要的吸收器官,其生长和分布直接影响植物对重金属的吸收能力和吸收效率,根系的变化也直接反映了重金属对植株的毒害程度。本试验表明,添加生物炭后,在分蘖期明显促进了水稻根系的生长,说明在水稻生长前期,生物炭有利于促进根系生长,随着时间的推移,生物炭的促进作用不明显,过量的生物炭甚至会抑制根系的生长。这可能与生物炭的理化性质有关,生物炭具有较高的含碳量,矿物养分含量低,施入土壤后会降低土壤养分[16],如果施入量过多,会影响水稻根系对土壤养分的吸收,进而影响地上部分的生长。
叶绿素是光合作用的物质基础,其变化必然直接或间接地影响光合作用[17-18]。本研究表明,随着镉浓度的增加,叶绿素含量降低,使得光合速率也随之降低,而光合速率的降低是叶片衰老的表现,说明镉胁迫会导致水稻叶片衰老,降低光合产量。同一镉浓度条件下,随着生物炭添加量的增加,水稻叶绿素总含量、叶面积、光合参数以及产量指标均随之先增加后降低,表明适量的生物炭(C21处理)对其有良好的促进作用。另外,分蘖期、灌浆期追施了一定量的尿素,也保证了水稻生长所需的养分,两方面的共同作用使得叶面积增加,单位面积叶绿素的含量得到提升,进而为进行更高效的光合作用提供了物质基础,同时,较大的叶面积可以获取更多的光能,提高水稻的光合作用,积累更多的光合产物,增加产量。
Pn、Tr、Ci、Gs 作为重要的光合参数,是衡量植物逆境胁迫下光合强度的重要指标[19]。本试验研究表明,同一炭量添加下,随着镉浓度的增加,水稻净光合速率、气孔导度、蒸腾速率、胞间CO2浓度均随之先升高后降低,Cd1浓度达到最大值,说明低浓度镉胁迫下,叶绿素含量越高,越有利于提升水稻对镉的耐性,增强光合能力,提高产量。随着镉浓度的增加,镉耐性降低,产量随之降低,这与Daud等[20]、钱雷晓等[21]的研究结果相同。施加生物炭(C21)后,不管有无镉胁迫,均可以提高净光合速率、气孔导度、蒸腾速率与胞间CO2浓度,提高水稻光合能力。按照Farquhar等[22]的研究,Ci与Gs 协同下降时,Pn的下降是由气孔限制,而本试验表明,胞间CO2浓度随着净光合速率的降低而增大,说明是非气孔限制。
对产量指标的研究表明,随着镉浓度的增加,水稻每盆产量先增加后降低,Cd1浓度产量最高,Cd4浓度产量最低,这与镉胁迫下水稻分蘖有关,低浓度镉可以促进水稻分蘖,增加有效穗数,提高产量;随着镉浓度的增加,水稻分蘖减少,有效穗数降低,产量下降。张秀芝等的研究也表明,随着重金属Cd、As、Cr浓度的增加,籽粒产量明显下降[22]。本试验也表明,添加生物炭后,C21处理组的每盆产量大于无炭添加C0处理,也大于过量生物炭C210处理,说明生物炭能缓解镉胁迫、增加水稻产量,最佳用量为C21处理,即每盆施加21 g生物炭,折合大田用量为4 t/hm2,且在Cd2浓度下效果最好。
参考文献:
[1]雷武生,杨宝林,戴金平.硫肥对镉胁迫下不同基因型水稻抗氧化系统和光合特性的影响[J]. 河北农业大学学报,2014,37(2):12-17.
[2]郑悦,郑桂萍,赵洋,等. 生物炭对粳稻垦鉴稻5号穗部性状及产量的影响[J]. 江苏农业科学,2015,43(6):59-62.
[3]陈志良,莫大伦,仇荣亮.镉污染对生物有机体的危害及防治对策[J]. 环境保护科学,2001,27(4):37-39.
[4]黄冬芬,奚岭林,王志琴,等. 结实期灌溉方式对水稻品质和不同器官镉浓度与分配的影响[J]. 作物学报,2008,34(3):456-464.
[5]Weng B S,Xie X Y,Weiss D J,et al. Kandelia obovata (S.,L.) Yong tolerance mechanisms to cadmium:subcellular distribution,chemical forms and thiol pools[J]. Marine Pollution Bulletin,2012,64(11):2453-2460.
[6]Su,Y,Liu,et al. Effect of iron deficiency on subcellular distribution and chemical forms Cd in peanut roots in relation to its translocation[J]. Environmental and Experimental Botany,2014,97:40-48.
[7]杨晓庆,侯仔尧,常梦婷,等. 生物炭对镉污染土壤的修复研究[J]. 江苏农业科学,2015,43(6):335-337.
[8]杨放,李心清,王兵,等. 生物炭在农业增产和污染治理中的应用[J]. 地球与环境,2012,40(1):100-107.
[9]徐楠楠,林大松,徐应明,等. 生物炭在土壤改良和重金属污染治理中的应用[J]. 农业环境与发展,2013,30(4):29-34.
[10]Kleiner K. The bright prospect of biochar[J]. Nature Reports-Climate Change,2009,3(6):72-74.
[11]Woolf D,Amonette J E,Street-Perrott F,et al. Sustainable biochar to mitigate global climate change[J]. Nature Communications,2010,1(56):1-9.
[12]孟军,张伟明,王绍斌,等. 农林废弃物炭化还田技术的发展与前景[J]. 沈阳农业大学学报,2011,42(4):387-392.
[13]慈敦伟,姜东,戴廷波,等. 镉毒害对小麦幼苗光合及叶绿素荧光特性的影响[J]. 麦类作物学报,2005,25(5):88-91.
[14]Wei S,Li Y,Zhan J,et al. Tolerant mechanisms of Rorippa globose (Turcz.) Thell. hyperaccumulating Cd explored from root morphology[J]. Bioresource Technology,2012,118:455-459.
[15]张晗芝,黄云,刘钢,等. 生物炭对玉米苗期生长、养分吸收及土壤化学性状的影响[J]. 生态环境学报,2010,19(11):2713-2717.
[16]陈心想,何绪生,耿增超,等. 生物炭对不同土壤化学性质、小麦和糜子产量的影响[J]. 生态学报,2013,33(20):6534-6542.
[17]姜永雷,唐探,陈嘉裔,等. 镉胁迫对水蕨幼苗叶绿素荧光参数和生理指标的影响[J]. 江苏农业科学,2015,43(9):357-360.
[18]赵素贞,洪华龙,严重玲. 钙对镉胁迫下秋茄叶片光合作用及超微结构的影响[J]. 厦门大学学报:自然科学版,2014,53(6):875-882.
[19]魏花朵,李悦,陈忠林,等. 氮肥对镉胁迫下结缕草光合和叶绿素荧光特性的影响[J]. 中国土壤与肥料,2015(4):88-92.
[20]Daud M K,Sun Y Q,Dawood M,et al. Cadmium-induced functional and ultrastructural alterations in Roots of two transgenic cotton cultivars[J]. Journal of Hazardous Materials,2009,161(1):463-473.
[21]钱雷晓,胡承孝,赵小虎,等. 镉胁迫对不同基因型小白菜氮代谢和光合作用的影响[J]. 华中农业大学学报,2015,34(3):69-75.
[22]Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology,1982,33:317-345.
[23]张秀芝,李强,彭畅,等. 不同添加量重金属对水稻产量及籽粒重金属富集的影响[J]. 吉林农业科学,2015,40(4):13-16.闫佳,张均,马超,等. 氮素水平对冬小麦地上部分锌积累和转运的影响[J]. 江苏农业科学,2016,44(5):102-106.
猜你喜欢
矿山安全信息(2022年40期)2022-04-07
当代水产(2021年10期)2021-12-05
军事文摘(2021年22期)2021-11-26
今日农业(2020年20期)2020-11-26
聚氯乙烯(2018年9期)2018-02-18
人间(2016年27期)2016-11-11
现代园艺(2016年13期)2016-09-02
