
鄱阳湖典型洲滩植物物种多样性季节动态特征①
2016-07-19 07:33杨桂山王晓龙万荣荣
土壤 2016年2期
关键词:鄱阳湖
李 冰,杨桂山,王晓龙*,万荣荣
(1 中国科学院南京地理与湖泊研究所,流域地理学重点实验室,南京 210008;2 中国科学院大学,北京 100049)
鄱阳湖典型洲滩植物物种多样性季节动态特征①
李冰1,2,杨桂山1,王晓龙1*,万荣荣1
(1 中国科学院南京地理与湖泊研究所,流域地理学重点实验室,南京210008;2 中国科学院大学,北京100049)
摘要:洲滩植物是湿地生态系统重要组成部分,在湿地生态系统物质循环与能量转换过程中起着关键作用。洲滩植物群落物种多样性体现湿地生态系统结构的稳定性与复杂性,对湿地生态系统健康与生态功能维持具有重要意义。本研究通过样方与样线调查法对鄱阳湖典型洲滩植物群落多样性进行了实地调查与分析。研究结果表明:鄱阳湖典型洲滩植物群落优势种重要值均值在0.5以上,在群落中占有主导地位;与春季相比,阿及苔草、水蓼、菰、香蒲和南荻等群落物种多样性在秋季有不同程度的下降,而�草群落和灰化苔草群落物种多样性在秋季则有所升高;芦苇和南荻群落在互为伴生过程中可能由于生长机理不同和生态位重叠现象,二者Shannon-Wiener多样性指数呈现出此消彼长的关系,其中春季南荻群落物种多样性较高,秋季则以芦苇群落物种多样性较高。相关分析表明,鄱阳湖湿地典型洲滩植物群落物种多样性与生物量之间无明显相关性,生物量主要受到群落内优势种的类型、生长特性、繁殖方式,以及微地形的影响。
关键词:物种丰富度;群落物种多样性;洲滩植物群落;鄱阳湖
湿地生态系统是全球生态系统的重要组成部分,也是全球生物多样性最为丰富的生态系统类型之一,在维护区域生态系统稳定、物种基因保护、资源利用乃至全球气候变化影响等方面具有其他系统所不能替代的作用[1-4]。洲滩植物群落是湿地生态系统的重要组成部分,是维持湿地生态结构和服务功能的重要支撑,其群落物种多样性已成为衡量湿地生态系统功能的重要指标[5-6]。此外,生产力作为表征生态系统功能的指标,其与物种多样性的关系也是生态学研究的热点[7-8]。已有研究表明植物物种多样性的保护对于湿地生态系统服务功能维持和改善具有重要的意义,如Engelhardt和Ritchie[9]认为维管束植物生物多样性保护是湿地生态系统服务功能维持的重要基础;Carvalho等[10]模拟了扰动发生后湿地植物多样性与群落恢复弹性的关系,发现植物群落的恢复能力与其多样性呈正相关。
鄱阳湖湿地面积约2 700 km2,1992年被列入《世界重要湿地名录》,是我国湿地生态系统中生物资源最丰富的地区,也是我国公布的首批国家重点湿地保护地之一。近年来鄱阳湖湖泊水位异常变动,春秋季干旱有所加剧,旱涝急转现象日渐凸显[11],给鄱阳湖洲滩湿地植物带来了一系列影响,使局部湖区湿地植物发生正向演替,突出表现在高滩湿地植被逐步为中生性草甸替代,水陆过渡带植物生物多样性下降以及新出露区域水生植被萎缩[12]。此外,由于泥沙的淤积和人工围垦使得鄱阳湖湿地面积急剧减少,植物多样性遭到严重威胁[13],鄱阳湖湿地植物多样性特征以及湿地生态系统功能研究已经受到越来越多国内外学者的关注。一些学者基于遥感图像,从景观尺度研究了鄱阳湖典型湿地植物的面积分布和植物演替[14-15],以及三峡水库蓄水运行对二者的影响机制[16]。此外,也有学者通过野外调查和实验观测探讨了鄱阳湖湿地植物群落生长特征和演替机制与水位季节变动、土壤环境因子、土壤微生物量的响应关系[17-20]。
水文过程是鄱阳湖生态系统动态平衡的关键驱动要素,由于鄱阳湖特殊的水文节律使洲滩湿地呈现周期性的淹露变化,鄱阳湖典型洲滩湿地植物群落存在春秋两个快速生长期[21],然而目前对于鄱阳湖湿地植物群落特征季节动态变化的客观认识不足,尤其对典型洲滩植物群落物种多样性和生物量的季节变化特征关注极少,同时对鄱阳湖湿地典型植物群落物种多样性与生产力的关系认识不足。开展典型洲滩植被群落物种多样性和生物量的季节动态调查,对于探明鄱阳湖湿地演变过程以及揭示湿地生态系统对环境因子变化的响应特征具有重要意义。本文拟通过实地调查与样品采集分析,查明鄱阳湖典型洲滩植物群落物种多样性季节动态,同时探讨植被群落物种多样性与生物量的相关关系,以期为鄱阳湖生态系统的稳定与生物多样性保护提供数据支撑与科学依据。
1 研究区域与研究方法
1.1研究区概况
鄱阳湖位于我国江西省北部,长江中下游南岸(115°49′ ~ 116°44′ E,28°24′ ~ 29°46′N),南北长170 km,东西平均宽度16.9 km,最大宽度约74 km。汇集五河(赣江、抚河、信江、饶河、修水)及博阳河、漳河、潼河之来水,经过调蓄后由湖口注入长江(图1)。该区气候属典型的亚热带季风气候,气候温暖湿润,雨量充沛,年平均气温17.6℃,年降水量1 450 ~ 1 550 mm,多集中在4—6月。鄱阳湖湿地以湖泊水体为核心,向外缘逐步演变为洲滩、平原、岗地、丘陵、山地渐次上升的地貌结构形态。鄱阳湖水文情势季节性周期性变化明显,具有“高水湖相,低水河相”的独特自然地理景观,丰水期湖泊面积可达4 600 km2[22],湿地处于典型的湖相水文状态,随着鄱阳湖水位的降低,不同高程的洲滩依次出露,滩地和沼泽广布,呈现出河、湖、滩交错分布的湿地景观特征[23]。这种特殊的状态使鄱阳湖湿地出现了大面积土壤肥沃、水热条件充裕的洲滩和浅滩,湿地植物大量发育。
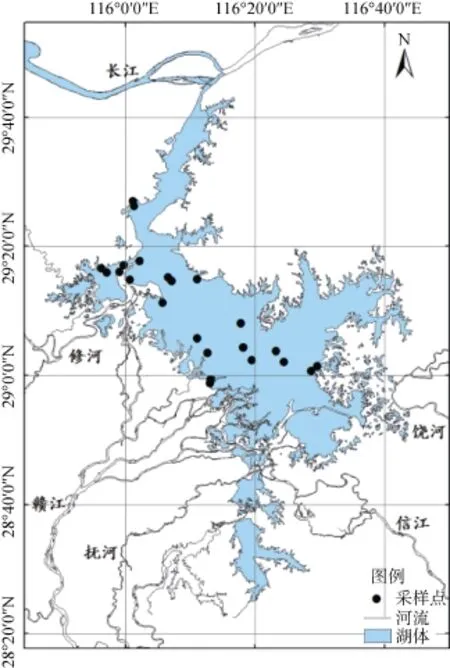
图1 鄱阳湖概况与采样点位置图Fig. 1 The Poyang Lake area and location of sampling sites
1.2样品采集与分析
由于夏季鄱阳湖水面上涨洲滩植物被淹没,短时间内沉水植物较难萌发,而在秋季洲滩再次出露后植物继续生长,为研究鄱阳湖洲滩湿地植物物种多样性季节动态,选取春季洪水期前以及秋季洪水完全消退后植被生长茂盛期作为鄱阳湖季节动态特征调查的对象。于2010年4月1—10日和10月15—25日对全湖范围内 9种典型洲滩植物群落进行样方与样线调查,包括虉草(Phalaris arundinacea)群落、灰化苔草(Carex cinerascens)群落、阿及苔草(Carex argi)群落、蓠蒿(Artemisia selengensis)群落、水蓼(Polygonum hydropiper)群落、芦苇(Phragmites communis)群落、南荻(Miscanthus sacchariflor)群落、菰(Zizania latifolia)群落和香蒲(Typha angustifolia)群落。两次调查均采用手持GPS定位,采样地点坐标尽量一致,针对每种群落选取了3条典型样地带,在各典型样地带内布置好样线后利用1 m带内布样方框随机抛取3 ~ 5个植物样方作为重复,记录每个样方优势种植物、植物物种数量、株数、盖度、平均长度、样地的海拔高度以及土壤类型等。现场利用长柄镰刀收割样方内全部植物植株,将腐黄植株去除并吸附干表面水珠后通过便携式电子天平称量地上部分的鲜重。各群落优势种与伴生种概况见表1。
1.3数据分析
由于湿地植物群落物种多样性指标无法用单一指标来全面的衡量,因此从多样性、丰富度、均匀度、优势度4个方面来比较[24-25]。
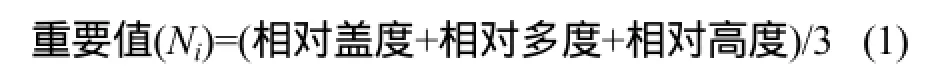
式中:相对盖度是群落中某植物盖度占总盖度的百分比,它可以反映植物种群在地面上的生存空间,在一定程度上是植物利用环境及影响环境程度的反映[26];相对多度是某植物的株数占所有种的株数的百分比;相对高度是某植物的高度占群落植物高度总和的百分比。

表1 鄱阳湖典型洲滩湿地植物群落优势种与伴生种Table 1 The dominant and companion species of typical hygrophilous vegetation in Poyang Lake
Simpson优势度指数:

式中:S表示群落中物种的数目,Ni表示第i种植物的重要值,N表示群落内总的重要值。Simpson指数是反映群落优势度的较好指标,能够表示群落内优势种的集中程度,也可以表示对环境的适应性,生态适应范围。
Shannon-Wiener多样性指数:
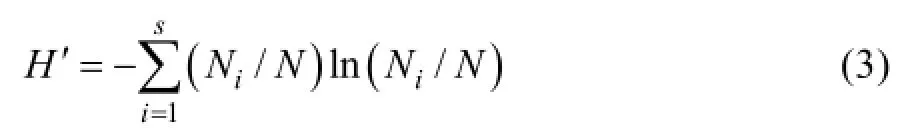
式中:各参数意义同式(2)。Shannon-Wiener指数被认为是一种较好地反映个体密度、生境差异、群落类型、演替阶段的指数[27]。也有学者指出该指数可用于表征植物的生态位宽度,生态位宽度越大,植物可利用资源越丰富且利用资源的能力越强,越倾向于广布物种[28]。
Margalef丰富度指数:

式中:P代表样方内植物的总株数,其余各参数意义同式(2)。
Alatalo均匀度指数:

式中:λ表示Simpson优势度指数,H′代表Shannon-Wiener多样性指数。均匀度指数是反映群落内个体数量分布均匀程度的良好指标,若种间的个体差异程度越小,群落内的均匀度就越高[29]。
用上述各式来计算各个样点的物种多样性,然后取各样点平均值作为植物群落的物种多样性指标[30]。数据处理和统计分析采用 Excel和 SPSS17.0软件,在P<0.05水平下进行方差分析显著性和相关性检验。
2 结果与分析
2.1鄱阳湖典型植物群落优势种重要值季节动态
从图2中可以看出,春季,9种典型群落优势种重要值以灰化苔草群落最高(0.64),其次为菰群落(0.62)和芦苇群落(0.61),三者无显著差异;南荻、水蓼和蓠蒿群落优势种重要值最低,分别为0.40、0.36、0.31;其中灰化苔草群落重要值显著高于虉草和蓠蒿群落。秋季,水蓼群落(0.72)、阿及苔草群落(0.69)、南荻群落(0.67)、菰群落(0.67)、香蒲群落(0.64)优势种重要值较高;蓠蒿(0.48)群落与灰化苔草群落(0.51)重要值最低;其中蓠蒿群落重要值显著低于菰和香蒲群落,芦苇群落重要值则显著低于南荻和菰群落。方差分析比较鄱阳湖两个季节各群落优势种重要值,发现各群落优势种重要值季节变化均未达到显著水平。
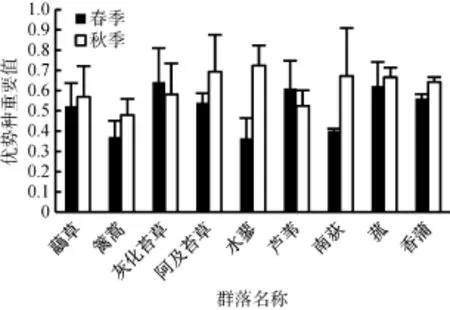
图2 鄱阳湖典型植物群落优势种重要值季节变化Fig. 2 Seasonal changes of important values of the dominant species of typical hygrophilous vegetation in Poyang Lake
2.2典型湿地植物群落物种丰富度与群落均匀度指数季节动态
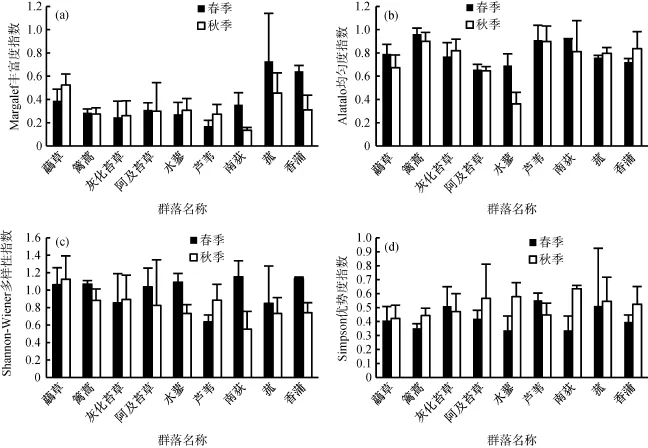
图3 鄱阳湖典型植被群落物种丰富度与群落多样性季节动态Fig. 3 Seasonal changes of biodiversity indices of typical hygrophilous vegetation in Poyang Lake
春季,蓠蒿、南荻和芦苇群落均匀度指数最高(图2b),分别为0.96,0.92和0.91,其余群落均匀度指数在0.66 ~ 0.79;其中灰化苔草和阿及苔草群落均匀度指数最低,草群落均匀度指数显著低于蓠蒿和南荻群落。秋季与春季相似,以蓠蒿群落(0.90)和芦苇群落(0.90)均匀度指数最高,其余群落均匀度指数在0.36 ~ 0.84,其中蓠蒿群落均匀度指数显著高于草、阿及苔草群落。与秋季相比,灰化苔草群落的均匀度指数存在极显著的升高(P<0.001),其他群落均匀度季节变化不显著。
2.3典型湿地植物群落生物群落多样性季节动态
春季,鄱阳湖典型植物群落 Shannon-Wiener多样性指数在0.69 ~ 0.96 (图2c),依次为南荻群落>香蒲群落>水蓼群落>蓠蒿群落>草群落>阿及苔草群落>灰化苔草群落>菰群落>芦苇群落,其中芦苇群落多样性指数最低。秋季,群落多样性指数以草群落最高,达到 1.12,其次为灰化苔草群落(0.89)、芦苇群落(0.88)、蓠蒿群落(0.88)、阿及苔草群落(0.83)、香蒲群落(0.74)、水蓼群落(0.73)和菰群落(0.73),其中南荻群落的多样性指数最低(0.55)。蓠蒿群落与芦苇群落的多样性指数季节变化显著,其中蓠蒿群落多样性指数秋季显著下降(P=0.03),芦苇群落多样性指数秋季显著上升(P=0.04),南荻和香蒲群落多样性指数也有较大的季节差异,但是差异不显著(P=0.06);其余群落多样性指数秋季均有所降低,但也未达到显著水平。
与 Shannon-Wiener多样性指数相反,春季鄱阳湖典型植物群落Simpson优势度指数以芦苇群落、灰化苔草群落和菰群落最高(图2d),分别为0.55、0.51、0.51;其次为阿及苔草群落(0.42)虉、草群落(0.41)、香蒲群落(0.40);蓠蒿群落(0.35)、水蓼群落(0.34)和南荻群落(0.34)的优势度指数则相对较低。秋季优势度指数在0.42 ~ 0.64,以南荻群落最高,草群落最低,其中蓠蒿群落与南荻、香蒲群落差异性显著,芦苇群落与香蒲群落差异性显著。与春季相比,蓠蒿群落的优势度指数在秋季有显著的升高(P=0.03),南荻群落与香蒲群落优势度指数也有季节变化,但是差异不显著(P=0.06),其余群落的优势度指数差异不显著。
2.4典型植物群落生物量与多样性指数的相关性分析
鄱阳湖湿地典型植物群落全年生物量以菰群落最高,春季达到(6 179.64 ± 593.18)g/m2,秋季(5 878.58 ± 463.18)g/m2,其余群落间无显著差异。蓠蒿、香蒲和芦苇群落生物量在秋季升高,其中蓠蒿和香蒲群落达到显著水平,其余植物群落生物量在秋季均有不同程度的降低。图4为不同样方植物生物量与各多样性指数间的回归分析,可以看出鄱阳湖典型湿地植物群落各多样性指数与生物量无明显关系。
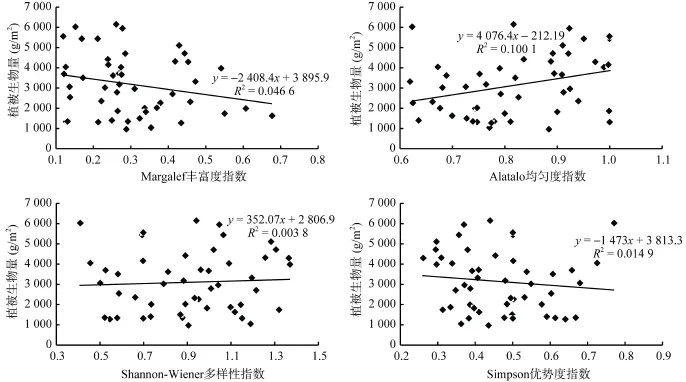
图4 鄱阳湖典型植物群落生物量与各多样性指数的关系Fig. 4 Relationship between biodiversity indices and vegetation biomass in Poyang Lake
3 讨论
优势种在群落中的功能状态对群落的性质、功能、动态和生境具有重要影响,优势种重要值可以表征群落内部优势种对生态环境资源的利用效率,以及对群落影响的重要程度[27]。灰化苔草适应的生境广,分布高程适中,营养物质丰富,为密集型克隆植物,因此灰化苔草在群落内重要值较高,但由于2010年秋季退水时间由9月中下旬延迟到10月中旬,灰化苔草休眠期比历史同期相应延长[31],抑制了其生长,进而削弱了秋季其在群落内的重要性。秋季水蓼的重要值最高,这是由于水蓼的花期一般在9—10月,春季采样时水蓼还处于发育阶段,秋季水蓼一旦长成,很快形成单一优势种群落。此外,各群落优势种重要值均值在0.5以上,在群落中占据主导地位,对维持群落结构的功能和稳定具有重要作用,不同植物群落优势种重要值存在差异,主要受到植物本身特性、土壤含水量、微地形和人为干扰的等因素的影响。
丰富度指数在单独表示多样性时有一定缺陷,在应用时必须与均匀度指数等结合,以准确反映物种多样性水平[31]。就丰富度和均匀度来说,全年来看虉草群落丰富度较高,这是由于草分布于12 ~ 15 m的高程,该区域主要为涨水前和退水后的泥滩,频繁的淹没与交替出露带来了大量的营养物质和充裕的光热条件,湿生植物繁茂,伴生种种类丰富[34]。相比其他群落,春季,芦苇群落内物种丰富度指数最小,均匀度指数较高,这是因为芦苇群落多生长于高程相对较高的台状突起地或圩堤,土壤砂粒含量高,利用群落内生态资源的能力强;此外,由于芦苇个体较大,群落内存在明显的分层现象,种间个体相对均一;灰化苔草、阿及苔草群落的丰富度指数较低,均匀度指数最低,已有研究结果也证实苔草群落结构一般较单一,伴生种较少,群落内部分布极不均匀[12,31]。秋季,香蒲群落内伴生种大多枯萎,同时由于香蒲地下茎繁殖能力强,种群扩展迅速,抑制了伴生种的生长,物种丰富度降低,随之群落均匀度升高;芦苇群落、香蒲和南荻群落丰富度指数存在显著的季节变化,这可以充分体现在伴生种的数量上(表1)。灰化苔草群落的均匀度指数存在极显著季节变化,这可能是由于秋季灰化苔草在退水延迟的影响下休眠期延长而引起的优势度降低。
Shannon-Wiener指数是能较为全面地反映植物群落多样性的指数,它的理想状态是群落内物种丰富而又分布均匀,受物种丰富度和均匀度的共同影响[30],在生态位研究中,该指数还用于揭示植物种群在群落中的功能地位、生态适应性[28,33]。而Simpson优势度指数则相反,是表征群落优势种集中程度的指标。就Shannon-Wiener指数和Simpson优势度指数而言,蓠蒿、水蓼和香蒲等大部分群落由于秋季群落内伴生种逐渐枯萎,多样性指数均有所下降。而秋季虉草群落、灰化苔草群落物种多样性较高,相关研究证实了植物对不利环境的耐受能力是以其他方面的竞争力为代价,淹水胁迫会削弱群落内竞争的影响[35],虉草和灰化苔草群落受到丰水期长期淹水胁迫,群落优势种的竞争力被削弱,为伴生种的扩展创造了机会;另一方面,已有研究表明生境条件的时间异质性较高时有利于植物的生长[36],由于二者所在的高程涨落水交替频繁具有显著的时间异质性,季节性淹水后带来的沉积物养分丰富,出露后光热资源丰富,土壤肥沃,适合多种植物生长。芦苇群落和南荻群落物种多样性季节变化显著,一方面是由于芦苇在春季长势较快[37],而南荻群落在秋季达到高峰,这使得群落内伴生种的生长空间相应变化;另一方面是因为两个群落的分布高程较为一致,常见共建群落,群落间生境竞争激烈,因此春季芦苇群落优势度较高,可能会挤压南荻群落的生境,南荻群落优势度降低,多样性相对较高;秋季则恰好相反。
总体而言,菰群落的全年生物量最高,采样点位于赣江中支口河道的两侧,植株高度在150 ~ 200 cm,群落盖度75% ~ 85%。鄱阳湖湿地大部分植物群落在秋季退水过后都过了花期和结果期,开始逐渐枯萎,生物量有所降低,而香蒲、芦苇群落的生物量在秋季有不同程度的升高,其中香蒲一般于 4月中旬至 5月上旬开始萌发,10月下旬至11月初枯黄,由于采样时间分别为4月初和10月中旬,春季采样时还未完全萌发,而秋季芦苇群落物种多样性增大,群落内伴生种数目和种类增多,如狗牙根、篱蒿、灰化苔草等,相应的生物量增加。
已有理论研究表明湿地生态系统的生产力与植物物种多样性间存在多种可能关系[38-40]。而鄱阳湖典型植物群落的生物量与物种多样性指数无明显相关,这与郑晓等[41]对呼伦贝尔草原放牧、割草两种利用方式下生物量与多样性的研究结论一致。Ma等[42]的研究也表明草地生物量与多样性之间无直接相关关系,而主要与气候因子有关;与Tilman等[43]在草地生态系统控制实验中生产力与物种数量关系的结论有所不同。这一方面可能由于不同植物不同生长阶段的含水量不同,鲜重的可对比性较差[26];另一方面,研究表明湿地植物生产力主要与洪水频率和洪水频率空间差异关系显著[38],鄱阳湖典型植物群落的生物量还与优势种的类型、生长特性、繁殖方式有关。如菰的植株相对高度高,具匍匐根状茎,生物量最大,但群落物种多样性不高。此外,Irena等[44]也证实了植物丰富度与群落生产力的关系是受尺度影响的,在较大尺度范围内,二者无显著关系。后续的研究将在固定样线的基础上加密随机采样点,消除随机性的影响,同时考虑针对同种植物同一时期内探讨群落生物量与多样性指数的响应关系。
4 结论
鄱阳湖湿地典型植物群落的优势种重要值均值在0.5以上,对群落结构稳定与生态功能维持具有重要作用。大部分植物群落如篱蒿、水蓼和香蒲等由于秋季伴生种大多数枯萎死亡,群落物种多样性均有不同程度的降低;而灰化苔草、草群落多样性在秋季则有所升高。芦苇和南荻群落由于生长过程的差异,秋季芦苇群落的多样性显著升高,而南荻群落则大幅下降。鄱阳湖典型植物群落的生物量与多样性指数间无明显的相关关系。
参考文献:
[1] Liao J J, Shen G Z, Dong L. Biomass estimation of wetland vegetation in Poyang Lake area using ENVISAT advanced synthetic aperture radar data[J]. Journal of Applied Remote Sensing, 2013, 7(1): 073579-073579
[2]Fang J Y, Wang Z H, Zhao S Q, et al. Biodiversity changes in the lakes of the Central Yangtze[J]. Frontiers in Ecology and the Environment, 2006, 4(7): 369-377
[3] Costanza R, d'Arge R, De Groot R, et al. The value of the world's ecosystem services and natural capital[J]. Nature,1997, 387(6630): 253-260
[4] Assessment M E. Ecosystems and human well-being: A framework for assessment[M]. Washington DC: Island Press, 2003
[5] Pan Y Z, Shi P J, Zhu W Q, et al. Measurement of ecological capital of Chinese terrestrial ecosystem based on remote sensing[J]. Science in China Series (Earth Sciences),2005, 48(6): 786-796
[6]Syrbe R U, Michel E,Walz U. Structural indicators for the assessment of biodiversity and their connection to the richness of avifauna[J]. Ecological Indicators, 2013, 31:89-98
[7] Guo Q, Berry W L. Species richness and biomass:dissection of the hump-shaped relationships[J]. Ecology,1998, 79(7): 2 555-2 559
[8]Tilman D, Wedin D, Knops J. Productivity and sustainability influenced by biodiversity in grassland ecosystems[J]. Nature, 1996, 379(6567): 718-720
[9] Engelhardt K A, Ritchie M E. Effects of macrophyte species richness on wetland ecosystem functioning and services[J]. Nature, 2001, 411(6838): 687-689
[10] Carvalho P, Thomaz S M, Kobayashi J T, et al. Species richness increases the resilience of wetland plant communities in a tropical floodplain[J]. Austral Ecology,2013, 38(5): 592-598
[11] 郭华, Hu Qi, 张奇. 近50年来长江与鄱阳湖水文相互作用的变化[J]. 地理学报, 2011, 66(5): 609-618
[12] 胡振鹏, 葛刚, 刘成林, 等. 鄱阳湖湿地植物生态系统结构及湖水位对其影响研究[J]. 长江流域资源与环境,2010, 19(6): 597-605
[13] 赵其国, 黄国勤, 钱海燕. 鄱阳湖生态环境与可持续发展[J]. 土壤学报, 2007, 44(2): 318-326
[14] 叶春, 刘元波, 赵晓松, 等. 基于 MODIS的鄱阳湖湿地植被变化及其对水位的响应研究[J]. 长江流域资源与环境, 2013, 22(6): 705-712
[15] 李仁东, 刘纪远. 鄱阳湖湿生植被生物量[J]. 地理学报,2001, 56(5): 532-540
[16] 余莉, 何隆华, 张奇, 等. 三峡工程蓄水运行对鄱阳湖典型湿地植被的影响[J]. 地理研究, 2011, 30(1):134-144
[17] 王晓龙, 徐立刚, 姚鑫, 等. 鄱阳湖典型湿地植物群落土壤微生物量特征[J]. 生态学报, 2010, 30(18): 5 033-5 042
[18] 董磊, 徐力刚, 许加星, 等. 鄱阳湖典型洲滩湿地土壤环境因子对植被分布影响研究[J]. 土壤学报, 2014, 51(3):618-626
[19] 张全军, 于秀波, 钱建鑫, 等. 鄱阳湖南矶湿地优势植物群落及土壤有机质和营养元素分布特征[J]. 生态学报,2012, 32(12): 3656-3669
[20] 许加星, 徐力刚, 姜加虎, 等. 鄱阳湖典型洲滩植物群落结构变化及其与土壤养分的关系[J]. 湿地科学, 2013,11(2): 186-191
[21] 张萌, 倪乐意, 徐军, 等. 鄱阳湖草滩湿地植物群落响应水位变化的周年动态特征分析[J]. 环境科学研究,2013(10): 1 057-1 063
[22] 周霞, 赵英时, 梁文广. 鄱阳湖湿地水位与洲滩淹露模型构建[J]. 地理研究, 2009, 28(6): 1 722-1 730
[23] 王晓鸿. 鄱阳湖湿地生态系统评估[M]. 北京: 科学出版社, 2004
[24] Magurran A E. Ecological diversity and its measurement[M]. Princeton: Princeton University Press, 1988
[25] 马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法. Ⅱ. β多样性的测度方法[J]. 生态学报, 1995, 3(1): 38-43
[26] 蒋政权, 洪剑明, 胡东. 北京市杨镇湿地植物群落多样性及优势种重要值的研究[J]. 湿地科学, 2004, 2(3):213-219
[27] 郑元润. 大青沟森林植物群落物种多样性研究[J]. 生物多样性, 1998, 6(3): 191-196
[28] 吴东丽, 上官铁梁, 张金屯, 等.滹沱河流域湿地植被优势种群生态位研究[J]. 应用与环境生物学报, 2006, 12(6):772-776
[29] 张桂萍, 张峰, 茹文明. 山西绵山植被优势种群生态位研究[J]. 植物研究, 2006, 26(2): 176-181
[30] 葛宏杰, 张峰. 汾河中下游湿地植被物种多样性研究[J].西部林业科学, 2008, 37(2): 29-33
[31] 胡豆豆, 欧阳克蕙, 戴征煌, 等. 鄱阳湖湿地灰化苔草草甸群落特征及多样性[J]. 草业科学, 2013, 30(6):844-848
[32] 汪殿蓓, 暨淑仪, 陈飞鹏. 植物群落物种多样性研究综述[J]. 生态学杂志, 2001, 20(4): 55-60
[33] 李峰, 谢永宏, 陈心胜, 等. 黄河三角洲湿地水生植物组成及生态位[J]. 生态学报, 2009, 29(11): 6 257-6 265
[34] 崔保山,杨志峰. 湿地学[M]. 北京: 北京师范大学出版社, 2006
[35] 杨涛, 宫辉力, 胡金明, 等. 长期水分胁迫对典型湿地植物群落多样性特征的影响[J]. 草业学报, 2010, 19(6): 9
[36] 王丽, 宋长春, 胡金明, 等. 不同时期毛苔草对不同水文情势的生长响应研究[J]. 草业学报, 2009(01): 17-24
[37] 刘晓燕, 胡东, 陈卫. 北京白河湿地主要植物群落生态学研究[J]. 湿地科学, 2004, 2(4): 296-302
[38] Pollock M M, Naiman R J, Hanley T A. Plant species richness in riparian wetlands-a test of biodiversity theory[J]. Ecology, 1998, 79(1): 94-105
[39] Waide R B, Willig M R, Steiner C F, et al. The relationship between productivity and species richness[J]. Annual Review of Ecology and Systematics, 1999: 257-300
[40] Currie D J, Mittelbach G G, Cornell H V, et al. Predictions and tests of climate based hypotheses of broad scale variation in taxonomic richness[J]. Ecology letters, 2004,7(12): 1 121-1 134
[41]翾郑晓, 王瑞东, 靳甜甜, 等. 呼伦贝尔草原不同草地利用方式下生物多样性与生物量的关系[J]. 生态学报,2008, 28(11): 5 392-5 400
[42] Ma W H, He J S, Yang Y H, et al. Environmental factors covary with plant diversity-productivity relationships among Chinese grassland sites[J]. Global Ecology and Biogeography, 2010, 19(2): 233-243
[43] Tilman D, Reich P B, Knops J M. Biodiversity and ecosystem stability in a decade-long grassland experiment[J]. Nature, 2006, 441(7093): 629-632
[44] Šímová I, Li Y M, Storch D. Relationship between species richness and productivity in plants: the role of sampling effect, heterogeneity and species pool[J]. Journal of Ecology, 2013, 101(1): 161-170
Seasonal Dynamic Characteristics of Species Diversity of Typical Hygrophilous Vegetation in Poyang Lakeshore Wetland
LI Bing1,2, YANG Guishan1, WANG Xiaolong1*, WAN Rongrong1
(1 Key Laboratory of Watershed Geographic Sciences, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing210008, China; 2 University of Chinese Academy of Sciences, Beijing100049, China)
Abstract:Hygrophilous vegetation plays a critical role in the processes of wetland ecosystem material circulation and energy transformation. Species diversity of hygrophilous vegetation communities can reflect the stability, as well as complexity of wetland ecosystem structure. Random subplots and sampling lines were utilized to investigate the typical hygrophilous vegetation communities in Poyang Lake wetland. Results showed that the average important values of dominant species of studied typical hygrophilous vegetation communities were above 0.5, which indicated their dominant status. Compared to those in spring, the diversity of Carex argi community, Polygonum hydropiper community, Zizania latifolia community, Typha angustifolia community and Miscanthus sacchariflor community declined in autumn. However, the biodiversity of Phragmites communis and Carex cinerascens community ascended in autumn, this is because that the dominance of the dominant species was weaken by delayed recession of water level, while frequently switching between grass beach and water brought substantial nutrients in autumn. Because of different growth mechanisms and niche overlapping, the diversity of Phragmites communis community and Miscanthus sacchariflor community had a reciprocal relationship along the seasons. At last, insignificant correlation was found between plant biomass and biodiversity indices.
Key words:Species richness; Community biodiversity; Hygrophilous vegetation community; Poyang Lake
中图分类号:K903
DOI:10.13758/j.cnki.tr.2016.02.014
基金项目:①国家重点基础研究发展计划(973计划)项目(2012CB417006),国家科技基础性工作专项(2013FY111800)和国家自然科学基金项目(41171024)资助。
* 通讯作者(wangxl@niglas.ac.cn)
作者简介:李冰(1991—),男,河南许昌人,博士研究生,主要从事湖泊湿地健康评价研究。E-mail: libing9133@126.com
猜你喜欢
水利水电快报(2022年7期)2022-07-18
快乐语文(2021年8期)2021-05-06
现代出版(2019年6期)2020-01-14
老友(2017年4期)2017-02-09
军事历史(1985年4期)1985-08-20
